


Abstract
Acute treatment of rats with bacterial endotoxin or particulate irritants induces the expression of CYP4A mRNAs in rat liver and kidney. To determine whether all or part of these effects could be caused by hypophagia associated with the treatments, we pair-fed saline-injected rats to rats injected with endotoxin or the particulate irritant BaSO4. The effects of endotoxin on hepatic or renal CYP4A1, CYP4A2, or CYP4A3 expression 24 h after injection were clearly distinguishable in kinetics and magnitude from those of pair feeding, indicating that the effects of endotoxin are not caused by hypophagia. Conversely, BaSO4 treatment caused a more profound hypophagia, and pair feeding to these animals produced effects similar to those of the irritant treatment, indicating that CYP4A induction by BaSO4 is mainly caused by reduced food intake. To gain further insight into the mechanism of induction of CYP4A by these inflammatory agents, we studied the sex dependence of their effects in Fischer 344 and Sprague-Dawley rats. No significant strain differences were observed, but the induction of hepatic CYP4A mRNAs by endotoxin or BaSO4 was either absent in females or significantly lower than in males. This sex specificity of induction of hepatic CYP4As has been reported previously for peroxisome proliferators, and thus our results are consistent with a role for the peroxisome proliferator-activated receptor-α in the induction of hepatic CYP4As by inflammatory agents.
Members of the CYP4A subfamily of cytochrome P450 (P4501) enzymes catalyze the ω- and ω-1-hydroxylation of fatty acids, including arachidonic acid, as well as arachidonic acid epoxidations (Nguyen et al., 1999). Expression of CYP4As in rodent livers and kidneys is induced by peroxisome proliferators (Sharma et al., 1989) and by physiological states such as diabetes or starvation in which fatty acid levels are elevated (Imaoka et al., 1990; Shimojo et al., 1993; Kroetz et al., 1998). Fatty acids and peroxisome proliferators bind to and activate peroxisome proliferator-activated receptor (PPAR)α, a member of the steroid hormone receptor superfamily (Wahli et al., 1999). PPARα binds as a heterodimer with the retinoid X receptor to enhancer elements on responsive genes, including the CYP4As (Johnson et al., 1996), resulting in their transcriptional activation. The in vivo induction of CYP4A mRNAs by chemicals, starvation, or diabetes requires PPARα, since mice lacking this receptor fail to respond to the respective stimuli (Lee et al., 1995; Kroetz et al., 1998).
CYP4A mRNAs are induced in the livers and kidneys of rats during inflammation and infection (Sewer et al., 1997), unlike most other hepatic P450 gene products, which are down-regulated (Morgan, 1997;Sewer et al., 1997). CYP4A mRNAs are also induced in the kidneys of female mice by treatment with bacterial lipopolysaccharide (LPS) treatment, and this effect is absent in PPARα-null mice, indicating that the renal induction is PPARα-dependent (Barclay et al., 1999). However, CYP4A mRNAs are down-regulated in female mouse liver after LPS treatment (Barclay et al., 1999), and therefore the participation of PPARα in hepatic CYP4A induction by inflammation in the rat remains to be elucidated. To gain further insight into this question, in the present study we examined the sex dependence of induction of CYP4As by inflammatory stimuli in the rat. A characteristic feature of hepatic CYP4A induction by peroxisome proliferators in this species is a pronounced sex difference (females are refractory) (Sundseth and Waxman, 1992).
Our previous work showing induction of renal and hepatic CYP4A mRNAs by inflammatory stimuli was conducted in Fischer 344 (F344) rats (Sewer et al., 1997). However, we also reported that hepatic induction of CYP4A2 mRNA by LPS treatment was absent in Sprague-Dawley (S-D) rats and that the hepatic induction of CYP4A1 and CYP4A3 mRNAs in this strain was attenuated relative to the effect seen in F344 rats (Sewer et al., 1997). Because a significant strain difference could be a potential tool to address the mechanism of CYP4A induction, a second goal of this study was to solidify this preliminary observation and to determine whether such a strain difference was restricted to LPS as the inflammatory stimulus.
As noted above, CYP4As are induced by starvation in a PPARα-dependent manner. LPS causes a reduction in food intake in mice (Kozak et al., 1994), but this hypophagia is not responsible for the induction of renal CYP4A by LPS because mice pair-fed with LPS-treated animals did not demonstrate renal CYP4A induction (Barclay et al., 1999). However, this observation did not rule out the possibility that hypophagia could contribute to induction of hepatic CYP4As in the rat, and the role of hypophagia in the response to particulate irritants has not been investigated. The present study addresses these questions as well. Our results are consistent with a sex-specific, PPARα-mediated induction of CYP4A mRNAs in rat liver, but the previously reported strain difference was not detected. We found that the induction of CYP4A expression by LPS in the rat is not caused by hypophagia, whereas the effect of the particulate irritant BaSO4appears to be mainly via an effect on the animals' feeding behavior.
Materials and Methods
Animals and Treatments.
All procedures were approved by the Institutional Animal Care and Use Committee of Emory University. Male or female S-D or F344 rats (Harlan Inc., Indianapolis, IN) were allowed to acclimatize to the Animal Care Facility for at least one week after delivery, before treatment. They were 8 to 10 weeks old at the time of injection. Rats received a single i.p. injection of either 1 mg/kg (0.15 mg/ml) chromatographically pureEscherichia coli LPS, serotype 0127:B8 (Sigma, St. Louis, MO), dissolved in sterile 0.9% saline; 3 g/kg (0.45 mg/ml) BaSO4 suspended in saline; or an equivalent volume of vehicle. We have shown that this dose of LPS produces a maximal suppression of total P450 and CYP2C11 content in rat liver (Morgan 1989) and induces CYP4A expression in rat liver and kidney (Sewer et al., 1996, 1997). The dose of BaSO4used was optimized previously for maximal CYP4A induction (Sewer et al., 1997). Animals were sacrificed by CO2asphyxiation 24 h after injection. Tissues were harvested and used immediately or flash-frozen in liquid nitrogen and stored at −80°C.
In the experiment to examine the effects of LPS-induced hypophagia, male Fischer 344 rats (7–8 weeks old, Charles River Laboratories, Wilmington, MA) were divided into eight groups of five rats, each cage containing two or three rats. Rats in the first and second groups were injected i.p. with 1 ml/kg sterile saline, and rats in the third and fourth groups received an injection of 1 mg/kg LPS. The food intakes of these groups were recorded every 6 h. Rats in the fifth and sixth groups (pair-fed) were injected with saline the following day and fed every 6 h with an amount equal to the mean intake of the LPS-treated groups for the corresponding period. All rats were injected at 8:00 AM. Rats in the seventh and eighth (starved) groups were deprived of food starting at 8:00 AM. Rats were killed by CO2 asphyxiation either 12 h (groups 1, 3, 5, and 7) or 24 h (groups 2, 4, 6, and 8) after injection or food removal, before collection of organs.
The experiment to examine the effects of BaSO4-induced hypophagia was conducted in male S-D rats (9 weeks old, Charles River Laboratories). The experimental design was identical to that for LPS-induced hypophagia, except that there were six rats in each group (three rats per cage) and only one time point (24 h) was examined. Starved animals were not included in this study. The dose of BaSO4 was 3 g/kg.
RNA Preparation and Northern Blotting.
Total RNA was prepared from fresh or frozen liver or kidney by acid-phenol extraction (Chomczynski and Sacchi, 1987). Samples were stored at −80°C until analysis. RNA concentrations were determined by absorbance at 260 nm. For Northern blot assays, RNA was fractionated on a 1% agarose gel electrophoresis in the presence of 5% formaldehyde and transferred to nylon transfer membrane filters (Sambrook et al., 1989) (MagnaGraph, Micron Separations, Inc., Westboro, MA). Blots were prehybridized, hybridized, and washed as described below. The amount of probe bound to the filter was quantitated using a Molecular Dynamics (Sunnyvale, CA) 445si phosphorimager and ImageQuant software. Results were normalized to the content of glyceraldehyde-3-phosphate dehydrogenase (GAP) mRNA or rRNA using the probes described below.
Probes and Hybridization Conditions.
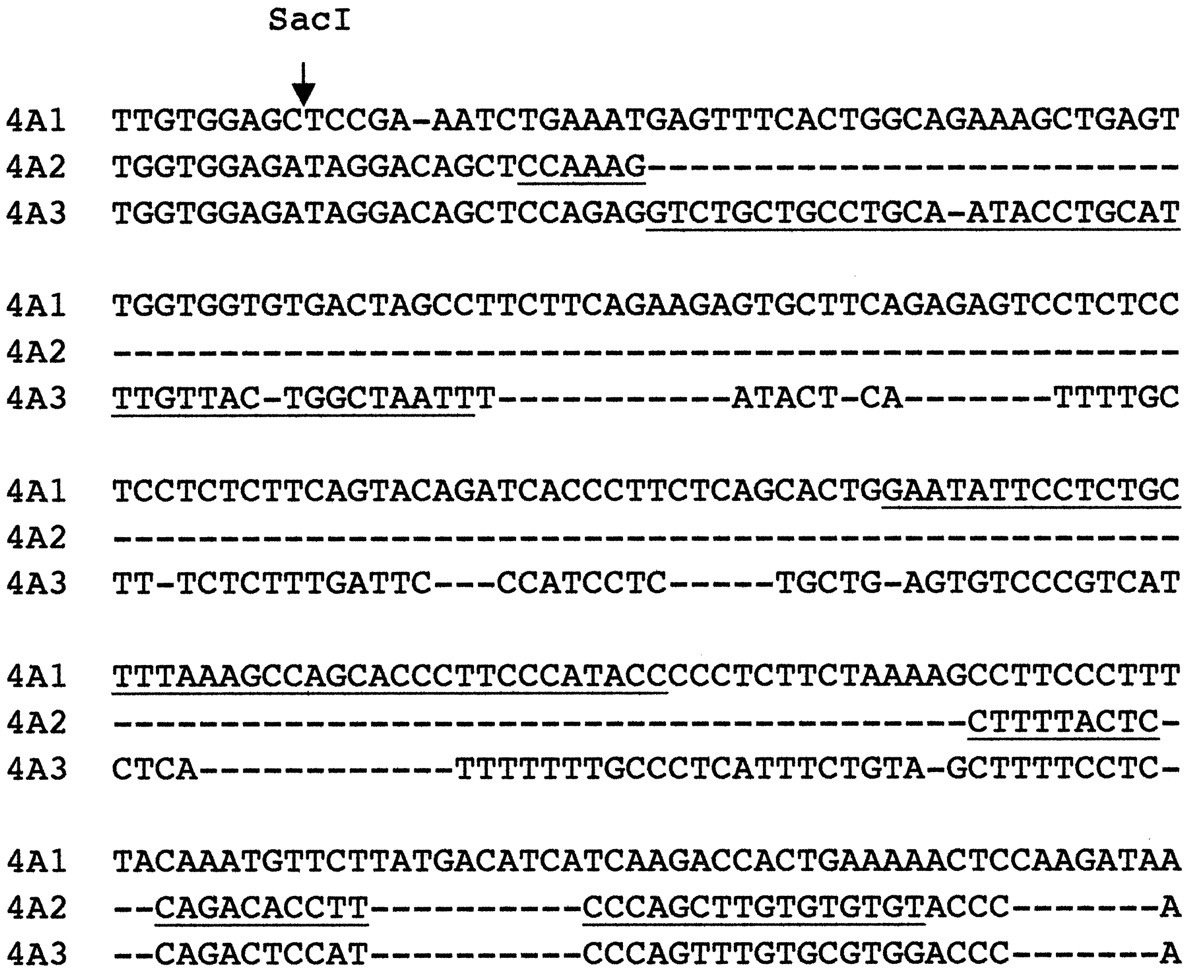
The mRNAs for CYP4A1, -4A2, and -4A3 were detected using custom synthesized oligonucleotide probes (Life Technologies Inc., Rockville, MD) designed to recognize regions of the 3′-untranslated regions of their mRNAs that have low sequence similarity. Thus, the CYP4A1 and CYP4A3 probes (Fig. 1) are complementary to a region of their mRNAs that is expressed as part of exon 12 but which is recognized as an intron, spliced, and removed in CYP4A2 (Helvig et al., 1998). The CYP4A2 probe spans this intron and therefore is specific for this mRNA (Fig. 1). The sequences of the probes were as follows: CYP4A1, 5′-GGTATGGGAAGGGTGCTGGCTTTAAAGCAGAGGAATATTC-3′; CYP4A2, 5′-ACACACACAAGCTGGGAAGGTGTCTGGAGTAAAAGCTTTGG-3′; and CYP4A3, 5′-AATTAGCCAGTAACAAATGCAGGTATTGCAGGCAGCAGAC-3′. CYP4A1 mRNA was also detected using the 3′ SacI-EcoRI fragment of its cDNA (GenBank accession no. X07259; Earnshaw et al., 1988). Fibrinogenβ-chain and GAP mRNAs were detected using their respective cDNAs. The fibrinogenβ-cDNA was kindly donated by Dr. Gerald Fuller (University of Alabama at Birmingham, Birmingham, AL); the GAP cDNA was purchased from the American Type Culture Collection (Manassas, VA). Rat 28S rRNA was detected using an oligonucleotide probe (Barbu and Dautry, 1989).
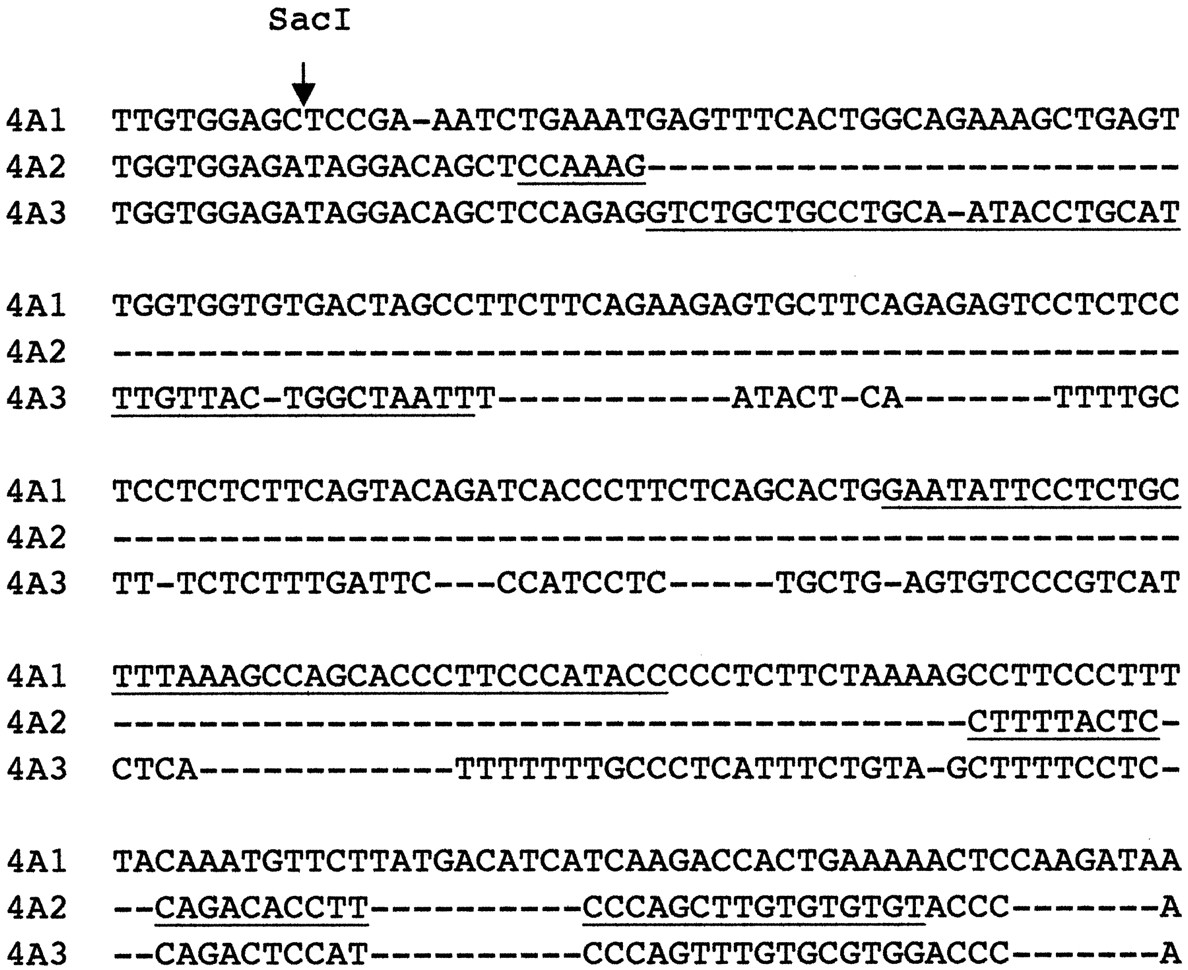
Portion of the 3′-untranslated region of rat CYP4A mRNAs.
The underlined nucleotides correspond to the portions of the sequence targeted by the oligonucleotide probes used in this study.
Oligonucleotide probes were labeled with γ-[32P]ATP using T4-polynucleotide kinase. The CYP4A2 and rRNA oligonucleotides were hybridized overnight at 50°C in 1× SSPE buffer (10 mM sodium phosphate, pH 7.7, containing 0.18 M NaCl and 1 mM EDTA); 1 mM EDTA, 0.5% SDS, 0.1 mg/ml yeast tRNA; and 5× Denhardt's solution (Sambrook et al., 1989). The CYP4A1 and CYP4A3 oligonucleotides were hybridized at 45 and 50°C, respectively, in 6× SSPE buffer containing 10% formamide, 1% SDS, 1 mM EDTA, 0.1 mg/ml yeast tRNA, and 10× Denhardt's solution. For all oligonucleotide probes, blots were washed twice for 3 min and twice for 30 min in 1× standard saline citrate (SSC) (15 mM sodium citrate pH 7.0, 0.15 M NaCl), 0.5% SDS, and twice for 30 min in 0.2× SSC, 1% SDS, all at room temperature. The final wash was for 30 min in 0.1× SSC and 0.5% SDS at 40°C. Optimal hybridization and wash conditions were determined empirically for each probe.
cDNA probes were labeled with α-[32P]dCTP by random primer labeling (Megaprime, Amersham, Arlington Heights, IL). Blots were hybridized overnight at 42°C with the CYP4A1, GAP, or fibrinogen cDNA probes in 5× SSPE buffer containing 50% formamide, 5× Denhardt's solution, 1% SDS, and 0.1 mg/ml yeast tRNA. Blots were washed twice for 30 min in 2× SSC, 0.1% SDS at room temperature, and three times for 20 min in 0.1× SSC, 0.1% SDS at 55°C (CYP4A1, GAP) or 62°C (fibrinogen).
Data Presentation and Statistical Analysis.
The Northern blot assay is semiquantitative only. Therefore, no attempt has been made to express measurements in any absolute units. Values for each experiment were calculated as a percentage of the normalized mean value (arbitrary units) for an appropriate control group. All results are expressed as the mean ± S.E.M. for each group. The numbers of animals used in each experiment are given in the figure legends. The number of observations in some groups was lower than the number of animals due to loss of samples during preparation and/or the assay procedure. Statistical analyses were performed using the Quick Statistica package (StatSoft, Inc., Tulsa, OK). One-way analysis of variance and the Student Newman-Keuls test were used to determine differences among treatment groups. In cases where the variances of the experimental groups were not equivalent, the Mann-Whitney Utest was used instead. The null hypothesis was rejected atP < 0.05.
Results
Specificity of the CYP4A Probes.
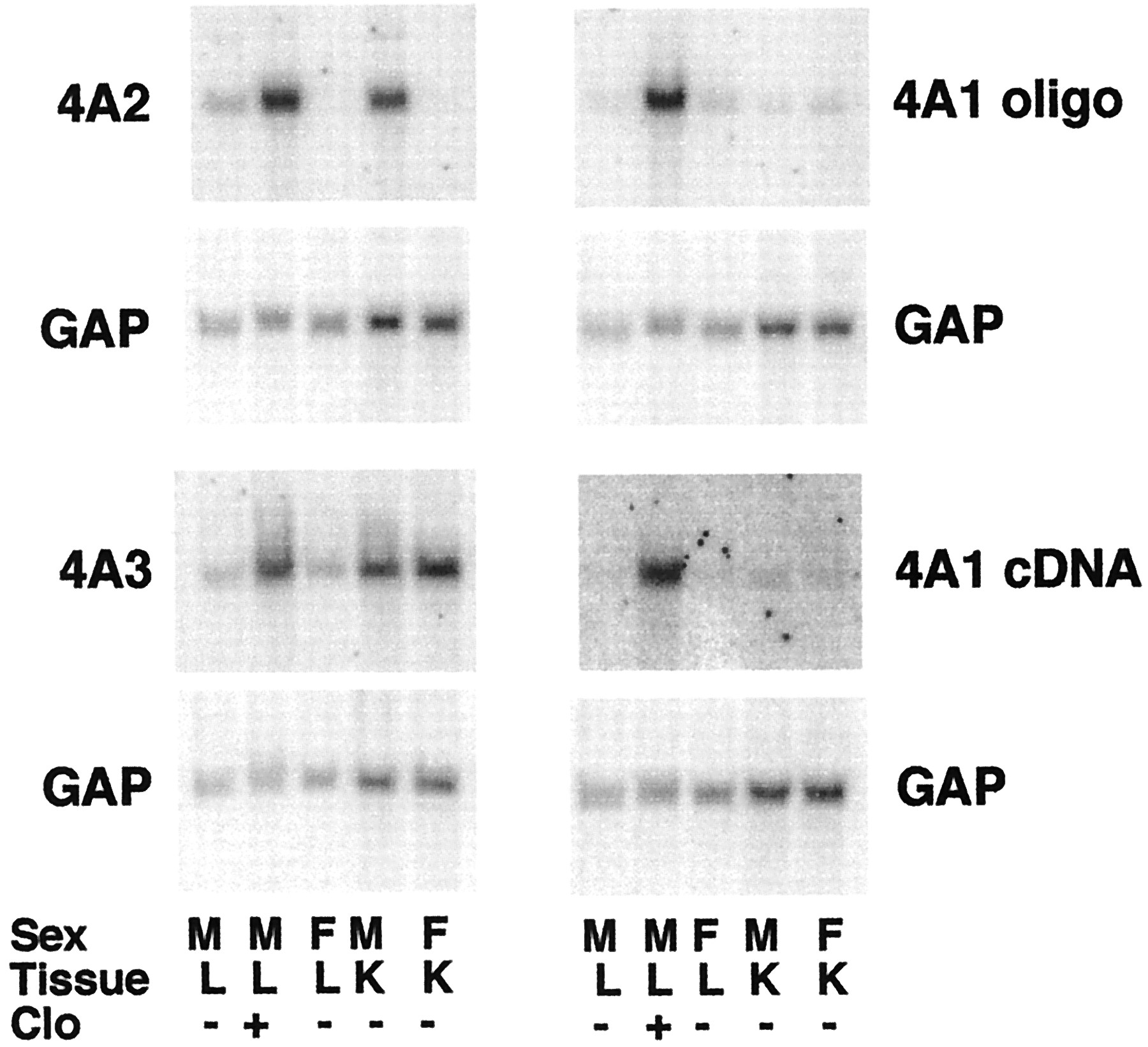
Figure 2 shows the specificities of the probes used to detect the CYP4A mRNAs, as judged by the known tissue and sex-specific expression of these genes. Thus, all three CYP4A mRNAs are induced by clofibrate. Both CYP4A2 and CYP4A3 are expressed at higher levels in kidney than in liver. CYP4A3 shows no sex specificity, whereas CYP4A2 is male-specific. The CYP4A1 oligonucleotide probe detected only low levels of constitutive expression of this mRNA in both liver and kidney, independent of gender (Fig. 2). Identical results were obtained using the CYP4A1 cDNA probe (Fig. 2). The CYP4A1 cDNA probe was selected for use in subsequent experiments because it was found to give a higher signal to noise ratio in most blots.
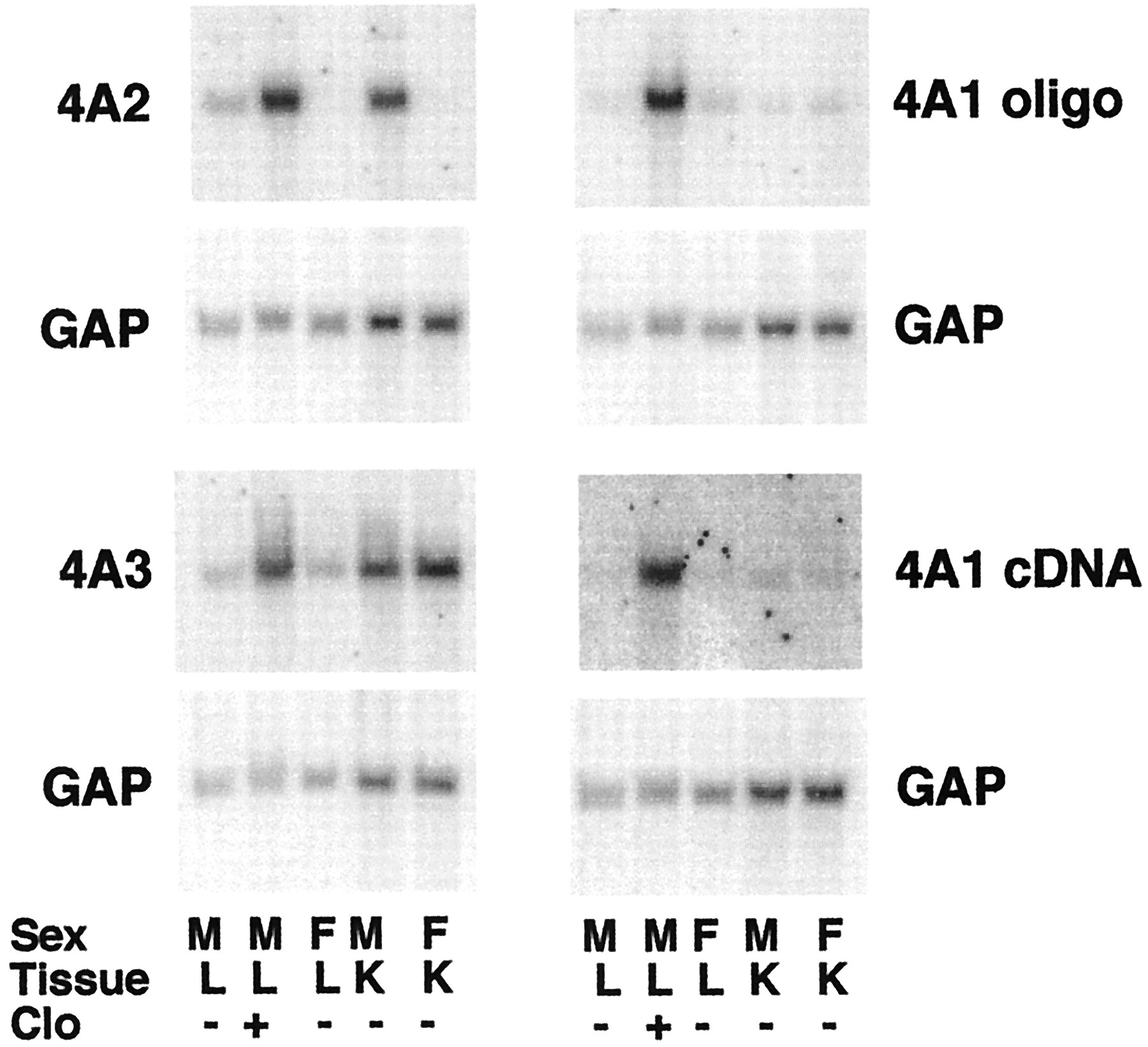
Northern blots showing specificities of the CYP4A probes.
Northern blots of sexually mature male or female F344 rat liver or kidney RNA (10 μg per sample) were hybridized with the indicated probes. Each sample was pooled from the tissues of two animals. M, male; F, female; Clo, male rats injected with a single dose of clofibrate (400 mg/kg) 24 h before sacrifice. Each blot was probed with the indicated CYP4A probe and subsequently stripped and reprobed with the GAP cDNA.
Role of Hypophagia in LPS and BaSO4 Induction of CYP4A mRNAs.
To determine the role, if any, of hypophagia in the induction of hepatic and renal CYP4A mRNAs by LPS, the food intake of saline- and LPS-treated rats was measured at 6-h intervals. The next day, animals in the pair-fed group were given food equal to the amount eaten by the LPS-treated rats during the equivalent time intervals. It was necessary to divide the treatment period into 6-h intervals to approximate the eating pattern of the LPS-treated rats. Otherwise, the pair-fed rats could eat their entire ration in a short period at the beginning of the experiment.
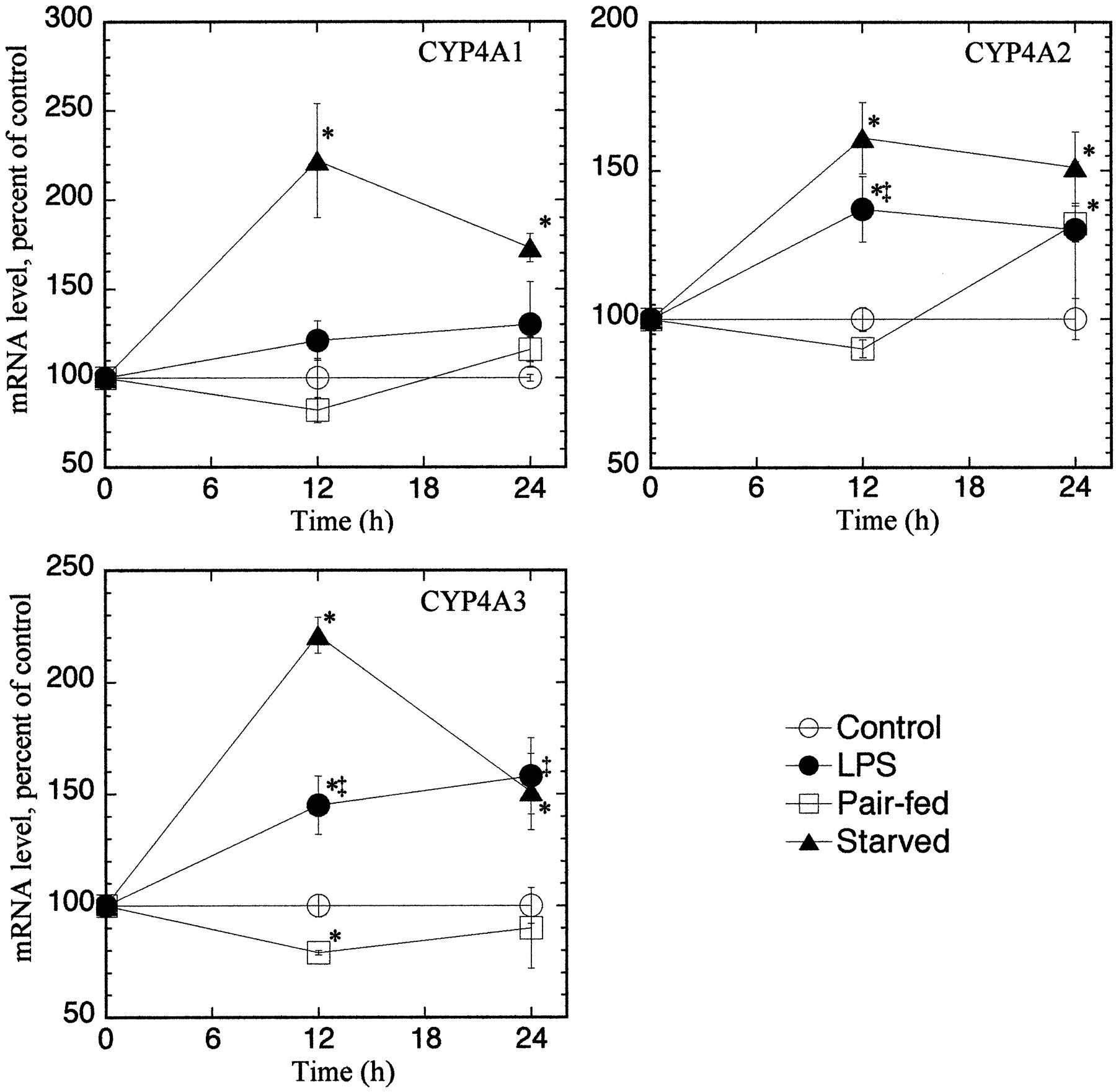
As shown in Table 1, LPS treatment caused a 54% reduction in food intake over the 24-h treatment period. Saline-treated animals ate only 29% of their daily intake during the first 12 h (light cycle) of the experiment and only 3% during the first 6 h. The effects of pair feeding on hepatic and renal CYP4A mRNA expression are compared with those of LPS treatment and starvation in Figs. 3 and4, respectively. In the liver, pair feeding had no significant effect on CYP4A expression, except for a 72% increase in CYP4A3 mRNA at the 24-h time point (Fig. 3). LPS significantly induced the mRNAs for CYP4A1 and CYP4A3, although the effect of starvation was greater than that of LPS. A significant induction of hepatic CYP4A2 mRNA by LPS was not detected in this experiment, although there was a tendency toward an increase. Similar trends were observed for renal CYP4A expression (Fig. 4). Thus, pair feeding did not induce renal CYP4A1 or CYP4A3 expression, and it only induced CYP4A2 mRNA at the 24-h time point. In contrast, LPS treatment induced CYP4A2 and CYP4A3 at the 12- and 24-h time points. CYP4A1 expression was induced by starvation but not by LPS treatment (Fig. 4).
Food intake in saline- and LPS-treated male F344 rats

Effect of LPS, pair feeding, and starvation on hepatic CYP4A mRNA expression.
Groups of four to five, 8-week-old (198 ± 8 g b.wt.), male F344 rats were treated as described under Materials and Methods and sacrificed at the indicated times. CYP4A mRNA expression was measured by Northern blotting and normalized to the rRNA contents of the samples. Values are the means of four to five observations per group. The data for one animal in the 24-h LPS group was discarded because it deviated by more than 2 standard deviations from the group mean. *Significantly different from control group mean;‡significantly different from pair-fed group mean.

Effect of LPS, pair feeding, and starvation on renal CYP4A mRNA expression.
Groups of four to five male F344 rats were treated as described underMaterials and Methods and sacrificed at the indicated times. CYP4A mRNA expression was measured by Northern blotting and normalized to the rRNA contents of the samples. Values are the means of four to five observations per group. *Significantly different from control group mean; ‡significantly different from pair-fed group mean.
A similar experiment was performed to investigate the role of hypophagia in the induction of CYP4A by the particulate irritant BaSO4. As shown in Table2, BaSO4 had a larger effect on food intake than did LPS (cf. Table 1), reducing it to 20% of the value in saline-treated animals. BaSO4 induced the expression of all three CYP4A mRNAs in the liver but only produced a small elevation of CYP4A1 expression in the kidney (Fig. 5). The induction of CYP4A1 and CYP4A3 by BaSO4 treatment was mimicked in the pair-fed animals (Fig. 5). Pair feeding also caused an elevation of CYP4A2 mRNA in the liver, although this was significantly less than the induction caused by BaSO4 (Fig. 5).
Food intake in saline- and BaSO4-treated male Sprague-Dawley rats

Effect of BaSO4 and pair feeding on hepatic and renal CYP4A mRNA expression.
Groups of six male S-D rats (8 weeks old, 284 ± 13 g b.wt.) were treated as described under Materials and Methodsand sacrificed at the indicated times. CYP4A mRNA expression was measured by Northern blotting and normalized to the GAP mRNA contents of the samples. Values are the means of four to six observations per group. Fib, fibrinogen. *Significantly different from control group mean; ‡significantly different from pair-fed group mean.
Sex- and Strain-Dependence of CYP4A mRNA Induction by LPS and BaSO4.
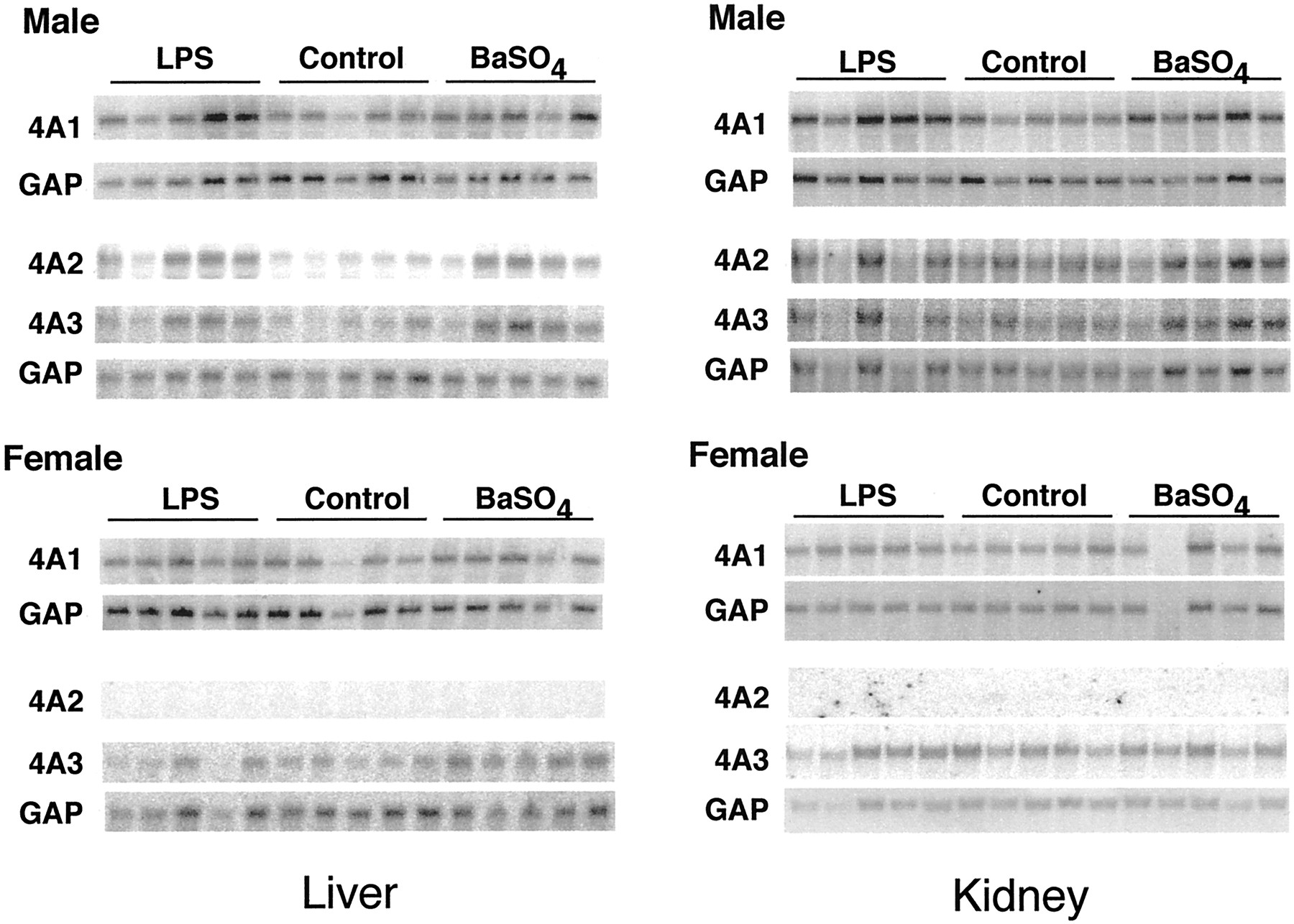
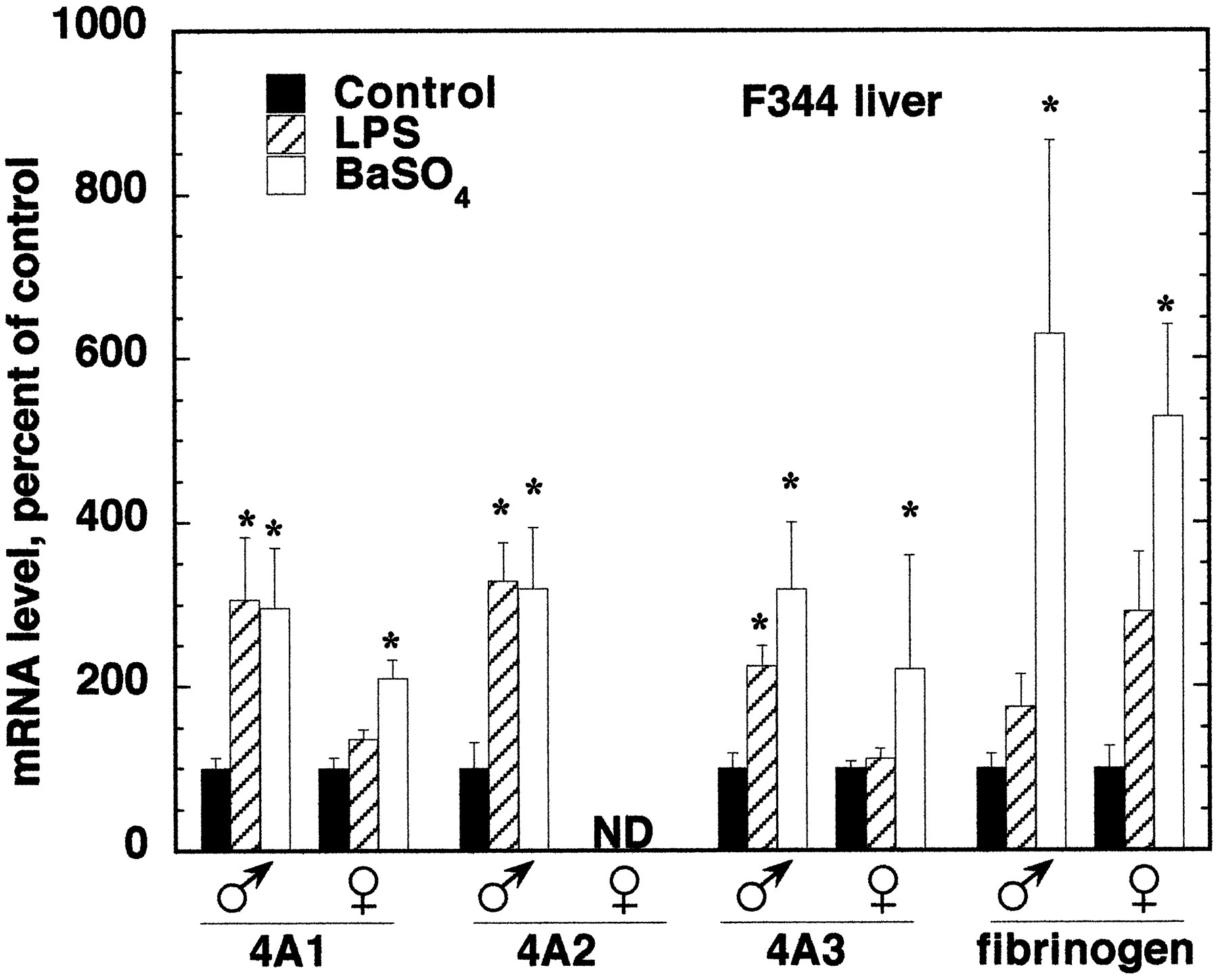
The effects of LPS or BaSO4 treatment on hepatic and renal CYP4A expression were compared in male and female rats of the F344 and S-D strains. Figure 6 shows Northern blots from the livers and kidneys of F344 rats. Figures7 and 8contain the quantitative data from this strain. Because the responses of CYP4A mRNAs in livers and kidneys of S-D rats of either sex were very similar to those of F344 rats, only the results for F344 rats are shown. LPS and BaSO4 significantly induced all three CYP4A mRNAs in the livers of F344 male rats (Fig. 7). The effects of LPS or BaSO4 treatment in male S-D rat livers were similar to those in the F344 animals, except that LPS treatment of male S-D rats produced only a trend toward an increase in CYP4A2 mRNA (LPS, 239 ± 32; control, 100 ± 74; P < 0.08) (not shown). LPS treatment failed to induce the expression of any CYP4A mRNA in the livers of female rats of either strain (F344, Fig. 7; S-D, results not shown). BaSO4 treatment produced elevations in CYP4A1 and CYP4A3 expression in female F344 and S-D rats, but these were smaller than the effects seen in the corresponding males (F344, Fig. 7; S-D, results not shown).
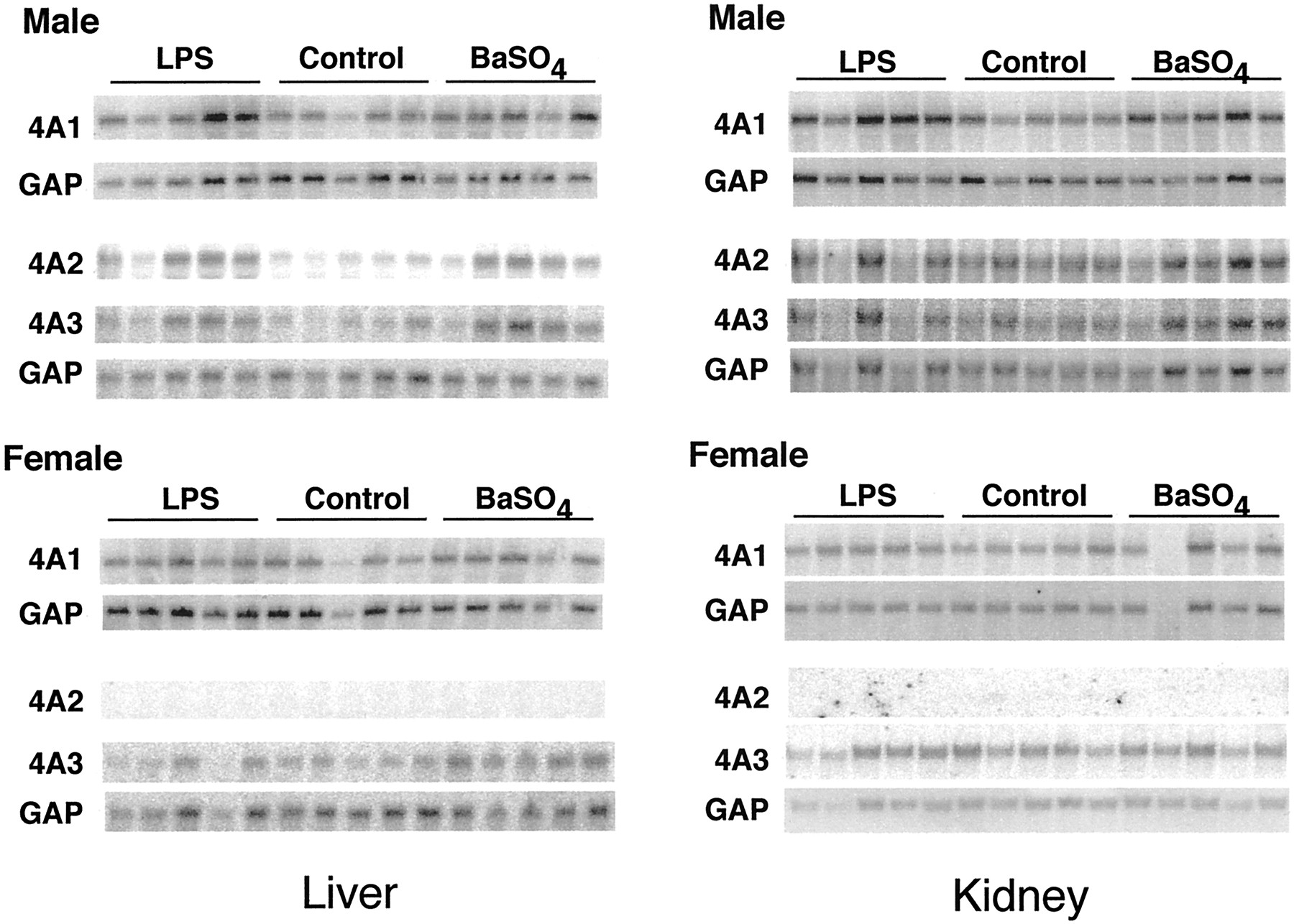
Sex dependence of CYP4A mRNA induction by LPS and BaSO4 in F344 rat liver and kidney.
Groups of five male (241 ± 10 g) and female (158 ± 11 g) F344 rats, 9 to 10 weeks old, were treated with LPS or BaSO4 as described under Materials and Methods. Twenty-four hours after treatment, total RNA was prepared and analyzed by Northern blotting. The samples shown are from individual rat livers.
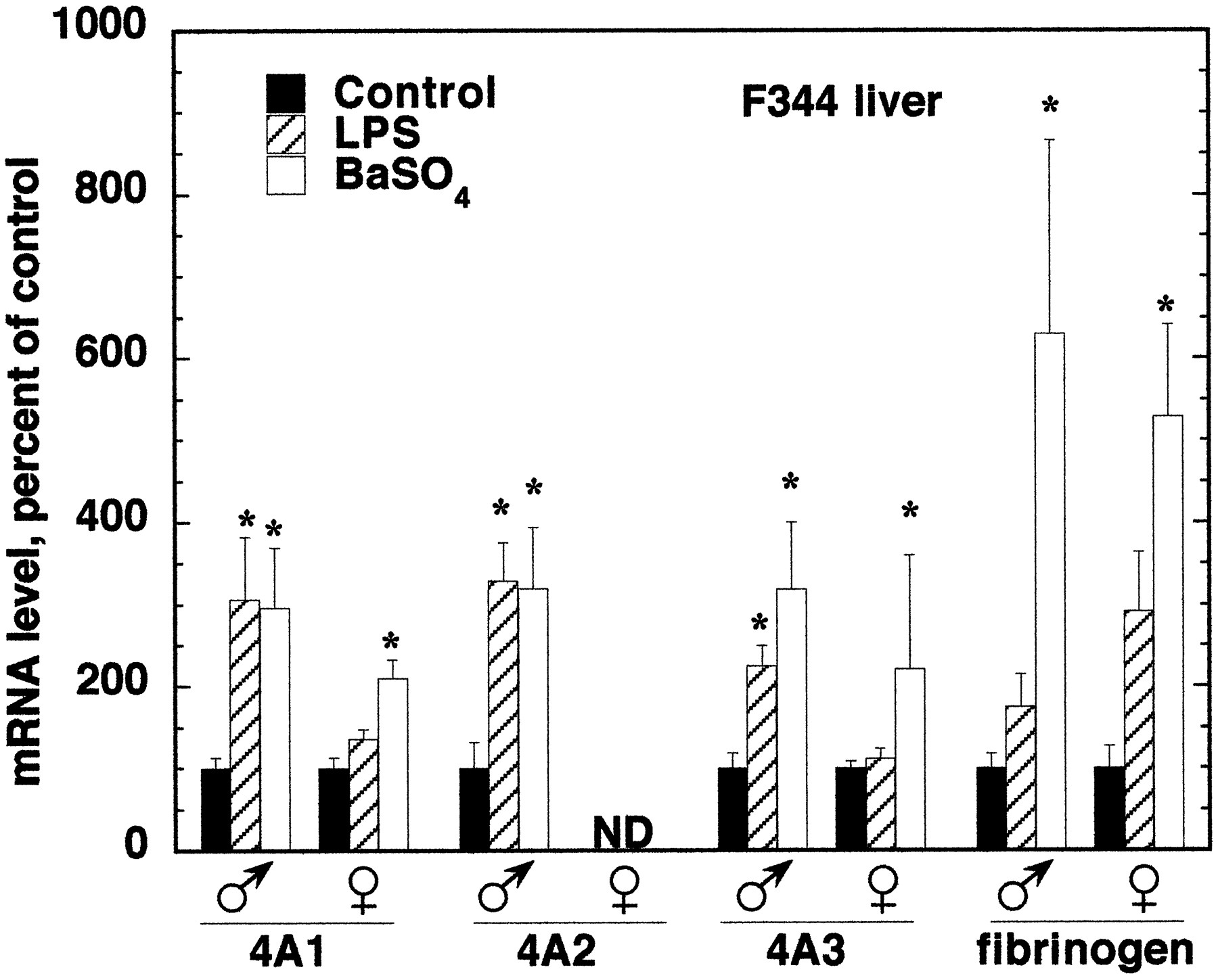
Sex dependence of CYP4A mRNA induction by LPS and BaSO4 in F344 rat liver.
Male and female F344 rats were treated with LPS or BaSO4 as described under Materials and Methods. Numbers, ages, and weights of F344 rats are given in Fig. 6. Twenty-four hours after treatment, total RNA was prepared, and the expression of CYP4A and fibrinogen mRNAs was analyzed by Northern blotting. Values are the means of four to five observations per group and are normalized to the GAP mRNA contents of the samples. *Significantly different from control group mean.

Sex dependence of CYP4A mRNA induction by LPS and BaSO4 in F344 rat kidney.
Expression of CYP4A mRNAs was measured 24 h after treatment with LPS or BaSO4. The animals and treatments were the same as described in Fig. 7. Values were normalized to the GAP mRNA contents of the samples. *Significantly different from control group mean.
The sex differences in the effects of the inflammatory stimuli were not caused by differences in the inflammatory responses of males and females, because LPS and BaSO4 produced similar effects on the expression of the hepatic acute phase protein fibrinogen in F344 rats of either sex (Fig. 7). However, the induction of fibrinogen mRNA by BaSO4 in S-D males was much greater than in females (891 ± 365% of control in males; 280 ± 63% of control in females).
In the kidney, LPS and BaSO4 treatment of F344 male rats significantly induced expression of CYP4A1 only (Fig. 8). This effect was not observed in F344 females. Conversely, LPS significantly induced renal CYP4A3 mRNA in F344 females but not in the males of this strain (Fig. 8). In S-D male rat kidneys, LPS and BaSO4 induced expression of CYP4A1, but to a lesser extent than in F344 animals (159 ± 15 and 150± 13% of control, respectively). No other effect of either stimulus on CYP4A expression was observed in S-D rat kidneys, except for a 50% increase in CYP4A1 expression in LPS-treated females (not shown).
Discussion
Our previous pair-feeding study in the mouse showed that the induction of renal CYP4A caused by LPS treatment was not caused by LPS-evoked hypophagia (Barclay et al., 1999). The results presented here indicate that this is also true of both hepatic and renal CYP4A induction in the rat. Pair feeding did not significantly elevate hepatic CYP4A1 and CYP4A2 or renal CYP4A1 and CYP4A3 expression. Although increases in hepatic CYP4A3 (Fig. 3) and renal CYP4A2 were observed with pair feeding at the 24-h time point, in both cases it can be seen that LPS treatment induced these mRNAs at the earlier (12-h) time when there was no effect of pair feeding. Thus, although LPS-induced hypophagia may produce some effect on CYP4A expression, we conclude that it is not the major mechanism. We have speculated previously that induction of CYP4As following LPS treatment is caused by generation of prostaglandins or leukotrienes during the inflammatory response, leading to activation of PPARα (Barclay et al., 1999).
Treatment of rats with the particulate irritant BaSO4 produces a more profound effect on food intake over the first 24 h than does LPS treatment (21% of control for BaSO4, 46% for LPS; see Tables 1 and2). BaSO4 treatment, or pair feeding with the BaSO4-treated animals, had similar effects on CYP4A expression in rat liver and kidney (Fig. 5), indicating that the induction of hepatic and renal CYP4As caused by BaSO4 treatment is mainly due to irritant-induced hypophagia. Thus, the threshold for reduced food intake causing a significant induction of CYP4A expression lies between 21% (Table 2, Fig. 5) and 46% (Table 1, Figs. 3 and 4).
Interestingly, although control rats ate only 25 to 29% of their daily intake in the first 12 h (and only 1–3% in the first 6 h, Tables 1 and 2), food deprivation during this short period caused significant induction of hepatic and renal CYP4As (starved animals, 12-h time point, Figs. 3 and 4). Whereas we observed no further induction over the next 12 h, Kroetz et al. (1998) reported that the induction of hepatic CYP4As at 48 h of starvation was more than double that observed at 24 h. Taken together, these observations suggest that the time course of CYP4A induction caused by starvation is biphasic, with peaks at 12 and 48 h.
Induction of hepatic CYP4A mRNAs by either LPS or BaSO4 treatment showed a clear sex dependence in rats of both strains tested (F344, Fig. 7; S-D, results not shown). Induction was either absent, or the responses were significantly smaller, in female rats. Sundseth and Waxman (1992)reported that the peroxisome proliferator clofibrate (400 mg/kg daily for 3 days) failed to induce CYP4A3 and CYP4A2 in female rats and that CYP4A1 was induced to a lesser extent in females than in males. Females were also less responsive to the induction of peroxisome marker enzymes than were males (Sundseth and Waxman, 1992). Thus, the hepatic CYP4A inducers LPS, BaSO4 (hypophagia), and peroxisome proliferators share the common characteristic of being male-specific or male-dominant, indicating that the mechanism of induction by each of these agents contains a common sex-dependent component. Since hepatic induction of CYP4As by peroxisome proliferators requires PPARα (Lee et al., 1995), our results are consistent with a role of PPARα in the hepatic induction of CYP4A by LPS in the rat, as well.
In contrast to the liver, we did not observe any clear sex dependence of renal CYP4A induction in the kidney. In S-D rats, we found that LPS or BaSO4 treatment induced only CYP4A1 mRNA, and this occurred to the same extent in either sex (not shown). In F344 rats, this effect was male-specific (Fig. 8). In contrast, CYP4A3 expression was induced by LPS treatment in F344 females only. Clearly, induction of renal CYP4A mRNAs by inflammatory stimuli is of smaller magnitude than that observed in liver (Fig. 5; compare Figs. 3 and 4, 7and 8), a characteristic that is shared with peroxisome proliferators (Sharma et al., 1989). The small magnitude of the renal effects undoubtedly contributes to the variability observed in the responses of individual CYP4As in different experiments (e.g., renal CYP4A2 and 4A3 were significantly induced by LPS in male F344 kidneys in the experiment in Fig. 4; these effects did not reach significance in the experiment in Fig. 8). More experiments with much larger numbers of animals would therefore be needed to determine whether sex and strain differences in renal CYP4A induction exist. The observed induction of CYP4A1 and CYP4A3 mRNAs in female rat kidneys (F344, Fig. 8; S-D, results not shown) is consistent with our previous report of a strong PPARα-dependent induction of renal CYP4A10 and CYP4A14 mRNAs in female mice (Barclay et al., 1999). To our knowledge, the sex dependence, if any, of renal CYP4A induction by peroxisomal proliferators in rats has not been reported, although the induction of renal peroxisomal enzymes acyl-CoA oxidase and bifunctional enzyme by dehydroepiandrosterone showed no marked sex difference (Yamada et al., 1991).
In a previous publication, we referred to unpublished data indicating that hepatic induction of CYP4A mRNAs in male S-D rats by LPS treatment was absent (CYP4A2) or reduced (CYP4A1, -4A3) compared with that seen in F344 rats (Sewer et al., 1997). The present results provide no evidence for this strain difference suggested by the previous preliminary study. The effects of LPS and BaSO4on hepatic CYP4A mRNAs were similar in both strains, except that the induction of CYP4A2 mRNA by LPS in S-D rats just failed to achieve statistical significance. This lack of a strain difference is perhaps not surprising, given that no major strain difference has been reported in the effects of peroxisome proliferators in rats and that the available evidence (present study; Barclay et al., 1999) suggests that induction of hepatic CYP4As following LPS treatment is PPARα-dependent.
In our characterization of the oligonucleotide and cDNA probes used to measure the CYP4A mRNAs (Fig. 2), the observed sex and tissue specificities of CYP4A2 and -4A3 expression detected by our probes were in general agreement with those of other groups (Sundseth and Waxman, 1992; Helvig et al., 1998). In contrast, Helvig et al. (1998) found high levels of expression of CYP4A1 in female S-D rat kidney using aSacI-EcoRI cDNA probe, whereas we observed very low basal expression of CYP4A1 mRNA in kidneys of F344 rats of either sex. The disparity was not caused by a strain or age difference: we repeated this experiment using RNA from 8- and 13-week-old S-D rats and found essentially the same results (not shown) seen in Fig. 2 using tissues from F344 rats. We obtained the same results using either an end-labeled oligonucleotide probe or a random primer-labeled cDNA fragment, both corresponding to a portion of the 3′-untranslated region of the CYP4A1 mRNA. Helvig et al. (1998) used the same cDNA fragment, labeled by nick translation. Therefore, the difference in our findings is unexplained. However, our results agree with those of Sundseth and Waxman (1992), who used an oligonucleotide probe.
In conclusion, we have shown that the induction of hepatic CYP4A after treatment of rats with LPS or the particulate irritant BaSO4 is male-specific, consistent with participation of PPARα in these responses. We have also demonstrated that whereas the induction of CYP4A expression caused by BaSO4 treatment is mainly due to hypophagia, the induction by LPS treatment is not caused by the reduction in food intake in these animals.
Footnotes
-
Send reprint requests to: Edward T. Morgan, Ph.D., Department of Pharmacology, Emory University, Atlanta, GA 30322. E-mail: etmorga{at}bimcore.emory.edu
-
This work was supported by Grant GM46897 from the National Institute of General Medical Sciences (to E.T.M.) and by a Howard Hughes Predoctoral Fellowship (to M.B.S.).
- Abbreviations used are::
- P450
- cytochrome P450
- GAP
- glyceraldehyde-3-phosphate dehydrogenase
- LPS
- bacterial lipopolysaccharide
- F344
- Fischer 344
- PPAR
- peroxisome proliferator-activated receptor
- S-D
- Sprague-Dawley
- SSC
- standard saline citrate
- Received May 10, 2000.
- Accepted August 28, 2000.
- The American Society for Pharmacology and Experimental Therapeutics
References
DMD articles become freely available 12 months after publication, and remain freely available for 5 years.Non-open access articles that fall outside this five year window are available only to institutional subscribers and current ASPET members, or through the article purchase feature at the bottom of the page.
|





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}