


Visual Overview
Abstract
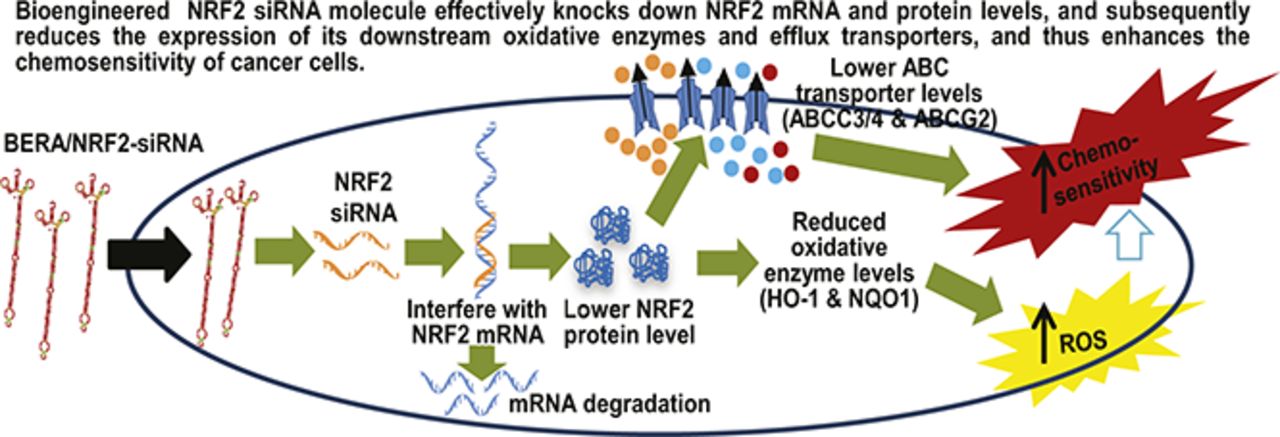
The nuclear factor (erythroid-derived 2)-like 2 (NRF2) is a transcription factor in the regulation of many oxidative enzymes and efflux transporters critical for oxidative stress and cellular defense against xenobiotics. NRF2 is dysregulated in patient osteosarcoma (OS) tissues and correlates with therapeutic outcomes. Nevertheless, research on the NRF2 regulatory pathways and its potential as a therapeutic target is limited to the use of synthetic small interfering RNA (siRNA) carrying extensive artificial modifications. Herein, we report successful high-level expression of recombinant siRNA against NRF2 in Escherichia coli using our newly established noncoding RNA bioengineering technology, which was purified to >99% homogeneity using an anion-exchange fast protein liquid chromatography method. Bioengineered NRF2-siRNA was able to significantly knock down NRF2 mRNA and protein levels in human OS 143B and MG63 cells, and subsequently suppressed the expression of NRF2-regulated oxidative enzymes [heme oxygenase-1 and NAD(P)H:quinone oxidoreductase 1] and elevated intracellular levels of reactive oxygen species. In addition, recombinant NRF2-siRNA was effective to sensitize both 143B and MG63 cells to doxorubicin, cisplatin, and sorafenib, which was associated with significant downregulation of NRF2-targeted ATP-binding cassette (ABC) efflux transporters (ABCC3, ABCC4, and ABCG2). These findings support that targeting NRF2 signaling pathways may improve the sensitivity of cancer cells to chemotherapy, and bioengineered siRNA molecules should be added to current tools for related research.
Footnotes
- Received September 22, 2017.
- Accepted October 18, 2017.
This work was supported in part by Outstanding Medical Academic leader Program of Hubei Province and Huanghe Yingcai Program of Wuhan (A.-X.Y.) and the National Institutes of Health [Grants R01GM113888 and U01CA175315 (A.-M.Y.)]. P.-C. L. was supported by a Graduate Student Fellowship from the Academic Exchange Program at Wuhan University. J.L.J. was supported by a NIGMS-funded Pharmacology Training Program Grant [T32GM099608].
- Copyright © 2017 by The American Society for Pharmacology and Experimental Therapeutics
DMD articles become freely available 12 months after publication, and remain freely available for 5 years.Non-open access articles that fall outside this five year window are available only to institutional subscribers and current ASPET members, or through the article purchase feature at the bottom of the page.
|





{kind=link}