


Abstract
Flunixin meglumine is a nonsteroidal anti-inflammatory drug (NSAID) widely used in veterinary medicine. It is indicated to treat inflammatory processes, pain, and pyrexia in farm animals. In addition, it is one of the few NSAIDs approved for use in dairy cows, and consequently gives rise to concern regarding its milk residues. The ABCG2 efflux transporter is induced during lactation in the mammary gland and plays an important role in the secretion of different compounds into milk. Previous reports have demonstrated that bovine ABCG2 Y581S polymorphism increases fluoroquinolone levels in cow milk. However, the implication of this transporter in the secretion into milk of anti-inflammatory drugs has not yet been studied. The objective of this work was to study the role of ABCG2 in the secretion into milk of flunixin and its main metabolite, 5-hydroxyflunixin, using Abcg2(−/−) mice, and to investigate the implication of the Y581S polymorphism in the secretion of these compounds into cow milk. Correlation with the in vitro situation was assessed by in vitro transport assays using Madin-Darby canine kidney II cells overexpressing murine and the two variants of the bovine transporter. Our results show that flunixin and 5-hydroxyflunixin are transported by ABCG2 and that this protein is responsible for their secretion into milk. Moreover, the Y581S polymorphism increases flunixin concentration into cow milk, but it does not affect milk secretion of 5-hydroxyflunixin. This result correlates with the differences in the in vitro transport of flunixin between the two bovine variants. These findings are relevant to the therapeutics of anti-inflammatory drugs.
Introduction
The ABCG2 membrane efflux transporter is responsible for excretion from the cells of a wide variety of endogenous compounds, xenobiotics, and drugs. It is located in the apical membrane of epithelial cells of several tissues and organs, including the intestine, liver, and mammary gland (van Herwaarden and Schinkel, 2006). Therefore, this protein transporter participates in drug-drug interactions, drug adverse effects, and even drug efficacy (Mealey, 2013; Lee et al., 2015; Robey et al., 2018).
In addition to the drug physicochemical properties related to membrane diffusion, ABCG2 is considered an important factor affecting drug transfer into milk (Ito et al., 2015). Its induced expression in mammary gland during lactation plays an important role in the active secretion into milk of different compounds (Jonker et al., 2005) such as antibiotics (Otero et al., 2013, 2016), carcinogens (van Herwaarden and Schinkel, 2006), antiparasitics (Mahnke et al., 2016), and natural compounds (Miguel et al., 2014; García-Mateos et al., 2017), among others.
Genetic variants of ABCG2 can alter the pharmacokinetics and bioavailability of its substrates (Mealey, 2013; Rocha et al., 2018). In the human species, there are some ABCG2 polymorphisms associated with a lower function of this transporter, such as Q141K (Tu et al., 2018) and V12M (Tamura et al., 2012), which are related to diseases and adverse drug effects in the population. In the veterinary field, the bovine ABCG2 Y581S single nucleotide polymorphism is widely spread in the Holstein population (Ron et al., 2006). It is a gain-of-function polymorphism (Merino et al., 2009) that also affects milk production and fat, protein, and lactose content of milk (Cohen-Zinder et al., 2005; Lopdell et al., 2017; Sanchez et al., 2017). Our previous in vitro studies using transduced cell models have revealed differential antimicrobial transport activity depending on the genetic variant (Real et al., 2011). In vivo pharmacokinetic studies with Y/Y homozygous and Y/S heterozygous cows have shown differences in secretion into milk of fluoroquinolones such as danofloxacin, enrofloxacin, and ciprofloxacin between both groups of animals (Otero et al., 2013, 2015, 2016).
Nonsteroidal anti-inflammatory drugs (NSAIDs) are widely used in human and veterinary medicine for their pharmacological effects such as analgesic, anti-inflammatory, and antipyretic actions. These drugs inhibit cyclooxygenase enzymes in the arachidonic acid cascade, which are involved in the biosynthesis of prostaglandins (Landoni et al., 1995; Cheng et al., 1998; Lees et al., 2004).
Flunixin (FLU) is the most important NSAIDs used in cattle (Fajt et al., 2011). It is the only anti-inflammatory agent authorized in the United States for use in milking cows (Food and Drug Administration, 2004) to treat pyrexia and endotoxemia associated with mastitis, and it is one of the few NSAIDs labeled for intravenous route authorized in lactating cows in the European Union (European Medicines Agency, 2000). However, NSAIDs are drugs under regulation in food animals due to the presence of residues that pose a serious risk for consumers (Smith et al., 2008). In the case of FLU therapeutics, 5-hydroxyflunixin (5OH-FLU) was identified as the main FLU metabolite and the marker residue in bovine milk (Feely et al., 2002) with a maximum residue limit of 40 µg/kg established by the European Medicines Agency (2000) and 2 parts per billion by the Food and Drug Administration (2004). In fact, 5OH-FLU levels are higher than FLU levels in cow milk (Feely et al., 2002), probably due in part to their different physicochemical properties and the extensive liver metabolism of FLU. Nevertheless, some studies have shown that alteration of the dose or route of administration, or the presence of disease processes, could affect the presence of violative residues even after withdrawal periods are finished (Kissell et al., 2015; Smith et al., 2015; Shelver et al., 2016).
Therefore, knowledge of the factors affecting the presence of residues of these compounds into milk is relevant for therapeutics and human health. The objective of our study was to investigate the role of ABCG2 in the pharmacokinetics and secretion into milk of FLU and its main metabolite, 5OH-FLU, using Abcg2(−/−) mice, and to research the effect of the bovine ABCG2 Y581S polymorphism in FLU and 5OH-FLU plasma and milk levels. Correlation of our in vivo data with in vitro cells studies involving murine Abcg2 and the two bovine ABCG2 variants (Y581 and S581) was also assessed.
Materials and Methods
Reagents and Chemicals.
FLU, 5OH-FLU, diclofenac sodium, niflumic acid, and Lucifer yellow were purchased from Sigma-Aldrich (St. Louis, MO). Ko143 was obtained from Tocris (Bristol, UK). For the pharmacokinetic studies, flunixin meglumine (Finadyne 50 mg/ml) was obtained from MSD Animal Health SL (Salamanca, Spain).
Cell Culture.
Madin-Darby canine kidney (MDCK) II (epithelial) parental cells and the MDCK-II Abcg2 murine transduced subclone were supplied by Dr. Schinkel (The Netherlands Cancer Institute, Amsterdam, The Netherlands). Generation and characterization of MDCK-II subclones transduced with the Y581 and S581 variants have been reported previously by our research group (Real et al., 2011). Cell culturing was performed as described elsewhere (González-Lobato et al., 2014).
Transport Studies.
Transepithelial transport assays were carried out as previously described with minor modifications (Pavek et al., 2005; Real et al., 2011) using parental MDCK-II, Abcg2 murine-transduced subclones, and bovine ABCG2-transduced subclones. At the end of the experiment, confluence of the monolayer was checked with Lucifer yellow permeability assays (Oltra-Noguera et al., 2015).
At the beginning of the experiment (t = 0) medium was substituted in an apical or basolateral compartment with fresh medium including 20 µM of FLU or 10 µM of 5OH-FLU, with or without Ko143. Concentrations were chosen based on sensitivity of high-performance liquid chromatography analysis. Hanks’ balanced salt solution (Sigma-Aldrich) supplemented with 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid and 25 mM HEPES (pH 7.4) was used as transport medium. Aliquots of 100 μl were sampled each hour up to 4 hours from the opposite compartment and the same volume of culture media was replaced each time. Samples were kept at −20°C until analysis.
Active transport across MDCK-II cells was reported as the relative transport ratio, determined as the concentration permeated toward the apical compartment (basolateral-to-apical transport) divided by the concentration permeated toward the basolateral compartment (apical-to-basolateral transport) at 4 hours.
Animal Experiment.
European legislation was applied to animal handling (European Community, 2010). Experimental methods were approved by the Animal Care and Use Committee of University of Leon and Junta de Castilla y Leon (ULE_011_2016 and ULE_002_2017).
Milk Secretion Studies in Mice.
Lactating female Abcg2(−/−) and wild-type mice (N = 3–7, 9–13 weeks of age) were supplied by Dr. Schinkel (The Netherlands Cancer Institute). They were maintained in an environment controlled for temperature and light/dark cycle.
Pups around 10 days old were removed from their mothers 4 hours before the experiment. For intravenous administration of FLU or 5OH-FLU (4 mg/kg), 150 µl of drug solution/30 g b.wt. was administered into the tail of mice under anesthesia with isoflurane. The drug solution consisted of 10% (v/v) Finadyne and 90% (v/v) saline solution for the FLU experiments. In the case of 5OH-FLU, the compound was dissolved in 10% (v/v) ethanol, 40% (v/v) polyethylene glycol 400, and 50% saline. Milk and plasma samples were collected using the method reported by Miguel et al. (2014).
Pharmacokinetic Studies on Dairy Cows.
Cows were kept on a private farm near Leon, Spain. Twelve lactating Holstein cows weighing ∼800 kg were used. Milk yield was 41.96 ± 6.67 kg/day. The Y581S genotypes were determined in accordance with Komisarek and Dorynek (2009). Accordingly, two groups of six Y/Y581 homozygous and six Y/S581 heterozygous cows were defined for experimental design. No differences were found in age, weight, or milk yield between the two sets of cows. Both groups were treated intramuscularly with 2.2 mg/kg of flunixin meglumine (Finadyne). Blood samples (5 ml) from the tail vein were obtained at 1, 3, 6, 14, 24, 30, and 38 hours after administration. Samples of milk were manually taken at 3 hours and using an automatic milking machine during normal milking at 6, 14, 24, 30, 38, 48, and 54 hours. No more sampling points were possible because of the private nature of the farm. Plasma was obtained by centrifugation and samples were kept at −20°C until analysis.
High-Performance Liquid Chromatography Analysis.
Determination of concentration of FLU and its main metabolite 5OH-FLU in plasma and milk samples from mouse experiments and samples of transport assays was based on previously published methods (Gallo et al., 2008) with some modifications using diclofenac as an internal standard. Mouse samples were cleaned up using the method described by Odensvik and Johansson (1995) with some modifications. Samples of culture medium were analyzed without processing. A 4 µm particle (250 × 4.6 mm) 80 Å Synergi column (Phenomenex, Torrance, CA) was used for the chromatographic separation. In transepithelial transport assays, the limit of detection (LOD) and limit of quantification (LOQ) values obtained were 0.01 and 0.03 µg/ml for FLU and 0.008 and 0.02 µg/ml for 5OH-FLU, respectively.
In the mouse experiments, the LOD and LOQ values obtained for FLU were 0.15 and 0.39 µg/ml in plasma and 0.05 and 0.11 µg/ml in milk, respectively. For 5OH-FLU, the LOD and LOQ values were 0.05 and 0.12 µg/ml in plasma and 0.03 and 0.07 µg/ml in milk, respectively.
Ultra-Performance Liquid Chromatography Quadrupole Time-of-Flight Mass Spectrometry Analysis.
Determination of concentration of FLU and its main metabolite 5OH-FLU in plasma and milk samples from pharmacokinetics studies in dairy cattle were based on previously published methods (Rúbies et al., 2016) with some modifications.
Plasma and milk samples must be processed before injection into the ultra-performance liquid chromatography system following the clean-up method previously described by Kissell et al. (2015) using niflumic acid as an internal standard. Analyses were performed using a Waters ACQuity UPLC H-Class system coupled to an ACQuity TQD tandem quadrupole mass spectrometer (Waters Corporation, Milford, MA). A Phenomenex Kinetex C18 column (2.6 µm, 100 × 3.00 mm) was used for the chromatographic separation. Experiments were performed in positive ion mode at mass-to-charge values of 313, 297, and 283 for 5OH-FLU, FLU, and niflumic acid, respectively. Masslynx software (version 4.1; Waters Corporation) was used for data acquisition and processing.
Plasma LOD was 2.84 ng/ml for FLU and 4.83 ng/ml for 5OH-FLU, whereas the LOQs were 7.67 and 6.68 ng/ml for FLU and 5OH-FLU, respectively. The milk LODs were 0.17 and 1.42 ng/ml for FLU and 5OH-FLU and the LOQs were 0.41 and 3.96 ng/ml for FLU and 5OH-FLU, respectively.
Pharmacokinetic Calculations and Statistical Analysis.
The pharmacokinetic parameters were determined as reported elsewhere (Otero et al., 2018). The results are reported as the mean ± S.D. Normal distributions of data were tested by the Shapiro-Wilk test. Data with normal distribution were analyzed by the two-tailed Student’s test. Data with non-normal distribution were tested using the Mann-Whitney U test. Differences were considered statistically significant when P < 0.05. Statistical analyses were carried out with the SPSS 24 software (IBM Corporation, Armonk, NY).
Results
In Vitro Interaction of FLU and Its Main Metabolite with the ABCG2 Transporter.
To study the in vitro interaction between ABCG2 and NSAID FLU and its main metabolite 5OH-FLU, transepithelial transport assays were carried out using parental MDCK-II and their subclones transduced with murine and two variants of bovine ABCG2 (Figs. 1 and 2).
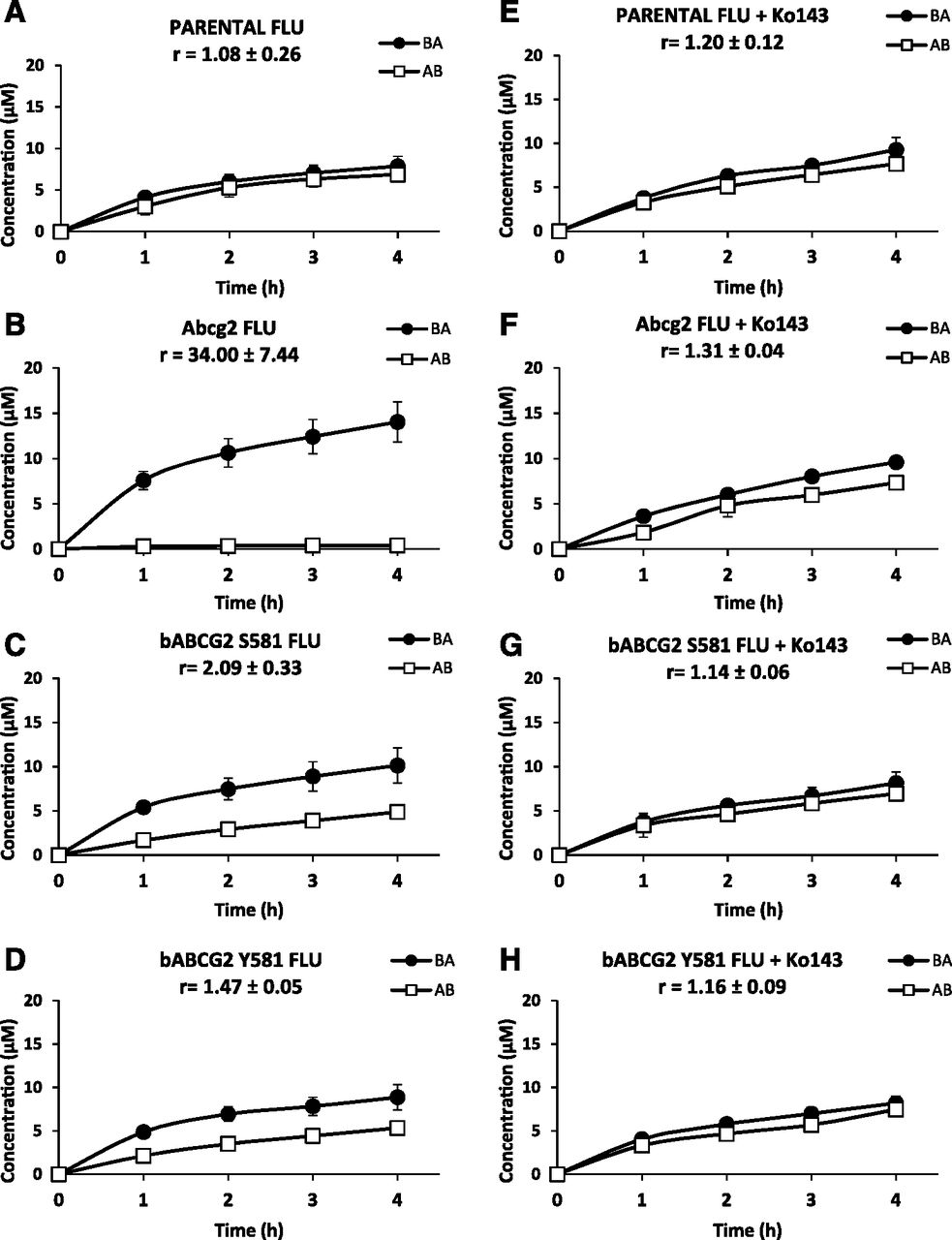
Transepithelial transport assay of FLU at 20 µM in parental MDCK-II cells and its subclones transduced with murine Abcg2 and the two bovine variants (Y581 and S581) in the absence (A–D) or presence (E–H) of the specific ABCG2 inhibitor Ko143. The experiment was started (t = 0) by replacing the medium in either the apical or basolateral compartment with fresh culture medium containing 20 µM of FLU. Aliquots of 100 µl were taken from the opposite compartment at 1, 2, 3, and 4 hours and concentrations were measured by high-performance liquid chromatography. Concentration permeated toward apical compartment represents basolateral-to-apical transport (●); concentration permeated toward basolateral compartment represents apical-to-basolateral transport (□). Results are represented as mean ± S.D.; “r” represents relative transport ratio: basolateral-to-apical transport divided by apical-to-basolateral transport at 4 hours (n = 3–6).

Transepithelial transport assay of 5OH-FLU at 10 µM in parental MDCK-II cells and its subclones transduced with murine Abcg2 and the two bovine variants (Y581 and S581) in the absence (A–D) or presence (E–H) of the specific ABCG2 inhibitor Ko143. The experiment was started (t = 0) by replacing the medium in either the apical or basolateral compartment with fresh culture medium containing 10 µM of 5OH-FLU. Aliquots of 100 µl were taken from the opposite compartment at 1, 2, 3, and 4 hours and concentrations were measured by high-performance liquid chromatography. Concentration permeated toward apical compartment represents basolateral-to-apical transport (●); concentration permeated toward basolateral compartment represents apical-to-basolateral transport (□). Results are represented as mean ± S.D.; “r” represents relative transport ratio: basolateral-to-apical transport divided by apical-to-basolateral transport at 4 hours (n = 3–6).
In the parental MDCK-II cells, apical-to-basolateral–directed translocation was equal to basolateral-to-apical translocation in FLU and 5OH-FLU assays (Figs. 1A and 2A). We observed basolateral-to-apical preferential transport of FLU and 5OH-FLU in murine Abcg2- and bovine ABCG2-transduced cells compared with the parental cells (Figs. 1, B–D, and 2, B–D). Relative efflux transport ratios at 4 hours were significantly higher for FLU (34.00 ± 7.44 in murine Abcg2, 1.47 ± 0.05 in bABCG2 Y581, and 2.09 ± 0.33 in bABCG2 S581) compared with parental cells (1.08 ± 0.26; P < 0.05). In the case of 5OH-FLU, these ratios were also significantly increased (4.30 ± 2.69 in murine Abcg2, 2.19 ± 0.46 in bABCG2 Y581, and 2.14 ± 0.65 in bABCG2 S581) compared with control cells (0.89 ± 0.14) (P < 0.05). When the selective ABCG2 inhibitor Ko143 was added the basolateral-to-apical preferential transport was completely reversed in all ABCG2-transduced subclones (Figs. 1, E–H, and 2, E–H). Furthermore, in the case of FLU the relative efflux transport ratio was significantly higher in the S581 bovine variant compared with the Y581 variant (Fig. 1, C and D) (2.09 ± 0.33 vs. 1.47 ± 0.05, respectively; P < 0.05), indicating that the S581 variant was more efficient in FLU transport. In contrast to the FLU results, no significant differences between the two bovine ABCG2 variants were found for 5OH-FLU (Fig. 2, C and D).
These results show that murine and bovine ABCG2 play an important role in the active efflux transport of the NSAID FLU and its main metabolite 5OH-FLU. Moreover, FLU is transported more efficiently by the bovine S581 variant compared with the Y581 variant. This difference was not reported for 5OH-FLU.
Plasma and Milk Levels of FLU and 5OH-FLU in Abcg2(−/−) Knockout Mice.
To study the involvement of Abcg2 in the plasma and milk levels of these compounds, we administered FLU or 5OH-FLU to wild-type and Abcg2(−/−) mice (Fig. 3). No 5OH-FLU was detected in plasma and milk after FLU administration. No significant differences were observed in the plasma concentration of FLU and 5OH-FLU after treatment in wild-type and Abcg2(−/−) mice. However, milk concentrations were more than 2.5-fold higher in wild-type mice compared with Abcg2(−/−) mice for FLU (1.85 ± 0.29 µg/ml vs. 0.72 ± 0.18 µg/ml; P < 0.01) and approximately 8-fold higher in wild-type compared with Abcg2(−/−) mice for 5OH-FLU (1.82 ± 0.90 µg/ml vs. 0.23 ± 0.17 µg/ml; P < 0.01). The milk-to-plasma ratio of FLU in wild-type mice was more than 3-fold higher than in Abcg2(−/−) mice (0.43 ± 0.17 vs. 0.14 ± 0.02; P < 0.05). In the case of 5OH-FLU, the milk-to-plasma ratio was 5-fold higher in wild-type compared with Abcg2(−/−) lactating mice (0.58 ± 0.25 vs. 0.11 ± 0.08; P < 0.01). Our results clearly show that Abcg2 plays an important role in the active secretion into milk of FLU and 5OH-FLU.

Plasma and milk concentration and milk-to-plasma ratio of FLU (A) and 5OH-FLU (B) in Abcg2(−/−) mice after intravenous administration at a dose of 4 mg/kg b.wt. Plasma and milk were collected 40 minutes after administration of FLU and 20 minutes after 5OH-FLU treatment. Milk and plasma levels were determined by high-performance liquid chromatography. Results are mean ± S.D.; *P < 0.05 and **P < 0.01 represent significant differences between both groups of mice (n = 3–6).
Plasma Pharmacokinetics and Secretion into Milk of FLU and 5OH-FLU in Dairy Cows Carrying the Y581S Polymorphism.
The effect of the Y581S polymorphism on plasma levels and secretion into milk of the NSAID FLU and its main metabolite 5OH-FLU was studied using Y/Y581 homozygous and Y/S581 heterozygous lactating cows (Figs. 4 and 5), after the administration of a single dose of Finadyne at 2.2 mg/kg b.wt. by intramuscular administration. The plasma profile was dominated by the parental drug FLU. No significant differences in the plasma levels (Fig. 4) and plasma pharmacokinetics parameters (Table 1) were observed between the two groups of animals.

Plasma concentration of FLU and 5OH-FLU after intramuscular administration of a single dose of Finadyne at 2.2 mg/kg b.wt. to Y/Y581 homozygous and Y/S581 heterozygous lactating cows. Plasma samples were collected at various times over 38 hours. Plasma levels were determined by ultra-performance liquid chromatography tandem mass spectrometry. The results are presented as mean ± S.D. (n = 6).
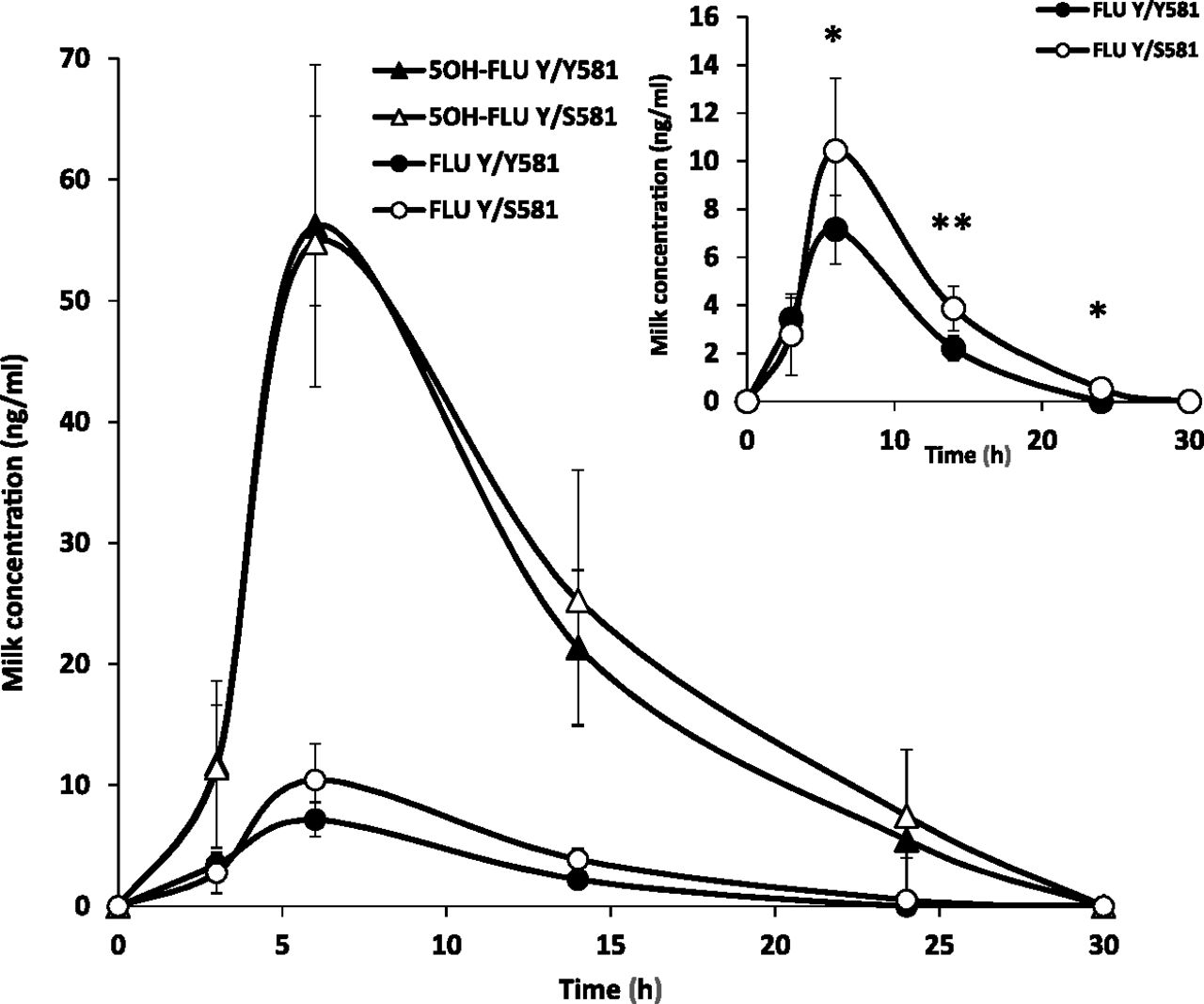
Milk concentration of FLU and 5OH-FLU after intramuscular administration of a single dose of Finadyne at 2.2 mg/kg b.wt. to Y/Y581 homozygous and Y/S581 heterozygous lactating cows. Milk samples were collected at various times over 54 hours. Milk levels were determined by ultra-performance liquid chromatography tandem mass spectrometry. Concentrations from 30 hours were undetectable. The inset shows FLU concentrations alone. The results are presented as mean ± S.D. (n = 6); *P < 0.05 and **P < 0.01 represent significant differences between both groups of cows.
Plasma pharmacokinetic parameters (mean ± S.D.) in Y/Y581 homozygous and Y/S581 heterozygous dairy cows after administration of a single dose of Finadyne at 2.2 mg/kg (n = 6)
The milk profile was dominated by 5OH-FLU (Fig. 5). No significant differences were obtained in the milk levels and milk pharmacokinetics parameters for 5OH-FLU between the two groups of cows. Only the half-life was significantly different (Table 2). However, significant differences for the secretion of FLU into milk between both groups of animals were shown (Fig. 4). The milk levels for FLU were significantly higher for Y/S581 compared with Y/Y581 animals at 6, 14, and 24 hours (P < 0.05). In addition, the area under the curve from zero to infinity (AUC0–∞) and Cmax values were approximately 1.5-fold higher in Y/S581 animals than in Y/Y581 animals (104.67 ± 25.86 vs. 69.53 ± 9.61 ng·h/ml for AUC0–∞ and 10.42 ± 2.99 vs. 7.15 ± 1.43 ng/ml for Cmax) (Table 2). The mean residence time was around 1.2-fold higher in cows carrying the Y581S polymorphism (P < 0.05; 9.15 ± 0.88 vs. 7.82 ± 0.52 hours) (Table 2). In addition, significant differences were observed between the two groups in AUC0–∞ milk-to-plasma ratios for FLU.
Milk pharmacokinetic parameters (mean ± S.D.) in Y/Y581 homozygous and Y/S581 heterozygous dairy cows after administration of a single dose of Finadyne at 2.2 mg/kg (n = 6)
Our results show that the bovine Y581S ABCG2 polymorphism increases the milk secretion of the parental compound FLU in dairy cattle; however, it does not affect secretion of 5OH-FLU into milk.
Discussion
Several pharmacokinetics and residue research studies and surveys have been previously undertaken with FLU and 5OH-FLU on different farm species such as dairy cows (Jedziniak et al., 2009, 2013; Deyrup et al., 2012; Kissell et al., 2012, 2013, 2015), sheep (Marini et al., 2016), and goats (Königsson et al., 2003). However, this is the first study to explore the in vivo role of the ABCG2 transporter in these processes for an anti-inflammatory drug in farm animals.
The use of knockout mice as a first step in the study of in vivo interaction with drug transporters has been validated (Giacomini et al., 2010), and it is a widely used model to test the in vivo relevance of some transporters such as P-glycoprotein or ABCG2 in drug pharmacokinetics and secretion into milk (Vlaming et al., 2009). Therefore, as a preclinical setup, our results with knockout mice show that Abcg2 is implicated in the secretion of FLU and 5OH-FLU into milk with higher secretion in wild-type mice than in Abcg2(−/−) mice (Fig. 3). The effect of compensatory changes in the expression of proteins that participate in metabolism in the Abcg2(−/−) mice cannot be completely excluded. However, evidence of such problematic effects in previous studies has never been shown.
With this outcome, we have added the anti-inflammatory FLU and its main metabolite 5OH-FLU to the list of compounds with Abcg2-mediated secretion into milk. These results are in accordance with our in vitro findings, which report that FLU and 5OH-FLU were also effectively transported by murine and bovine ABCG2 (Figs. 1 and 2). The very high relative transport ratio in the case of murine Abcg2-mediated transport of FLU is noteworthy (34.00 ± 7.44). All these data suggest that ABCG2 active transport is involved in the secretion of these compounds into milk.
Regarding the effect of the bovine Y581S ABCG2 polymorphism in the secretion into milk of these compounds, a pharmacokinetic study using cows carrying this polymorphism was designed. Our results confirmed that the predominant compound detected in plasma was FLU (Odensvik and Johansson, 1995; Jedziniak et al., 2007; Kissell et al., 2015). The plasma pharmacokinetics parameters (Table 1) in both groups of lactating cows were similar to those obtained in previous studies in dairy cattle (Rantala et al., 2002; Jedziniak et al., 2007). However, no significant differences in both groups of animals were obtained for FLU and 5OH-FLU in the plasma levels and pharmacokinetics parameters (Fig. 4; Table 1).
Moreover, our data regarding milk levels are in good agreement with previous publications that describe 5OH-FLU as the main FLU metabolite presented into milk (Feely et al., 2002; Daeseleire et al., 2003; Ngoh et al., 2003; Kissell et al., 2012, 2015; Jedziniak et al., 2013). A large amount of 5OH-FLU appeared rapidly in cow milk following administration of FLU. 5OH-FLU milk levels presented a fast rate of elimination after 24 hours of treatment (Ngoh et al., 2003; Jedziniak et al., 2009, 2013; Kissell et al., 2012, 2015). This decrease is more pronounced in the intravenous route than in other routes (Kissell et al., 2012). We obtained significant differences between both groups of cows in milk secretion of the parental drug FLU at 6, 14, and 24 hours post-treatment and in its milk pharmacokinetics parameters Cmax and AUC0–∞, with 1.5-fold higher levels in cows carrying the polymorphism than in noncarrier animals (Fig. 5; Table 2). In addition, the mean residence time elimination parameter significantly increased in the Y/S581 heterozygous animals compared with the Y/Y581 homozygous animals. These results suggest that milk levels of FLU were higher in the Y/S581 animals and that drug persistence in the milk may be longer, indicating that higher concentrations of this NSAID are secreted in the Y/S581 animals for a longer period of time. These results are in good agreement with previous studies by our group in which we demonstrated higher secretion into milk of several antibiotics such as danofloxacin, ciprofloxacin, and enrofloxacin by cows carrying the Y581S polymorphism (Otero et al., 2013, 2015, 2016). However, this is the first time that such an effect has been observed for an anti-inflammatory drug. This contrasts with our results in plasma where no effect of the Y581S polymorphism was observed, probably due to the potential influence of several additional mechanisms at the systemic level. In fact, Miyazaki et al. (2001) and Horii et al. (2004) reported that the OATP-2 active membrane transporter was involved in the bioavailability and biodisposition of FLU in rabbits and cats. However, ABCG2 is the only ATP-binding cassette transporter induced in the mammary gland during lactation (Jonker et al., 2005). Organic cation transporters have also been related to drug transfer in mice (Ito et al., 2014). Otero et al. (2013) also reported that no significant differences were found in plasma levels between Y/Y581 and Y/S581 cows after danofloxacin administration at 1.25 mg/kg b.wt., although these authors observed higher milk secretion of this antimicrobial in cows carrying the Y581S polymorphism than in noncarriers.
The difference observed between the two bovine genotypes regarding secretion of FLU into milk is in agreement with our in vitro data, where significant differences in the relative transport ratio of FLU at 4 hours were found between cells transduced with Y581 and S581 variants, with higher apically directed transport in the polymorphic variant (Fig. 1, C and D). However, no differences in the basolateral-to-apical transport of 5OH-FLU between the two bovine ABCG2 variants were observed (Fig. 2, C and D), which is in agreement with the lack of difference in the secretion of 5OH-FLU into milk between the two groups of cows (Fig. 5). All these data indicate that in vitro studies using MDCK-II cells are strong and useful tools to research and predict in vivo results. Previous studies have reported the use of in vitro systems for prediction and monitoring of the concentration of drug residues in ruminant milk mediated by ABCG2 (Real et al., 2011; Wassermann et al., 2013; González-Lobato et al., 2014).
Our results show that ABCG2 is implicated in the secretion of FLU and 5OH-FLU into milk. This knowledge will help to understand and manage the presence of anti-inflammatory drugs and their metabolite residues in milk. To the best of our knowledge, although there is no available information about the pharmacological activity of 5-OH-FLU, food residues of these compounds are deemed a potential risk to consumers and maximum residue limits for them in food products of animal origin, including milk, have been established (European Medicines Agency, 2000; Food and Drug Administration, 2004).
Apart from the genetic Y581S polymorphism, several factors influence the activity of ABCG2, including gender, diet compounds, and coadministration with drugs that interact with the transporter. In fact, ABCG2-mediated secretion of drugs into milk can be greatly diminished by administration of ABCG2 inhibitors present in the diet such as flavonoids or lignans. This effect has been shown with ABCG2 substrates such as fluoroquinolones and nitrofurantoin in ruminants. This antimicrobial secretion into milk was decreased using flavonoids (soy-enriched diet) or lignans (flaxseed-enriched diet) (Pulido et al., 2006; Perez et al., 2013; Otero et al., 2018). Regarding drug modulation of milk residues, the milk secretion of the antimicrobial danofloxacin was reduced by coadministration of the macrocyclic lactone ivermectin, an ABCG2 inhibitor (Real et al., 2011). Moreover, the coadministration of the anthelmintic triclabendazole, another ABCG2 inhibitor, with moxidectin reduced milk levels of this macrocyclic lactone (Barrera et al., 2013). Therefore, active transport of FLU and its main metabolite into milk by ABCG2 may have implications in the coadministration of FLU with other drugs or natural feed components that also interact with ABCG2 in the treatment of dairy cows because it may affect milk levels of drug residues with an important health risk for human consumption.
Our findings are a new step toward the identification of factors that alter drug exposure in livestock and are involved in interindividual variability. This newly discovered factor involved in interindividual disposition variability of anti-inflammatory drugs could potentially affect treatment efficacy and contribute to milk residues.
In conclusion, we demonstrate that ABCG2 is involved in the secretion into milk of FLU and 5OH-FLU and that the bovine Y581S variant increases the levels of FLU in milk but does not affect 5OH-FLU milk levels. Furthermore, our results obtained from the pharmacokinetics and milk secretion studies with mice and dairy cattle are in agreement with our in vitro assays.
Acknowledgments
We thank Dr. A.H. Schinkel (The Netherlands Cancer Institute, Amsterdam, The Netherlands), who provided parental MDCKII cells its murine Abcg2-transduced subclones. The authors thank Virginia Medina (Laboratorio de Tecnicas Instrumentales, University of Leon) for technical assistance and support. We thank James McCue for help in language editing.
Authorship Contributions
Participated in research design: Garcia-Mateos, Alvarez, Merino.
Conducted experiments: Garcia-Mateos, Garcia-Lino, Alvarez-Fernandez, Blanco-Paniagua, de la Fuente.
Performed data analysis: Garcia-Mateos, Merino.
Wrote or contributed to the writing of the manuscript: Garcia-Mateos, Alvarez, Merino.
Footnotes
- Received November 12, 2018.
- Accepted March 7, 2019.
This work was supported by the Spanish Ministry of Economy and Competitiveness and the European Regional Development Fund (MINECO/FEDER, UE) [Project AGL2015-65626-R]; the Spanish Ministry of Education, Culture and Sport [Pre-Doctoral Fellowship FPU14/05131] (to D.G.-M.); the Ministry of Economy and Competitiveness [Pre-Doctoral Fellowship BES-2016-077235] (to A.M.G.-L.); and the Junta de Castilla y Leon and European Regional Development Fund [Post-Doctoral Fellowship LE011P17] (to I.A.-F.).
Abbreviations
- 5OH-FLU
- 5-hydroxyflunixin
- AUC0–∞
- area under the curve from zero to infinity
- FLU
- flunixin
- LOD
- limit of detection
- LOQ
- limit of quantification
- MDCK
- Madin-Darby canine kidney
- NSAID
- nonsteroidal anti-inflammatory drug
- Copyright © 2019 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}