


Abstract
Many drugs prescribed to children are drug transporter substrates. Drug transporters are membrane-bound proteins that mediate the cellular uptake or efflux of drugs and are important to drug absorption and elimination. Very limited data are available on the effect of age on transporter expression. Our study assessed age-related gene expression of hepatic and intestinal drug transporters. Multidrug resistance protein 2 (MRP2), organic anion transporting polypeptide 1B1 (OATP1B1), and OATP1B3 expression was determined in postmortem liver samples (fetal n = 6, neonatal n = 19, infant n = 7, child n = 2, adult n = 11) and multidrug resistance 1 (MDR1) expression in 61 pediatric liver samples. Intestinal expression of MDR1, MRP2, and OATP2B1 was determined in surgical small bowel samples (neonates n = 15, infants n = 3, adults n = 14). Using real-time reverse-transcription polymerase chain reaction, we measured fetal and pediatric gene expression relative to 18S rRNA (liver) and villin (intestines), and we compared it with adults using the 2−∆∆Ct method. Hepatic expression of MRP2, OATP1B1, and OATP1B3 in all pediatric age groups was significantly lower than in adults. Hepatic MDR1 mRNA expression in fetuses, neonates, and infants was significantly lower than in adults. Neonatal intestinal expressions of MDR1 and MRP2 were comparable to those in adults. Intestinal OATP2B1 expression in neonates was significantly higher than in adults. We provide new data that show organ- and transporter-dependent differences in hepatic and intestinal drug transporter expression in an age-dependent fashion. This suggests that substrate drug absorption mediated by these transporters may be subject to age-related variation in a transporter dependent pattern.
Introduction
Drug transporters are membrane-bound proteins whose primary function is to facilitate the trafficking of drugs and their metabolites across a biologic membrane. Expressed in structural cells that compose the organs of importance to drug disposition, such as enterocytes and hepatocytes, they are involved in the active uptake and elimination of orally administered drugs. In adults, drug transporters are recognized as key determinants of variation in the pharmacokinetics of many drugs, as shown in studies using primary cell and ex vivo organ cultures as well as clinical studies (DeGorter et al., 2012). In contrast, such data in children are scarce, and clinical studies are absent (Chen et al., 2005, 2008; Fakhoury et al., 2005; Miki et al., 2005; Fukudo et al., 2006; Mizuno et al., 2014).
Pharmacokinetics in children are known to be affected by developmental changes with age (Kearns et al., 2003). Drug metabolizing enzyme activity, for example, changes significantly from fetal to adolescent age associated with alterations in metabolic clearance of many drugs (de Wildt, 2011). Notably, hepatic and intestinal expression of cytochrome P450 subfamily CYP3A4 shows a clear developmental pattern, and systemic clearance of midazolam, a known substrate of CYP3AA, changes with age (Johnson et al., 2001; de Wildt et al., 2002; Leeder et al., 2005). Similarly, compared with adults, differences in body composition and renal function in children affect drug tissue distribution and renal drug excretion, respectively.
As a consequence, it has become apparent that simply extrapolating drug doses from adults to children based on body weight is likely to be inaccurate owing to our lack of understanding of age-dependent differences in the maturation of the drug absorption and elimination processes involved in drug disposition. As the expression of drug metabolizing enzymes appears to be age-related, it is also likely that this is the case for drug transporter expression. Data from animal studies report the occurrence of transporter-specific maturation (Klaassen and Aleksunes, 2010). In humans, several efflux transporters that belong to the ATP-binding cassette family (ABC), including multidrug resistance protein 1 (MDR1/ABCB1), multidrug resistance-associated protein 2 (MRP2/ABCC2), and breast cancer resistance protein (BCRP/ABCG2), were found to be expressed in fetal liver or intestine, but otherwise little is known of the developmental patterns of individual drug transporters (Chen et al., 2008; Fakhoury et al., 2009; Sharma et al., 2013). Systematic characterization of age-related differences in transporter expression will aid the selection of the most appropriate dose for substrate drugs in children.
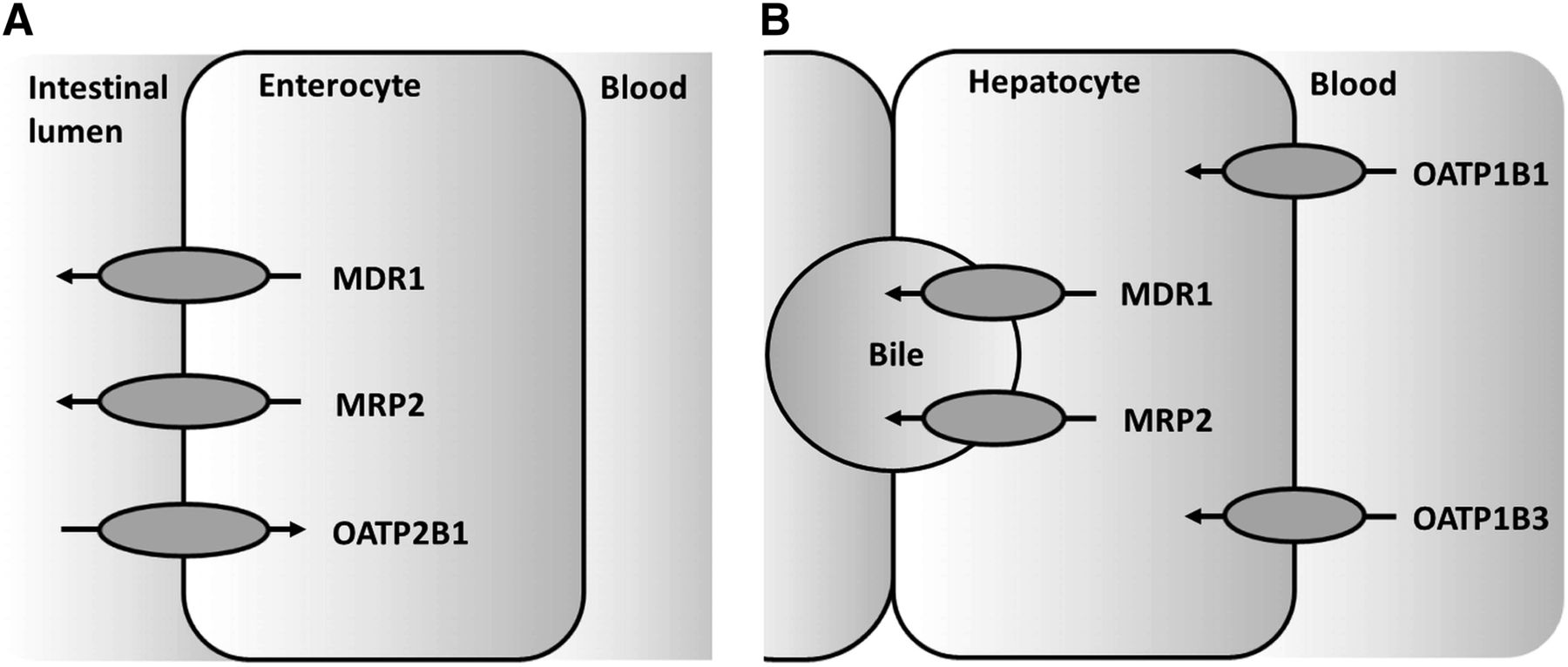
In a first step toward elucidating the developmental changes, this study determined the gene expression of clinically relevant hepatic and intestinal drug transporters across the pediatric age range. We focused on transporters with well-defined roles in drug pharmacokinetics in adults (DeGorter et al., 2012) (Fig. 1). These included the efflux carriers MDR1 and MRP2 in liver and intestine, and uptake carriers of the organic anion transporting polypeptide (OATP/solute carrier organic anion [SLCO]) family, including the liver-specific transporters OATP1B1 and OATP1B3, and OATP2B1 in the intestine.

Transporters in enterocytes and hepatocytes. Only transporters in (A) enterocytes and (B) hepatocytes were selected for this study.
Materials and Methods
Tissue Samples.
Autopsy liver tissue samples from fetuses and from children up to 18 years of age were obtained from the Erasmus MC Tissue Bank. An opt-out clause was in place for use of tissue from the tissue bank. The Erasmus MC research ethics board provided a waiver for ethics approval according to the Dutch law on research in humans. Fresh intestinal tissue was collected during surgery at the time of resection. Intestinal samples from children were derived from other research projects, and informed consent was previously obtained for the use of this tissue in the context of these studies approved by the Erasmus MC research ethics board. After resection, all liver and intestinal tissues were immediately snap frozen in liquid nitrogen, stored at −80°C, and processed on ice for mRNA isolation.
Healthy human adult liver samples were obtained from the Liver Tissue Cell Distribution System at the University of Minnesota under National Institutes of Health (NIH) contract N01-DK-7-0004/HHSN267200700004C. Adult intestinal samples were obtained from the University of Western Ontario, and the use of these samples was approved by the research ethics board.
Additional pediatric tissue specimens for MDR1 analyses were obtained from NIH-supported tissue programs: the Liver Tissue Cell Distribution System (LTCDS) from the Minnesota and Pittsburgh collection centers; the University of Maryland Brain and Tissue Bank for Developmental Disorders (Baltimore, MD); and the Laboratory of Developmental Biology at the University of Washington (Seattle, WA). Additional samples were obtained from Xenotech, LLC (Lenexa, KS). The use of these tissues was declared nonhuman subjects research by the Children’s Mercy Hospitals and Clinics pediatric institutional review board.
The sources of the obtained samples are listed in Table 1.
Tissue sources and age distribution
mRNA Isolation and cDNA Synthesis.
Frozen human tissue samples were mechanically homogenized on ice. Homogenate was applied to a QiaShredder column (Qiagen, Valencia, CA), and RNA was extracted using the RNeasy Mini Kit (Qiagen) according to the manufacturer’s instructions. RNA was treated with DNase to digest genomic DNA remnants. The quantity and integrity of RNA were determined by microfluidics-based analyses using the 2100 BioAnalyzer (Agilent, Santa Clara, CA). Samples with an RNA integrity number (RIN) of <5 were excluded from this study. Complementary DNA (cDNA) was synthesized using the High-Capacity cDNA Archive Kit (Applied Biosystems, Foster City, CA) and random priming.
Real-Time Reverse-Transcription Polymerase Chain Reaction.
The mRNA expression was measured by SYBR green quantitative real-time polymerase chain reaction with the ABI 7500 sequence detection system (Applied Biosystems). Previously optimized primers were used (Supplemental Material) (Glaeser et al., 2007). Modified primers were used for villin and OATP2B1 (reverse primer only), and the primer sequences were obtained via the qPrimer Depot (http://primerdepot.nci.nih.gov/). After each polymerase chain reaction, a melting curve analysis was performed to confirm product specificity. Eukaryotic 18S rRNA was assessed as the endogenous control with the use of a TaqMan VIC/MGB probe (4319413E; Applied Biosystems). Transporter transcript levels were normalized to 18S rRNA, and relative expression was determined using the 2−ΔΔCt method comparing pediatric samples to adult liver or intestine (calibrator samples) (Schmittgen and Livak, 2008). The adult target gene value was determined by using the median of the adult threshold cycle (Ct) values.
For comparison, all 96-well plates analyzing liver samples included a specific liver control sample to normalize expression for the gene of interest and 18S rRNA, if appropriate. Similarly, a specific intestinal sample was added to plates to normalize for the genes of interest, 18S rRNA, and villin if appropriate.
Statistics.
All data were analyzed by Kruskal-Wallis analysis, followed by group comparisons using a nonparametric Mann-Whitney test and post-hoc Bonferroni multiple comparison test. All statistical analyses were performed using GraphPad Prism version 5.00.2 and IBM SPSS Statistics software (SPSS Statistics for Windows, version 21.0; IBM, Armonk, NY). P < 0.05 was considered statistically significant. Data are presented as median and range, unless otherwise indicated.
Results
Sample Characteristics.
Characteristics of all samples are provided in Table 1. Forty-one liver samples from anonymous donors were collected from the Erasmus MC Tissue Bank. Seven samples were excluded because of poor mRNA quality, with 34 samples remaining for analysis. The causes of death were the following: congenital disorders (n = 9) [including hydrops fetalis (n = 2), ruptured giant omphalocele, hemangioendothelioma, extended congenital abnormalities, trisomy 7 or 8, cardiac, osteogenesis imperfecta], cardiac failure (n = 6), respiratory disorders (n = 6), sepsis (n = 4), gastrointestinal disorders (n = 4), neurologic disorders (n = 3), and unknown causes (n = 2). Adult liver samples (n = 11) were derived from histologically normal parts of transplanted livers. Death was related to anoxia (n = 5), trauma (n = 3), neurologic disorders (n = 2), or unknown cause (n = 1), and ages were unknown. The RIN median of fetal, pediatric, and adult liver samples was 7 (range 5–10). The MDR1 mRNA expression was also measured in additional pediatric liver samples: three fetuses, 1 neonate, 4 children aged 1 month to 12 months, and 17 children aged 12 months to 17 years. Thus, the total set of liver samples in which MDR1 mRNA expression was measured consisted of 9 fetal samples and 52 neonatal/pediatric samples. Cause of death in the additional pediatric donors was unknown.
Twenty-eight intestinal resection samples were obtained, of which 10 were excluded due to poor mRNA quality. The main reasons for resection were necrotizing enterocolitis and intestinal atresia (n = 11); other reasons (n = 7) were volvulus, persistent ductus omphaloentericus, Meckel’s diverticulum, meconium peritonitis, and ileostomy closure. Resection areas were aimed at the most proximal part of the intestine. Adult intestinal samples (n = 14) were endoscopic biopsy samples and negative for any pathology. RIN median of pediatric intestinal samples was 7 (range 5–9). The RIN values of samples from adult intestinal donors were not measured. In pediatric and adult intestinal samples, 18S was significantly correlated with villin mRNA (n = 32, r=0.6110, P < 0.05). We also determined CYP3A4 gene expression. The mRNA expression of CYP3A4 is well reported in the literature. Patterns of intestinal and hepatic CYP3A4 mRNA expression were the same as those reported in the literature (data not shown) (Miki et al., 2005).
Information about medication use by the donors was not available.
Transporter-Specific Ontogenetic Profile of MDR1, MRP2, OATP1B1, and OATP1B3 mRNA Expression in Liver.
Hepatic transporter expression was significantly associated with age: MDR1 H(4) = 35.3, P < 0.05; MRP2 H(4) = 18.0, P < 0.05; OATP1B1 H(4) = 27.4, P < 0.05; OATP1B3 H(4) = 28.1, P < 0.05. Fetal, neonatal, and infant (up to 12 months of age) gene expression of all hepatic transporters was lower than in adults. In the age group 1 to 7 years, only two samples were assessed for the hepatic transporters MRP2, OATP1B1, and OATP1B3, as a result of which median mRNA expression could not be determined and comparison with adult expression was not possible.
MDR1 mRNA expression in fetal [0.061 (0.027–0.792)] and neonatal [0.036 (0.011–0.364)] age groups was 20- to 30-fold lower than in adults (Fig. 2). MDR1 mRNA expression in infants was slightly lower [0.248 (0.010–1.46)] than in adults (Fig. 2). Median MDR1 gene expression in children aged 1 to 7 years was not different from adults [0.777 (0.012–8.928); P = 0.663]. Interindividual variability in all age groups was large.
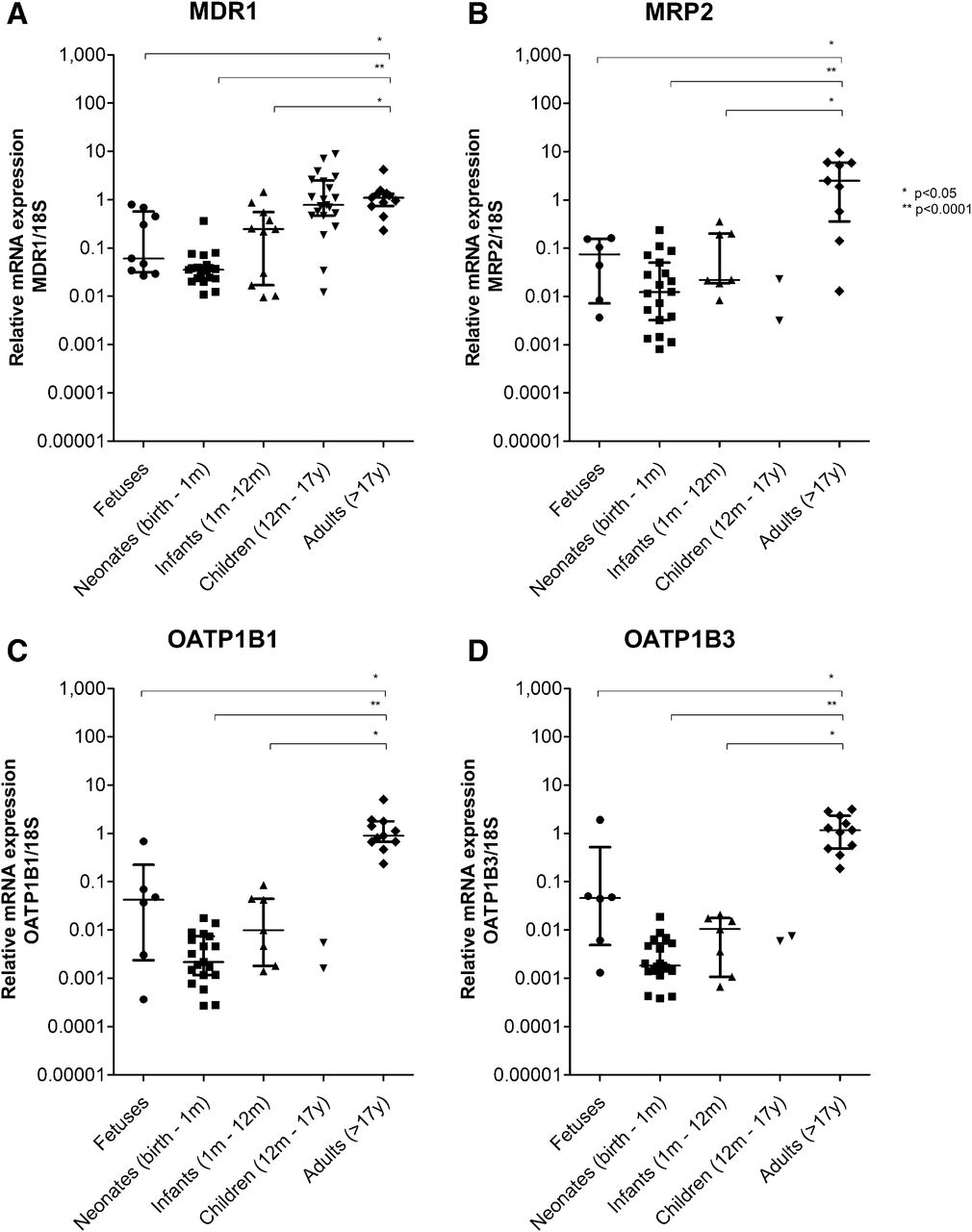
Transporter gene expression in liver. Relative mRNA expression of (A) MDR1, (B) MRP2, (C) OATP1B1, and (D) OATP1B3 from fetal, pediatric, and adult liver samples was normalized to 18S mRNA expression and adult expression using the 2−ΔΔC method. Lines represent median and interquartile range.
MRP2 mRNA expression was about 30-fold lower in fetal liver samples [0.074 (0.004–0.162)], 200-fold lower in neonates [0.012 (0.001–0.238)], and 100-fold lower in infants [0.022 (0.008–0.357)] than in adult liver samples (Fig. 2). Compared with adult liver, OATP1B1 mRNA expression was 20-fold lower in fetal liver samples [0.042 (0.0004–0.683)], 500-fold lower in neonates [0.002 (0.0003–0.018)], and 90-fold lower in infants [0.010 (0.001–0.086)] (Fig. 2). Expression profiles of OATP1B3 in the liver samples were similar to those of OATP1B1. Expression of hepatic OATP1B3 was 30-fold lower in fetuses [0.046 (0.001–1.910)], 600-fold lower in neonates [0.002 (0.0004–0.0187)], and 100-fold lower in infants [0.010 (0.001–0.021)] than in adults (Fig. 2).
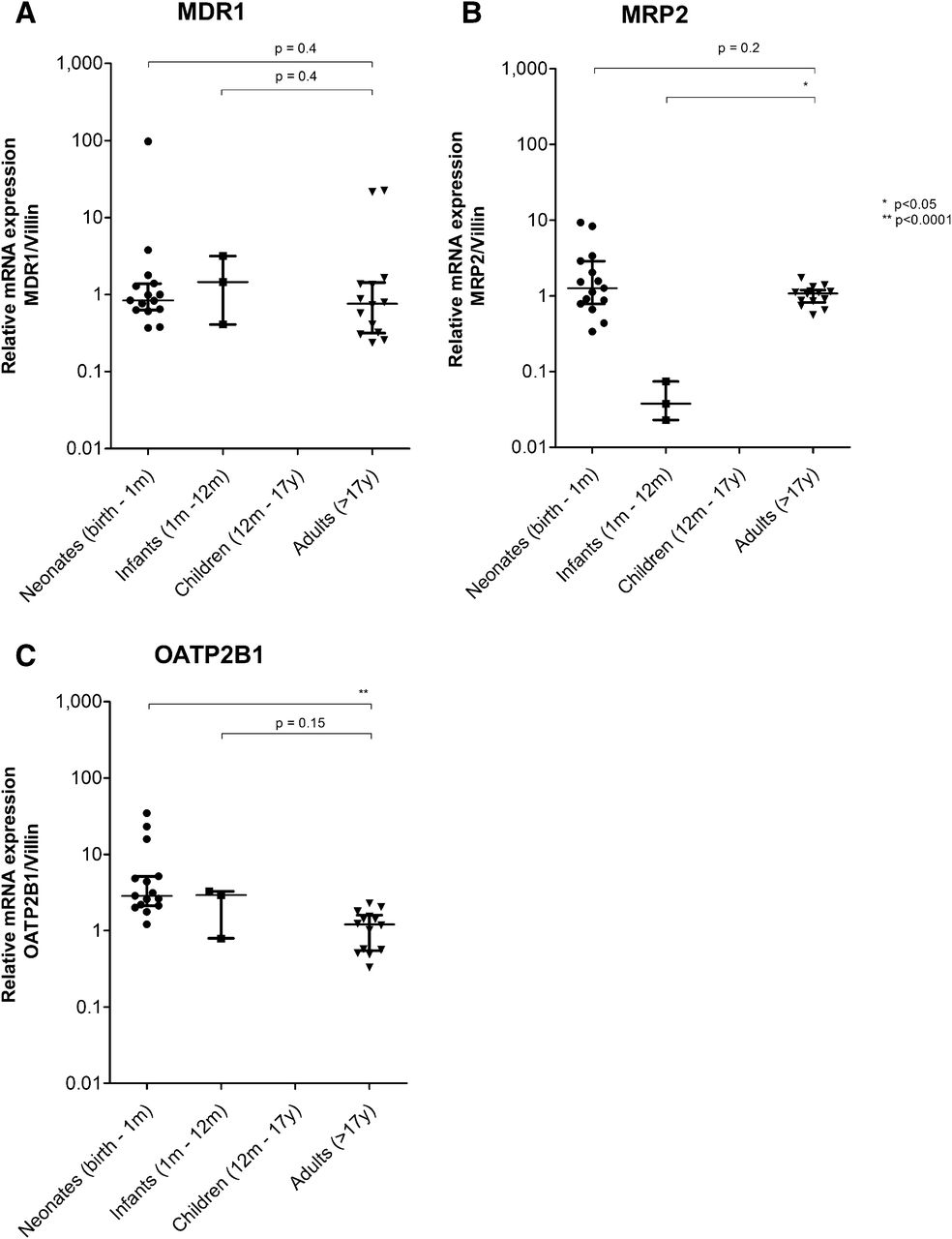
Similar MDR1 and MRP2 mRNA Expression in Neonatal and Adult Intestines, and Higher OATP2B1 in Neonatal Intestines.
Intestinal transporter expression was statistically significantly associated with age for MRP2 H(2) = 9.1, P < 0.05 and OATP2B1 H(2) = 15.8, P < 0.05, but not for MDR1, H(2) = 1.3, P = 0.5. MDR1 expression in intestinal samples of neonates was 0.838 (0.370–97.0; P = 0.395) and of infants was 1.454 (0.409–3.174; P = 0.413) and hence similar to adults (Fig. 3). Expression of MRP2 mRNA in the intestinal samples of neonates [1.262 (0.338–9.266); P = 0.248] was similar to the adult expression (Fig. 3). In infant intestinal samples, however, MRP2 mRNA expression was statistically significantly lower [0.038 (0.023–0.074); P < 0.05] than in adult samples. Interestingly, the uptake transporter OATP2B1 showed a higher mRNA expression in neonates [2.86 (1.212–34.45); P < 0.0001]. In infants up to 12 months of age, OATP2B1 was expressed at the same level as in adult intestinal samples [1.21 (0.328–2.288); P = 0.147] (Fig. 3).

Transporter gene expression in intestine. Relative mRNA expression of (A) MDR1, (B) MRP2, (C) OATP2B1 from pediatric and adult intestine samples was normalized to villin mRNA expression and adult expression using the 2−ΔΔC method. Lines represent median and interquartile range.
Discussion
Little is known about transporter gene expression in children. Our study used samples from fetal to adult age to examine the gene expression of hepatic transporters MDR1, MRP2, OATP1B1, and OATP1B3; they were found to be age-related, as their expression was lowest in fetal and pediatric liver samples. In contrast, intestinal OATP2B1 expression in neonates was higher than in adults. The intestinal MDR1 and MRP2 expression was similar between neonates and adults.
Ontogeny in Hepatic Transporters.
Hepatic MDR1 expression is already detectable in early human fetal life. The maturation pattern found in our study is consistent with previous data on hepatic MDR1 mRNA expression (van Kalken et al., 1992; Miki et al., 2005; Fakhoury et al., 2009). Our findings are also supported by immunohistochemical staining showing that MDR1 expression followed the architectural changes during fetal development (van Kalken et al., 1992). We show that MDR1 expression changes after birth, with low mRNA expression until 12 months of age whereupon it increases to adult levels. In a recent study that used a novel liquid chromatography with tandem mass spectrometry method to estimate protein expression, from 7 years onward MDR1 was stable up to 70 years of age in a cohort of more than 50 patients (Prasad et al., 2014).
Low fetal MRP2 expression in humans has been reported earlier in two small studies (n = 10, 14–20 weeks’ gestational age; and n = 3, 20–21 weeks’ gestational age) (Chen et al., 2005; Sharma et al., 2013). In the latter, however, two of five control adult liver samples were from normal tissue in patients with hepatocellular carcinoma. Because organic cation transporter 1 (OCT1) expression has been shown to be lower in nontumor hepatocellular carcinoma liver than healthy liver, a similar phenomenon with other transporters cannot be excluded (Schaeffeler et al., 2011). Using immunohistochemical staining, distinct canalicular expression of MRP2 in adult liver was observed but a fuzzier canalicular occurrence in fetal liver, which may suggest immaturity of the localization pattern (Chen et al., 2005). In addition, fetal human liver tissues obtained at 19 weeks’ gestational age showed higher MRP2 signal intensity than tissues obtained at 14 weeks (Cizkova et al., 2005). In our study, neonatal MRP2 gene expression was lower than in adults. From 7 years onward, MRP2 protein expression was stable (Prasad et al., 2013).
The observed lower fetal OATP1B1 and OATP1B3 mRNA expression levels in the present study are also in line with human data derived from three fetal (21–23 weeks’ gestational age) and three adult livers (Sharma et al., 2013). In our study, OATP1B1 and OATB1B3 expression was also low in neonates. In contrast, the reported neonatal OATP1B1 and OATP1B3 protein expression (n = 5) was similar in adults (Yanni et al., 2011). In 80 human pediatric liver samples ranging from age 2 months to 12 years, OATP1B1 was low until 6 years of age, whereas OATP1B3 protein levels were high at birth then rapidly decreased in the first year to then increase again from 8 years of age onward to adult levels (Thomson et al., 2013). From 7 years onward, OATP1B1 and OATP1B3 protein expression was stable, with only genotype related to OATP1B1 abundance (Prasad et al., 2014).
Ontogeny in Intestinal Transporters.
We observed stable intestinal MDR1 expression from neonatal to adult age, a finding that fills a gap between currently available fetal and pediatric data. Previously, fetal intestinal tissue obtained after induced abortion showed no or minimal MDR1 mRNA expression in samples after 11, 13, and 14 weeks, but clear expression at 16 and 20 weeks’ gestational age (van Kalken et al., 1992). Another study reported similar small intestinal MDR1 mRNA expression in fetuses (14–20 weeks’ gestational age), neonates, and adults (Miki et al., 2005). However, a limitation is the small sample size [n = 5, n = 12 (fetus plus neonates)] (van Kalken et al., 1992; Miki et al., 2005). Stable MDR1 mRNA expression in children from 1 month to adulthood was shown in nondiseased duodenal biopsy and jejunal tissue from liver donor recipients (Fakhoury et al., 2005; Mizuno et al., 2014). We also found stable intestinal MRP2 expression but higher expression levels of OATP2B1 in neonates compared with adults.
The transporter maturation we observed likely reflects the physiologic roles of these transporters in organ development. Epigenetic mechanisms such as DNA methylation are thought to be involved in the maturation of drug metabolizing enzymes. For example, DNA methylation seems to partly explain a developmental switch from expression of CYP3A7 to CYP3A4 in human fetal and adult liver (Kacevska et al., 2012). Epigenetic mechanisms may be also involved in the developmental regulation of drug transporter expression, but studies are currently lacking.
Due to mediation by transcription factors (which in their turn can be induced by chemicals or pathologic conditions), some transporters show relatively high whereas others show low expression (Klaassen and Aleksunes, 2010). For example, expression of the steroid- and xenobiotic-sensing pregnane X receptor (PXR) was shown to correlate with intestinal breast cancer resistance protein (BCRP), MRP2, and MDR1 mRNA in many different human tissues (e.g., intestine and liver) (Synold et al., 2001; Albermann et al., 2005; Miki et al., 2005). We may further speculate that the ontogeny of transporters is a combined function of developmental changes, genetic heterogeneity, and exposure to substrates (drug or environmental) that induce or inhibit expression and/or activity. For example, CYP1A2 and CYP3A4 activity, assessed by phenotyping caffeine and dextromethorphan, respectively, was shown to be affected by the type of infant feeding (breast milk or formula) (Blake et al., 2006). Dietary supplements in infant formula decreased MDR1 expression in colon cell cultures (Bebawy et al., 2009). Moreover, the effect of feeding is more variable when a child’s diet is expanded, as specific nutrients can influence MDR1 expression (Schwarz et al., 2005; Schwarz et al., 2007).
This study is one of the first exploratory studies characterizing developmental changes in hepatic and intestinal transporters. Some limitations of the data should be addressed. First, most fetal liver samples were obtained in the second trimester of gestation, thus the results might not be representative for the entire range of gestational age. Second, the variation in fetal transporter mRNA expression may be partly due to the small number of samples representing ages more distant from the median age. Given the small number per age group, the range of expression might not be representative for the entire population.
Third, all liver samples were collected within 24 hours after death, but the exact time is unknown. Still, RNA stability was minimized by RNA integrity determination.
Also, our samples may display disease-dependent variability in transporter expression. Inflammation has been shown to affect drug metabolism and transport. Decreased midazolam clearance in critically ill children is likely a result of reduced CYP3A activity (Vet et al., 2012). Inconsistency has been reported from studies assessing the effect of proinflammatory cytokines on intestinal MDR1 expression in vitro and in animal models, but they suggest a trend toward decreased expression (Fernandez et al., 2004). In children with Crohn disease, MDR1 expression was higher in both inflamed and noninflamed duodenal biopsies compared with healthy age-matched controls (Fakhoury et al., 2006). Inflammation was also shown to affect the expression of other transporters in primary human hepatocytes (i.e., OATP1B1, OATP1B3, MDR1, MRP2), resulting in reduced mRNA expression, protein, and activity (Le Vee et al., 2011). Thus, evidence exists that the potential effects of underlying disease should be considered when interpreting age-related changes in transporter expression.
Finally, transporter genetic polymorphisms likely contribute to the observed variability in gene expression (DeGorter et al., 2012). Recently, Nies et al. (2013) and Prasad et al. (2014) showed that hepatic expression of OATP1B1, but not OATP1B3, is significantly affected by genetic variants.
The observed age-dependent maturation and large interindividual variability in drug transporter expression may have implications for oral drug absorption and hepatic drug excretion. The question whether posttranscriptional modifications occur and if expressed mRNA level can be extrapolated to protein expression and ultimately in vivo activity remains to be answered (Johnson et al., 2001; Fakhoury et al., 2005). Human hepatic OATP1B1 and OATP1B3 protein expression showed a trend with high interindividual variability in the first year of life followed by lower variability until the age of 8 years, when variability increases again (Thomson et al., 2013). This typical age-related change in variability at a younger age is also reflected by our findings on OATP1B1 and OATP1B3 mRNA expression. In contrast, others could not correlate protein levels with OATP1B1 or MDR1 mRNA expression (Ulvestad et al., 2013). Further protein expression and transporter activity studies are needed to confirm these mRNA expression results.
In conclusion, hepatic MDR1, MRP2, OATP1B1, OATP1B3, and intestinal MDR1, MRP2, OATP2B1 drug transporter expression show organ- and transporter-specific maturation patterns during childhood. Therefore, the disposition of drugs substrates for these transporters may be subject to age-related changes other than that of body size alone.
Acknowledgments
The authors thank Y. Simons-Oosterhuis, S. LeMay, and M. Leon-Ponte for laboratory executive support, J. Hagoort for editing support, and Drs. N. Burger-Van Paassen, P.J. Puiman, M.W. Schaart, and A.E.C.J.M. Struijs for collecting intestinal tissue samples.
Human liver tissue was obtained from two tissue retrieval and distribution sources supported by the Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD): the Brain and Tissue Bank for Developmental Disorders at the University of Maryland, Baltimore, and the Laboratory of Developmental Biology at the University of Washington, Seattle. The role of the NICHD Brain and Tissue Bank is to distribute tissue, and therefore it cannot endorse the studies performed or the interpretation of results.
The content of this manuscript does not necessarily represent the official views of the Eunice Kennedy Shriver National Institute of Child Health and Human Development of the National Institutes of Health.
Authorship Contributions
Participated in research design: Mooij, de Koning, Schwarz, Spaans, de Wildt.
Conducted experiments: Mooij, de Koning, de Wildt.
Contributed new reagents or analytic tools: Leeder, Gaedigk, Kim, Samsom.
Performed data analysis: Mooij, Gaedigk, de Koning, de Wildt.
Wrote or contributed to the writing of the manuscript: Mooij, de Wildt, Kim, Tibboel, Van Goudoever.
Footnotes
- Received January 7, 2014.
- Accepted May 14, 2014.
This work was supported by a Novartis investigator-initiated and Canadian Institutes of Health Research grant [Grant MOP-89753]. The project entitled “Laboratory of Developmental Biology” was supported by the National Institutes of Health Eunice Kennedy Shriver National Institute of Child Health and Human Development [Grant 5R24-HD000836].
↵
 This article has supplemental material available at dmd.aspetjournals.org.
This article has supplemental material available at dmd.aspetjournals.org.

Abbreviations
- ABC
- ATP-binding cassette transporters
- Ct
- cycle threshold
- CYP3A
- cytochrome P450 family 3A
- MDR1
- multidrug resistance 1
- MRP2
- multidrug resistance protein 2
- OATP
- organic anion transporting polypeptide
- OCT
- organic cation transporter
- RIN
- RNA integrity number
- Copyright © 2014 by The American Society for Pharmacology and Experimental Therapeutics
References
DMD articles become freely available 12 months after publication, and remain freely available for 5 years.Non-open access articles that fall outside this five year window are available only to institutional subscribers and current ASPET members, or through the article purchase feature at the bottom of the page.
|





{kind=link}
{kind=link}
{kind=link}