Article Text

Abstract
Ageing is the largest risk factor for age-related macular degeneration (AMD), and soft drusen and basal linear deposits are lipid-rich extracellular lesions specific to AMD. Oil red O binding neutral lipid represents a major age-related deposition in the Bruch membrane (BrM) and the first identified druse component. Decades after these seminal observations, a natural history of neutral lipid deposition has been articulated and a biochemical model proposed. Results obtained with multiple biochemical, histochemical, and ultrastructural methods, and supported indirectly by epidemiology, suggest that the RPE secretes apolipoprotein B (apoB)-lipoprotein particles of unusual composition into BrM, where they accumulate with age eventually forming a lipid wall, a precursor of basal linear deposit. The authors propose that constituents of these lesions interact with reactive oxygen species to form pro-inflammatory peroxidised lipids that elicit neovascularisation. Here, the authors summarise key evidence supporting both accumulation of BrM lipoproteins leading to lesion formation and lipoprotein production by the RPE. The authors update their model with genetic associations between AMD and genes historically associated with plasma HDL metabolism, and suggest future directions for research and therapeutic strategies based on an oil-spill analogy.
- Age-related macular degeneration
- ageing
- retinal pigment epithelium
- Bruch membrane
- lipoproteins
- apolipoprotein B
- drusen
- basal deposits
- cholesterol
- cholesteryl ester
- degeneration
- biochemistry
- eye (tissue) banking
- pathology
- anatomy
- retina
- macula
- neovascularisation
Statistics from Altmetric.com
- Age-related macular degeneration
- ageing
- retinal pigment epithelium
- Bruch membrane
- lipoproteins
- apolipoprotein B
- drusen
- basal deposits
- cholesterol
- cholesteryl ester
- degeneration
- biochemistry
- eye (tissue) banking
- pathology
- anatomy
- retina
- macula
- neovascularisation
Introduction
The chorioretinal biology underlying the initiation and progression of age-related macular degeneration (AMD) is poorly understood, despite clinical success in abating AMD's neovascular complications.1 The lipid-rich lesions associated with AMD, basal linear deposits (BlinD) and drusen, are perhaps given less attention than is their due. Further, the tissue compartment in which these lesions arise, the Bruch membrane (BrM), undergoes a dramatic neutral lipid accumulation throughout adulthood2 that is frequently overlooked. Because these processes in BrM both set the stage for choroidal neovascularisation and harm to overlying cells, they demand our attention in the quest for improved understanding of both wet and dry AMD.
A 1990 histochemical study by Pauleikhoff et al2 indicated a dramatic, universal and previously unknown accumulation of lipids in BrM with ageing. Biochemical studies during that era confirmed the strongly age-related nature of the deposition.3 4 Three decades prior, oil red-O binding lipid was the first molecular constituent identified in drusen.5 6 Pioneering studies in the 1990s by Marshall employed BrM explants to test the concept that lipid deposition in BrM could impair the transport of compounds necessary for outer retinal health across this tissue.7–10 Within the genomic era, genes encoding lipid-related pathways were shown to be associated with modified risk for AMD, including apoE411–13 and two genes (hepatic lipase and cholesteryl ester transfer protein) historically associated with plasma HDL metabolism.14 15 How do these disparate lines of evidence fit together? Here, we update our model of how an RPE-derived lipoprotein accumulating in BrM can lead to lesion formation.16 17
Natural history of BrM lipid deposition and lesion formation
As the inner margin of the choroid, BrM is effectively a vessel wall. Because lipid deposition in vessel walls is a well-established causative agent in atherosclerotic cardiovascular disease, we looked to this arena for guidance on basic mechanisms, laboratory techniques and potential therapeutic approaches. We relied on the conceptual framework known as the Response-to-Retention hypothesis of atherosclerosis18 19 (figure 1) in establishing the natural history of this lipid deposition in BrM.

Parallel processes in AMD and atherosclerotic coronary artery disease. Both diseases involve lipoprotein retention in vessel walls. In atherosclerosis, apolipoprotein B (apoB) lipoproteins (low-density lipoprotein (LDL), chylomicron remnants) cross the vascular endothelium of large arteries, bind to proteoglycans, become modified via oxidative and non-oxidative processes, and launch numerous downstream deleterious events, including macrophage recruitment and differentiation into foam cells, cytokine release, plaque instability, neovascularisation, haemorrhage and thrombosis (the ‘Response-to-Retention’ hypothesis).18 19 Coronary artery disease is a price humans pay for the apoB lipoprotein system, which transports and delivers energy-rich lipids and lipophilic nutrients (exogenous via dietary chylomicrons, endogenous via hepatic very-low-density lipoprotein (VLDL)) to body tissues. Statins work largely by reducing plasma levels of atherogenic apoB-containing lipoprotein particles. In AMD, lipoprotein-like particles with abundant esterified cholesterol accumulate with age in the macular Bruch membrane (BrM), especially in the plane between the retinal pigment epithelium (RPE) basal lamina and the inner collagenous layer where type I choroidal neovascularisation dissects in wet AMD. In situ modifications of these lipoproteins could enable complement activation and multiple pro-inflammatory, pro-angiogenic processes. Strong evidence supports the RPE as an intraocular source of BrM lipoproteins, without definitely excluding plasma sources. Biological processes driving RPE lipoprotein assembly and secretion are presently uncertain, but lipid compositional data reviewed herein suggest that lipophilic nutrient delivery via plasma lipoproteins could be important. Details in Curcio et al.16
Using ultrastructural techniques to preserve lipids and histochemical techniques to detect esterified and unesterified cholesterol (EC, UC), we confirmed and extended the original observation that BrM lipid deposition begins by the fourth decade of life. Profiles previously called vesicles proved to be solid, space-filling particles, 60–80 nm in diameter. These first gather among fibrils of the BrM elastic layer in early adulthood.20 21 This accumulation then extends inward to fill the inner collagenous layer, eventually leading to a new layer between the inner collagenous layer and basal lamina of BrM in many older eyes that we called the lipid wall (figure 2A,C). The oil red O binding material proved to be EC, which with UC accumulates markedly in BrM, in sevenfold-higher quantities in macula than periphery.22 23 Among lipids, EC is confined exclusively to BrM, whereas UC and phospholipids additionally localise to nearby cellular membranes.24
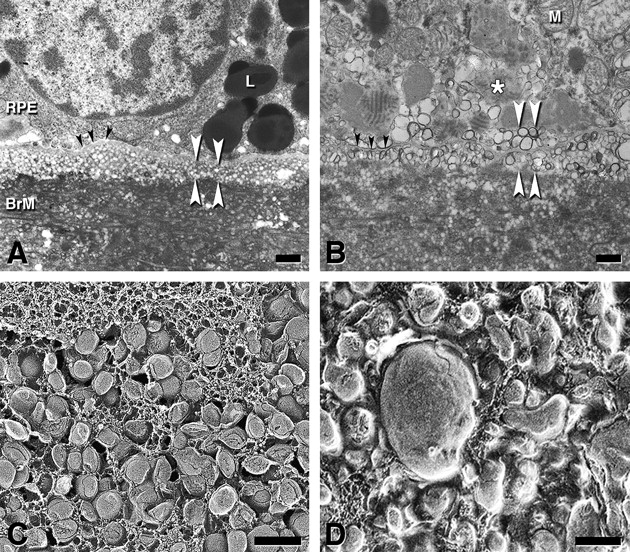
The Lipid Wall and basal linear deposits (BlinD). The Lipid Wall is a precursor to BlinD, a specific lesion of AMD. (A, B) Thin-section transmission electron micrograph showing lipoproteins as vesicles; osmium postfixation, vertical plane, bars=1 μm. (C, D) Quick freeze deep etch showing modified lipoproteins as solid particles; oblique plane, bars=200 nm. In (A), lipoproteins (spherical vesicles of uniform diameter) accumulate 3–4 deep between the retinal pigment epithelium (RPE) basal lamina (black arrowheads) and the inner collagenous layer of the Bruch membrane (BrM) (lipid wall, white arrowheads). L, lipofuscin. In (B), BlinD (white arrowheads) appears as membrane-bounded vesicles in the same plane as the lipid wall in (A). The high electron density at the vesicular borders is associated with increased unesterified cholesterol content. Black arrowheads, RPE basal lamina. M, mitochondrion. In (C), tightly packed BrM lipoproteins in the lipid wall display classic core and surface of lipoproteins.21 In (D), in BlinD from a 78-year-old donor with geographic atrophy, lipoproteins have more heterogeneous sizes and shapes. Pooled lipid is also apparent, consistent with a model of surface degradation and particle fusion.
The participation of neutral lipids in AMD-specific lesions is also now more firmly established. Because the lipid wall is in precisely the same plane as BlinD, a thin layer of soft-druse-like material specific to AMD,25 26 it is the best candidate as this lesion's precursor. Indeed, lipid aggregates interpretable as fused lipoprotein particles localise to BlinD (figure 2C,D). Further, EC and UC are present in all drusen, representing (with phosphatidylcholine) >40% of hard druse volume, the single largest druse component.27 28 Soft drusen are oily upon dissection and biomechanically more fragile than hard drusen,29 consistent with a high lipid content. Drusen contain apolipoproteins B, A-I, E, C-I, and C-II, and little plasma-abundant C-III.30 The full distribution of lipid-containing extracellular lesions in AMD has been more fully appreciated through laboratory and clinical characterisation of subretinal drusenoid debris and basal mounds.29 31 32 Next, we review evidence that this material represents lipoproteins.
Lipoproteins of RPE origin
We summarise lipoprotein basics in figure 3. In atherosclerosis, lipoproteins cross vascular endothelium of large arteries, bind to proteoglycans, become modified via oxidative and non-oxidative processes, and launch numerous downstream deleterious events, including inflammation, macrophage recruitment and neovascularisation.18 19 We examined processes occurring with ageing and in disease in BrM in this light.

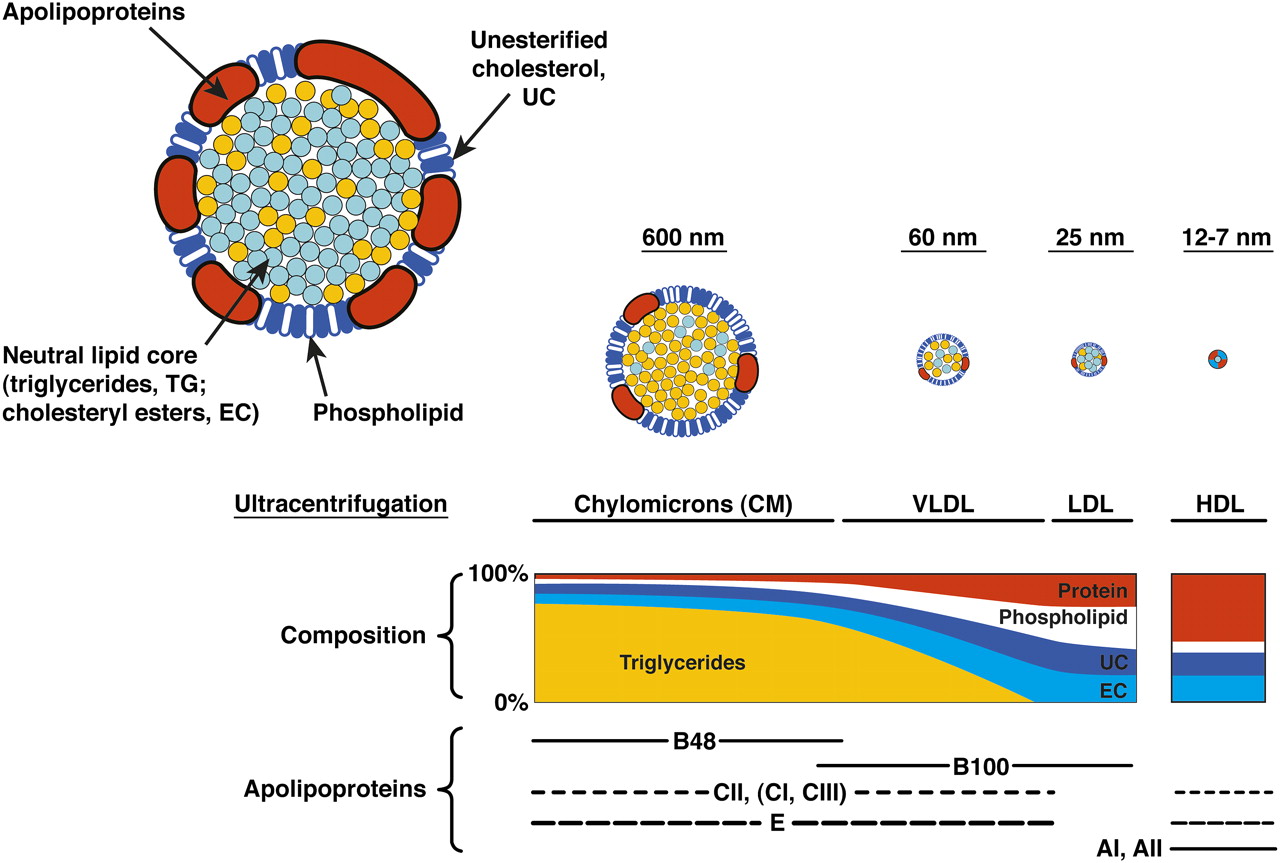
Lipoprotein basics. Lipoproteins (upper left) are multimolecular assemblies that solubilise oil droplets rich in esterified cholesterol (EC) and triglyceride (TG) for transport through an aqueous environment within a thin surface of phospholipid, unesterified cholesterol (UC), and apolipoproteins (like apoB or apoA-I). Plasma lipoproteins are the best characterised (CM, very-low-density lipoprotein (VLDL), low-density lipoprotein (LDL), high-density lipoprotein (HDL)) but lipoproteins are also secreted by brain, heart, placenta and kidney. Of the lipoprotein components, cholesterol is either unesterified (free) in membranes (UC) or esterified to a fatty acid for storage and transport (EC). UC is released from cells via several mechanisms, such as release to circulating HDL and enzymatic conversion to oxysterols. EC is released directly from cells only as part of apoB-containing lipoproteins. The neutral lipids EC and TG comprise long-chain fatty acids esterified to backbone molecules UC and glycerol, respectively. Two fatty acids of interest are linoleate (18:2n6), the most abundant in the body, and docosahexaenoate (22:6n3), especially abundant in brain and retina. Linoleate (18:2n-6) and alpha-linoleate (18:3n-3, precursor to docosahexaenoate) are not synthesised by humans and are considered dietary essential fatty acids.33 ApoB, a member of the Large Lipid Transfer Protein family, is found in a full-length form associated with hepatic VLDL (apoB-100, 512 kDa) or a shorter form associated with intestinal chylomicrons (apoB-48). VLDL is the precursor particle to LDL, popularly known as the bad cholesterol. After Vance and Vance.34
Key evidence supporting the concept of a large apoB-lipoprotein in BrM was obtained with histochemical, ultrastructural and biochemical methods. First, when viewed with lipid-preserving ultrastructural techniques, solid particles in BrM have a characteristic surface-and-core morphology resembling lipoproteins (figure 2C). Second, particles of comparable diameter with lipoprotein-like flotation properties are isolable from human BrM.35 36 Their spherical shapes indicate neutral lipid cores (figure 4A). Third, the same fractions include apoB, apoA-I and apoE. Fourth, BrM cholesterol is EC-enriched (EC/total cholesterol=0.56), as replicated by five different methods (colorimetric and fluorimetric enzymatic assay, liquid chromatography-mass spectrometry, electrospray ionisation (ESI)-MS, thin layer chromatography). Fifth, BrM neutral lipid is EC-enriched as compared with triglycerides (EC/TG=4–11), via two methods (colorimetric enzymatic assay, liquid chromatography-mass spectrometry) for both BrM and isolated lipoproteins. Of note, hepatic VLDL, of similar diameter, is TG-rich. An early report of TG-enriched BrM neutral lipid4 was not replicated.

Bruch membrane (BrM) lipoprotein. (A) Lipoprotein particles isolated from BrM. The particles are large, spherical and electron-lucent.35 Bar=50 nm. (B) BrM lipoprotein composition data from direct assay35 36 and by inference from druse composition and retinal pigment epithelium gene expression.27 30 Apo, apolipoproteins; EC, esterified cholesterol; PL, phospholipid; TG, triglyceride; UC, unesterified cholesterol. The question mark indicates that not all apolipoproteins are known.
Abundant EC points to the mechanism by which our model suggests that these lipids are released directly from cells, an apoB lipoprotein. The important question is which cells—the hepatocytes or enterocytes that make plasma lipoproteins, or cells intrinsic to the eye? Significantly, RPE expresses apoB gene and protein, along with microsomal triglyceride transfer protein (MTP), required for apoB lipidation and secretion. Lack of functional MTP is the basis of abetalipoproteinaemia, a rare inherited disorder that includes a pigmentary retinopathy.37 38 The combination of apoB and MTP within a single cell type marks the RPE as a constitutive lipoprotein secretor. A definitive experiment—secretion of full-length apoB—has been demonstrated by two laboratories using metabolic labelling and immunoprecipitation in rat-derived and human-derived RPE cell lines.39 40 The pattern of age-related lipid deposition in BrM, whereby particles first appear in the elastic layer and fill in towards the RPE, is also consistent with an RPE origin.21
Identifying the upstream sources of BrM lipoprotein constituents is essential for understanding the biological purpose of this pathway and exploiting it to clinical advantage. A major clue to source comes from compositional studies showing that each lipid class examined to date is rich in the fatty acid linoleate and poor in docosahexaenoate (DHA). Among studies varying in sample choice, sample purity, normalisation methods and assays, those using the most highly purified samples (isolated lipoproteins36 and BrM-choroid EC41) report a high mole percentage of linoleate (>40%) and low DHA (<1%), for all lipid classes. Analysis of more heterogeneous samples (neutral lipids42 or total fatty acids for BrM43) yields a 12- to 14-fold excess of linoleate over DHA. A recent report of higher DHA (2–9%) and lower linoleate still showed excess linoleate in RPE/choroid.44 This composition strongly points away from one upstream lipid source (photoreceptor outer segments, 35% DHA in membrane phospholipids) and towards another (plasma lipoproteins, 45–55% linoleate in all lipid classes). Without knowledge of a functional RPE apoB pathway, one could easily conclude from the high linoleate content of BrM lipoproteins that they originated from liver or intestine and arrived via the choroid. Rather, we interpret these results to signify that plasma lipoproteins are major contributors upstream to an apoB lipoprotein of RPE origin. DHA paucity in ageing BrM also suggests that an AMD-relevant mouse model premised on DHA abundance may involve mechanisms active at locations other than BrM,45 or that peroxidation of even small DHA concentrations can initiate significant pathological consequences.
In contrast to our conclusions for EC fatty acids, source(s) of BrM UC are not yet known and could be outer segments, plasma lipoproteins, endogenous synthesis or a combination. Upstream UC sources cannot be inferred from compositional studies, and several pathways for UC egress exist in the RPE.46–48 These studies collectively call for reconceptualising Hogan's four-decade old idea49 that age-related BrM debris derives from outer segment disposal. Rather, the lipoprotein may be assembled from several sources, including outer segments, but also their nutrient supply system, and endogenous synthesis.
Indirect evidence for a local lipoprotein also emerges from the epidemiological literature. A relationship between AMD and the concentration of plasma cholesterol or apolipoproteins has been sought repeatedly since the early 1960s,50 using measures of both atherogenic (apoB, LDL) and antiatherogenic (apoA-I, HDL) lipoproteins. If the cholesterol deposition in BrM and AMD-associated lesions were a manifestation of systemic atherosclerosis, then a strong positive correlation between disease status and plasma lipoprotein levels, like that documented for coronary artery disease,51 might be expected. None has yet emerged. Perhaps also for this reason, short-term systemically delivered statins, which act largely by reducing plasma cholesterol levels, have not been shown to appreciably benefit AMD patients.52
Oil spill: a pathobiology model
Figure 5 proposes an oil-spill time line from biology to pathobiology, based on the Response-to-Retention hypothesis and incorporating the lipid wall seen in our studies of ageing BrM. Parts of this schema are speculative, but every step offers testable hypotheses. Just as atherosclerosis begins with apoB-mediated fuel and lipophilic vitamin delivery to body tissues, we propose that an apoB-lipoprotein participates in a nutritional and recycling system serving the RPE and photoreceptors (numbered bullets refer to figure 5).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Oil spill in the Bruch membrane (BrM) from biology to pathobiology. See text for references at each step of this hypothesis. From top to bottom, photoreceptor outer segments, retinal pigment epithelium (RPE), BrM and choriocapillaris are depicted. RPE cells contain nuclei and mitochondria (magenta). Lipofuscin is omitted for clarity. (1) Plasma low-density lipoprotein (LDL) and high-density lipoprotein (HDL) delivering lipophilic nutrients are taken up at LDL and scavenger receptor class B type I (SR-BI) receptors on the basolateral RPE. (2) Proteins classically associated with plasma HDL metabolism are expressed in the subretinal space and may be involved in rapid turnover of unesterified cholesterol (UC) in the neurosensory retina. Docosahexaenoate (DHA) is cycled between RPE and retina. (3) A basolaterally secreted apolipoprotein (apo)B lipoprotein is assembled from multiple sources, including uptake of plasma lipoproteins, endogenously synthesised lipids and/or photoreceptor degradation products. Fatty acids (FA), especially linoleate (18:2n6), come largely from taken-up plasma lipoproteins. UC, from as-yet undetermined sources, is re-esterified (not shown). BrM lipoproteins also contain retinyl ester. (4) RPE expresses both apoB and microsomal triglyceride transfer protein and secretes esterified cholesterol (EC)-rich particles into BrM (gold circles), where they are retained and eventually cleared through the choriocapillary endothelium. (5) Lipoprotein particles begin to accumulate during adulthood for unknown reasons, building up a layer 3–4 deep external to the RPE basal lamina (the lipid wall). Other outflow pathways for UC include apoE, ATP-binding cassette A1 (ABCA1)-mediated transfer to circulating HDL, apoE secretion and enzymatic conversion to oxysterols (27-COOH). (6) Over time, perhaps due to reactive oxygen species from neighbouring mitochondria, pro-inflammatory/toxic species such as linoleate hydroperoxide and 7-ketocholesterol appear. Particles could fuse to form lipoprotein-derived debris, the principal component of basal linear deposits and soft drusen. Translocation of necessary compounds from plasma is impeded (blocked arrows). (7) Inflammation is elicited in the inner BrM. (8) Neovascularisation (type I) ensues. CETP, cholesteryl ester transfer protein; 7-KCh, 7-ketocholesterol; LIPC, hepatic lipase.
Plasma lipoproteins are vehicles for delivery of lipophilic nutrients (carotenoids,53 vitamin E and cholesterol54) to photoreceptors by RPE. RPE receptors for plasma LDL and HDL are functional in vitro55 56 and in vivo.57 58 Nutrients are stripped from these lipoproteins by the RPE for delivery to the photoreceptors.
Proteins known from plasma HDL metabolism localise in the subretinal space and are thought to constitute a rapid UC supply route to the retina, perhaps via HDL-like particles.54 57 Classic studies indicated that DHA is recycled back to photoreceptors,59 and dietary DHA does not influence retinal DHA content in humans.41
BrM lipoproteins are co-translationally assembled from multiple lipids, like other apoB lipoproteins. Excess UC is cytotoxic, and regardless of source, must be off-loaded by RPE. UC is re-esterified, perhaps via the enzyme ACAT-2 (60, not shown) as in other lipoprotein-secretors, to provide the abundant EC in BrM. Fatty acids, especially linoleate, could come from taken-up LDL and HDL. Isolated lipoprotein particles contain retinyl ester, possibly a part of a postulated retinoid efflux pathway.61 The RPE may thus have two recycling mechanisms—DHA from outer segments is recycled back to the photoreceptors apically, and lipoproteins from plasma are reassembled and recycled back to plasma basolaterally. At the same time, other mechanisms to offload UC (apoE, ATP-binding cassette A1 (ABCA1)-mediated transfer to circulating HDL and enzymatic conversion to oxysterols) are present in the RPE.46 48 62
Clearance of lipoprotein particles across BrM presumably occurs by diffusion, and across the choriocapillary endothelium and into plasma by transcytosis. However, the accumulation of highly hydrophobic EC beginning in early adulthood can account quantitatively for the observed increased resistivity to hydrophilic compounds transported across BrM to the RPE.63 The participation of smaller plasma lipoproteins (11 nm HDL, 22 nm LDL) in forming this transport barrier cannot be excluded; however, such particles were not noted ultrastructurally in BrM.21
Lipoproteins captured in BrM over time eventually lead to the formation of the lipid wall external to the RPE basal lamina, an ‘oil spill’ on the inner surface of aged BrM.
Over time, modified species with harmful properties, like linoleate hydroperoxide,43 7-ketocholesterol64 and others,65 are formed. Linoleate hydroperoxide, abundant in atherosclerotic plaque,66 is chemotactic, pro-inflammatory, and cytotoxic. Oxidative and non-oxidative processes lead apolipoprotein components to degrade, causing particle fusion and formation of lipoprotein-derived debris, a new name for membranous debris (major soft druse component25) that more accurately reflects the likely source. This ‘Response-to-Retention’ alters the lipid wall and initiates the formation of AMD-specific lesions, BlinD and drusen, final stages of the oil spill.
7,8. This scenario fits well at the front end of a hypothesis that peroxidised lipids in BrM evoke choroidal neovascularisation through inflammation,67 as confirmed in animal models.68 69
Despite lacunae requiring experimental verification, a chorioretinal Response-to-Retention hypothesis based on the oil spill in BrM is comprehensive in scope and direct, and it incorporates many observations. Lipoprotein secretion by the RPE can account for several abundant druse constituents with one mechanism, and modification of lipoproteins could plausibly account for many more druse components including those with known toxic and/or inflammatory properties. This model explains more data than other extant models of AMD lesion formation (eg, RPE debris shed into BrM70–73 or accumulation of RPE lipofuscin74). Accumulation of BrM lipoprotein-derived, pro-inflammatory lipids is the best trigger identified so far for complement activation75 late in life of individuals harbouring susceptibility alleles of CFH and other genes. The potent bio-activity of peroxidised long-chain fatty acids, including the activation of innate immunity, is well documented from the cardiovascular literature.76–78
Oil-spill strategies
Many specific strategies for handling oil spills were widely publicised during the Deepwater Horizon summer of 2010. Applying these strategies to treating a prevalent disease of older persons is ambitious but worth considering:
Remove accumulated lipids with pharmaceutical skimmers and dispersants. Such steps are pertinent to treatments like RPE transplants on a refurbished substrate. Apolipoprotein mimetics can sequester oxidised lipids avidly.79 80 Lipoprotein particles could also be released in vivo from BrM retention sites via high-salt solutions or other agents.
Modulate the flow of apoB lipoproteins from the RPE into BrM by a top kill. This could target apoB directly—for example, with apoB antisense,81 or an enzyme in lipidation pathways upstream to apoB (MTP,39 82 ACAT-1,2,60 83 stearoyl coenzyme A desaturase-184 85). Some enzymatic inhibitors that were developed as plasma lipoprotein modulators were unsuitable for systemic use, owing to associated hepatic steatosis. However, they could be confined to the eye via localised delivery systems.86 Intraocular application of statins, which directly reduce apoB secretion in addition to upregulating LDL receptor via HMG coenzyme A reductase inhibition, could also be considered.40 87
Prevent the oil spill altogether by using a bottom kill. Lipoprotein outflow into BrM could be modulated through dietary manipulation of the upstream lipid sources and lipase activity,88 fulfilling a long-anticipated potential of diet for AMD risk reduction.89–91 Alternatively, mechanisms underlying lipoprotein retention (eg, age-related changes in extracellular matrix or changes in the quantity or character of RPE-secreted lipoproteins) could be exploited to delay or prevent this age-related change. Other approaches can be contextualised within the oil-spill framework—for example, antioxidants may suppress lipoprotein modification, and complement inhibitors may blunt the effects of lipoprotein retention.
Oil-spill action items
Converting knowledge of an RPE apoB pathway into improved diagnoses, preventions and treatments for AMD patients will be best served by a comprehensive understanding of this new area of eye physiology and developing better tools and model systems. We suggest several critical questions.
In human eyes
Can linoleate hydroperoxide and apolipoproteins beyond B, A-I, and E be localised to BrM lipoproteins, thus providing a more complete picture of the particle?
What mechanisms unique to BrM extracellular matrix regulate lipoprotein retention?
In RPE cells
Can the upstream lipid sources, steps of intracellular processing and parameters of apoB secretion be determined in polarised RPE cell culture systems that permit delineation of apical and basolateral pathways?92 93
How do RPE gene expression levels, proteins secreted and mechanisms for EC-rich particle biosynthesis compare to other well-characterised lipoprotein secretors?
How does an RPE apoB pathway interact with UC transfer to HDL via ABCA1, release of oxysterol intermediates, DHA recycling and retinoid processing?
In animal models
Can the effects of local and systemic lipoproteins on outer retinal function be determined through tissue-specific knockouts of genes in these pathways?94
Can laboratory animals currently exhibiting BrM lipid deposition95 96 be induced to produce EC-enriched drusen or BlinD?
By what mechanisms are lipophilic nutrients delivered to retina?
In patient populations
Convergence of information from genomics and tissue-level gene expression on a suite of related pathways (cholesterol and lipoprotein metabolism) can be considered mutually reinforcing, as it was for complement. Genetics studies of pathways known to affect plasma apoB levels97 98 should be mined for ideas relevant to the control of RPE apoB secretion. Interpretations of recently described associations of hepatic lipase (LIPC) and cholesterol ester transfer protein (CETP) with AMD,14 15 as well as previously reported associations with ABCA199 and apoE4-conferred protection,11 should include consideration of intraocular mechanisms. Genome-wide scans can be now followed with cell- and tissue-level studies designed for hypothesis testing.
Detecting the oil spill clinically
Spectral-domain optical coherence tomography, widely used by retinal physicians, clearly shows drusenoid elevations/inclusions and thickening of RPE-associated signal100 but does not distinguish between basal laminar and BlinD, diffuse lesions conferring risk for disease progression.101 102 EC is abundant in the necrotic core of an atherosclerotic plaque and is visualisable via several modalities.103–107 Separation of previously indistinguishable lesions by lipid content may eventually be achievable in the eye.
The apoB pathway, leading to hepatic VLDL and its metabolite LDL, is one of the most intensely studied pathways in cardiovascular, diabetes and obesity research. Discovery of an apoB lipoprotein in BrM signifies that an immense knowledge base is now available to ophthalmologists wishing to pursue new approaches to AMD through modulation and imaging of the pathognomonic extracellular lesions and BrM itself. Note that cardiovascular medicine has enjoyed a century of animal models with lipid-rich arterial lesions and 20 years of worldwide clinical success with statins. The budget of the National Heart Lung and Blood Institute historically exceeds that of the National Eye Institute by fourfold. A vast wealth of experience and expertise can now be tapped for the benefit of patients with AMD.
Acknowledgments
We thank JD Messinger, DC, for preparing figure 2, and D Fisher, for graphic design.
References
Footnotes
Funding NIH grants EY06109 and EY014662, International Retinal Research Foundation, American Health Assistance Foundation, EyeSight Foundation of Alabama, Research to Prevent Blindness, Inc., Macula Vision Research Foundation, Roger Johnson Prize in Macular Degeneration Research, and Deutsche Forschungsgemeinschaft.
Competing interests None.
Patient consent Obtained.
Provenance and peer review Not commissioned; externally peer reviewed.