


Abstract
The objective of the current study was to evaluate the mechanism of absorption and metabolism of a PEGylated peptide, MRL-1 (46 kDa), after s.c. dosing in dogs and rats. Thoracic lymph duct-cannulated (LDC) dog and rat models were developed that allowed continuous collection of lymph for up to 8 days. When [3H]MRL-1 was administered s.c. to LDC dogs, ∼73% of the administered radioactivity was recovered in pooled lymph over a period of 120 hours, suggesting that lymphatic uptake is the major pathway of s.c. absorption for this peptide. In agreement with these data, the systemic exposure of radioactivity related to [3H]MRL-1 in LDC dogs was decreased proportionately when compared with that in noncannulated control dogs. After i.v. dosing with [3H]MRL-1 in LDC dogs, 20% of the administered radioactivity was recovered in pooled lymph over 168 hours, suggesting some level of recirculation of radioactivity related to [3H]MRL-1 from the plasma compartment into the lymphatic system. Experiments conducted in the LDC rat model also resulted in similar conclusions. Analysis of injection site s.c. tissue showed significant metabolism of [3H]MRL-1, which provides an explanation for the <100% bioavailability of therapeutic proteins and peptides after s.c. dosing. After s.c. dosing, the major circulating components in plasma were the parent peptide and the PEG-linker [3H]MRL-2. The metabolism profiles in lymph were similar to those in plasma, suggesting that the loss of peptide was minimal during lymphatic transport. After i.v. dosing in rats, [3H]MRL-1 was metabolized and excreted primarily in the urine as metabolites.
Introduction
Subcutaneous (s.c.) administration is the patient-friendly route of delivery for biologic drugs such as peptides and proteins. Due to poor membrane permeability and high enzymatic degradation in the gastrointestinal tract, their bioavailability after administration via nonparenteral routes is low and variable (Tang et al., 2004; Lin, 2009). Even though s.c. delivery has been used for biologic drugs for many years, little is known about the processes that govern the absorption of macromolecules from the subcutaneous space and the impact on bioavailability and pharmacokinetic profiles in various species. Based on studies conducted in thoracic lymph-duct cannulated (LDC) sheep model, it has been suggested that the lymphatic system plays an important role in the absorption of large molecules after s.c. administration.
It has been demonstrated that the proteins larger than 16–20 kDa are taken up primarily by the lymphatic system and transported to the systemic circulation via the thoracic duct whereas smaller molecules primarily diffuse directly into blood capillaries (Supersaxo et al., 1990; Porter and Charman, 2000; Tang et al., 2004). The walls of blood capillaries are composed of a sealed endothelium with tight junctions that only allow small molecules but limit large molecules to diffuse. On the other hand, the lymphatic capillary has a more open structure, with an incomplete basal lamina and an absence of tight junctions, which allows free passage for large molecules. Once they are administered through s.c. injection, large proteins travel from the local extracellular space in the subcutis into the peripheral lymphatics and regional lymph nodes, collecting at the central lymphatics, and finally entering the vascular circulation (Swartz, 2001; McDonald et al., 2010). Studies conducted in the sheep model have also contributed to our understanding of lymphatic uptake after central versus peripheral lymph duct-cannulation, the impact of injection site on lymphatic uptake, and the potential for drug clearance during lymphatic transport (Supersaxo et al., 1988; Charman et al., 2000; Charman et al., 2001; McLennan et al., 2005, 2006; Kota et al., 2007).
However, it is not clear whether the molecular weight dependency for s.c. lymphatic absorption established in sheep is also applicable to other animals. In the process of drug discovery and development, preclinical models for pharmacokinetics and nonclinical safety studies usually consist of one rodent (usually the rat) and one large animal species (typically dogs or monkeys). Therefore, developing LDC models in relevant animal species such as the rat and dog is of great interest to better understand the absorption, biodistribution, and metabolism of biologic drugs after s.c. administration.
There have been conflicting reports with lymphatic absorption studies conducted in a rat model (Porter et al., 2001; Edwards et al., 2001; Kaminskas et al., 2009). Lymphatic absorption was a minor component in the s.c. absorption of bovine insulin (5.6 kDa), recombinant human erythropoietin alfa (30.4 kDa), and bovine albumin (66 kDa) in the rat (Kagan et al., 2007). In another study in rats, the thoracic lymph recovery of five proteins (molecular weight 7.5 to 75 kDa) was low after s.c. administration (Xie and Hale, 1996). The reason for the discrepancy between the results obtained in rat and those in sheep are not clearly understood.
In the present study, we used a 46 kDa pegylated peptide, a neuromedin-U receptor agonist (MRL-1), to investigate the following: 1) the mechanism of absorption after s.c. administration, 2) the potential for recirculation of systemic MRL-1 to the lymphatic system after i.v. administration, 3) drug loss at the injection site and during lymphatic transport, and 4) the mechanism of elimination. Thoracic LDC dog and rat models were developed that allowed continuous lymph collection for up to 8 days. The results obtained for lymphatic absorption, in vivo metabolism, and the elimination mechanism of [3H]MRL-1–related radioactivity are discussed.
Materials and Methods
Chemicals.
The PEGylated peptide MRL-1, [3H]MRL-1 and [3H]MRL-2 (Fig. 1) were prepared at Merck Research Laboratories (Rahway, NJ), as described previously elsewhere (Ingallinella et al., 2012). Briefly, the free (unconjugated) peptide was dissolved in 8 M urea, 4 mM EDTA, and 0.1 M sodium phosphate (pH 7.1) and was reacted with 40 kDa PEG-maleimide linker (1.1 eq., dissolved in water) for 1 hour at room temperature. MRL-1 was purified by sequential cation exchange and size-exclusion chromatography. The purity of MRL-1 was >99% as determined by high-performance liquid chromatography (HPLC). The specific activity of [3H]MRL-1 was 22 Ci/mmol, and the radiochemical purity was >99%, as determined by HPLC. The calculated isoelectric point was 8.7 for the native peptide. The 40 kDa amino-PEG intermediate was obtained from NOF Corporation (Tokyo, Japan). Acetonitrile and methanol (HPLC grade) were purchased from Thermo Fisher Scientific (Fair Lawn, NJ).

Chemical structures of [3H]MRL-1 and [3H]MRL-2.
Thoracic LDC Animal Models and Study Design.
All experiments were performed according to procedures approved by the Merck Institutional Animal Care and Use Committee. A parallel study design with thoracic LDC and sham-operated control dogs and rats was used in which a single dose of [3H]MRL-1 was administered to each animal either i.v. or s.c. In both animal models, the s.c. injection sites are located at a similar hind-leg region to minimize the potential variation in dose recovery from lymph due to the location of injection site (Tilney, 1971; Porter and Charman, 2000). The treatment groups, administered doses, and collected samples are listed in Table 1. [3H]MRL-1 was formulated as a solution in 5% mannitol buffer containing 6 mM sodium acetate (pH 5.0).
Treatment groups, administered doses and collected samples in rats and dogs
LDC Dog Model.
Adult male beagle dogs (∼5 kg) were surgically prepared ∼24 hours before dosing (Khoo et al., 2001; Shackleford et al., 2003). The dogs were fed with lipid meal before surgery to facilitate identification and isolation of the thoracic duct. The dogs were anesthetized and positioned in dorsal recumbency, with the forelimbs pulled caudally to expose the ventral neck and thoracic inlet region. An incision was made over the left external jugular vein, and the cephalic vein and the omocervical artery were ligated and divided. This facilitated exposure of the thoracic duct ampulla at its entrance into the jugular vein as well as the portion of the duct traveling into the thoracic inlet. Any branches entering the duct in this region were ligated.
The duct was cannulated close to the ampulla with a 0.03-inch inner diameter silastic catheter, which was tunneled under the skin to a small incision over the animal's shoulder region and exteriorized. Similarly, for the sham-operated control animals the same procedures were performed to separate the thoracic lymph duct without the actual cannulation.
After anesthetic recovery, the cannula was connected to a sterile fluid collection bag containing 1000 IU of heparin. Saline solution (200–400 ml) was given s.c. to facilitate lymph flow and to prevent dehydration. The frequency of the saline supplement was determined by the veterinary staff. At the conclusion of the study, the catheters were removed percutaneously with no adverse effects on the animal. The lymph flow rate in the dogs ranged between 8–40 ml/h, with an average flow of 23.1 ml/h (Edwards et al., 2001).
The LDC dogs and the corresponding control dogs were administered [3H]MRL-1 s.c. (3 mg/kg, 0.5 ml/kg) at the popliteal region of the dog's hind limb, on the caudal aspect of the knee, or as an i.v. bolus (1 mg/kg, 0.5 ml/kg) via a jugular vein cannula. The carotid artery of all animals was also cannulated for blood collection. Blood samples were collected before the dose, then at 5 (i.v. only), 15, and 30 minutes, then at 1, 2, 4, 8, 24, 48, 72, 96, and 120 hours in heparinized tubes, and were spun in a centrifuge at 1600g at 4°C for 10 minutes to obtain plasma. Lymph fluid samples were collected over the following periods: 0–1, 1–2, 2–4, 4–6, 6–8, 8–24, 24–26, 26–28, 28–30, 30–32, 32–48, 48–72, 72–96, and 96–120 hours. Urine and feces samples were collected at 0–4, 4–8, and 8–24 hours, then once a day during the study period. The weights of the lymph fluid and urine samples were also recorded. The plasma, lymph, and urine samples were stored at −80°C until analysis.
LDC Rat Model.
Male Sprague-Dawley rats (∼350 g) were surgically prepared ∼48 hours before dosing (Edwards et al., 2001; Ionac, 2003; Kagan et al., 2007). The rats were anesthetized in a 5% isoflurane induction chamber with oxygen flow rate at 1 l/min. The surgical areas were shaved and prepared for aseptic surgery with DuraPrep (3M Company, St. Paul, MN). After transferring it to a nose cone, the isoflurane concentration was reduced to 2%. Rats were given 0.5–1.0 ml of olive oil by gavage ∼0.5 hours before the operation for visualization of lymph vessels.
After setting up a sterile surgical field, a midline abdominal incision was made approximately two-thirds of the length of the abdomen to the xiphoid cartilage. The liver and the gut were mobilized (pulled upward and to the right, respectively), and were kept moist using warm, saline-soaked gauze. The thoracic duct and dorsal aorta were located using cotton-tipped applicators to gently pull apart the connective tissue. The dorsal aorta was separated from the psoas muscle, exposing the thoracic duct. The duct was then separated from the psoas muscle and dorsal aorta using fine forceps and cotton-tipped applicators. The duct was cannulated (3–5 mm) with a heparin (500 units/ml) and saline-filled catheter (described below).
Once the lymph flow had been established, the cannula was tied down by 5-0 silk ligatures. If a proper seal could not be achieved with ties, tissue adhesive would be placed over the ligatures. The cannula was then tunneled s.c. around the back and exteriorized at the back of the neck. A spring tether system was installed and attached to a swivel to allow free movement of the animal. The abdominal muscles were then closed by using 4-0 PDS-II sutures. The skin was closed using sterile wound clips or nonabsorbable sutures (3-0 silk). For the sham-operated control animals, the same procedures were performed to separate the thoracic lymph duct without the actual cannulation.
A modified catheter system was used that has two catheters combined with a mixing chamber (Wang et al., 2010). Lymph fluid flows from the cannula to the perforation in catheter one, then enters the mixing chamber, which encases the perforated portion of catheter one. Microinfusion of heparin (500 units/ml) was applied from the second catheter into the mixing chamber at a constant flow rate of 50 μl/h to prevent the lymph fluid coagulation and maintain a continuous flow.
After the surgical procedures, the rats were monitored during the recovery period until they regained full consciousness. Electrolyte replacement solution (PRANG; Bio-Serv, Frenchtown, NJ) along with water was supplied to the rats to encourage fluid intake and to facilitate overall lymph flow. Rats were allowed to recover and stabilize for approximately 48 hours before dosing. Lymph fluid was continuously collected during this period; the lymph flow rate in rats ranged between 0.8 and 3.4 ml/h, with an average flow of 2.2 ml/h (Edwards et al., 2001).
The LDC rats and the corresponding control rats were administered [3H]MRL-1 s.c. (3 mg/kg, 0.5 ml/kg) at the lateral lower part of the left hind limb, or as an i.v. bolus (1 mg/kg, 0.5 ml/kg) via a femoral vein cannula. The carotid artery of all animals was also cannulated for blood collection. Blood samples were collected before the dose, then at 5 (i.v. only), 15, and 30 minutes, then at 1, 2, 4, 8, 24, 48, 72, 96, 120, 144, and 168 hours in heparinized tubes. The blood samples were spun in a centrifuge at 1600g at 4°C for 10 minutes to obtain plasma. Lymph fluid samples were collected over the following periods: 0–1, 1–2, 2–4, 4–6, 6–8, 8–24, 24–26, 26–28, 28–30, 30–32, 32–48, 48–72, 72–96, 96–120, 120–144, and 144–168 hours. Urine and feces samples were collected at 0–4, 4–8, and 8–24 hours, then once a day for a week in preweighed plastic bottles. The plasma, lymph, urine, and feces samples were stored at −80°C until analysis.
In Vivo Metabolism in Plasma and at the Site of Injection in Dogs and Rats.
To determine the metabolism at the injection site and plasma, we dosed intact dogs and rats (n = 1/time point/species) with [3H]MRL-1 s.c. at 3 mg/kg. Plasma and injection site skin samples were obtained at the predefined time points of 1, 7, and 48 hours after dosing in dogs, and 1, 8, 48, and 432 hours after dosing in rats. Dogs scheduled for termination were used in this study. Skin samples at the injection site were cut in 3-inch (rat) or 5-inch (dog) diameters (7.62 cm or 12.7 cm, respectively) along with s.c. tissue. The plasma and skin samples were stored at −80°C until further analysis.
Sample Preparation for Metabolite Analysis in Plasma, Lymph, and Injection Site.
Plasma and lymph samples (2 ml) from dog and rat were mixed with equal volumes of acetonitrile. The mixture was vortex-mixed and spun in a centrifuge at 14,000g for 10 minutes. The extraction recovery of radioactivity was >90%. All operations involving skin samples were conducted at 4°C. Skin samples were thawed, cut into small slices, and transferred into a clean plastic bottle chilled on ice. Water (w/v; 1:3) was added to it and homogenized using Polytron PT3100 homogenizer (6 × 30 seconds). The skin homogenate was extracted with acetonitrile as described above for plasma. The supernatants of plasma and skin extracts were then concentrated under nitrogen and subjected to HPLC separation (see below). The peak fractions were collected, concentrated under nitrogen and treated with 0.5 M NaOH at ambient temperature for 1 hour to cleave the peptide from 40 kDa PEG. The reaction mixtures were then neutralized and the cleaved peptides were analyzed by liquid chromatography with mass spectrometry (LC-MS/MS).
Measurement of Radioactivity.
Aliquots of plasma, lymph fluid, skin homogenate, and urine were mixed with Ultima-Flo scintillation fluid (Packard BioScience, Meriden, CT) and the total radioactivity in these samples was determined by direct counting in a PerkinElmer Tri-Carb 3100 Liquid Scintillation Analyzer (PerkinElmer Life and Analytical Sciences, Waltham, MA). Aliquots of fecal homogenate (in water, 3:1; v/w) were dissolved in two volumes of Solvable (PerkinElmer) and bleached in two volumes of sodium hypochlorite before mixing with Hionic-Fluor (PerkinElmer) and direct counting as described previously.
Instrumentation.
A Shimadzu HPLC system (Shimadzu Corporation, Kyoto, Japan) equipped with two Series LC-10ADVP micropumps and Series SIL-10ADVP autosampler was used to obtain the metabolism profiles. Separation of metabolites was achieved on a Jupiter Proteo C12 column (4 µm particle size, 90 Å pore size, 4.6 × 250 mm) (Phenomenex, Torrance, CA). Mobile phases A and B consisted of water and acetonitrile, respectively, and contained 0.1% of formic acid. The column was eluted with a gradient of 40%–50% B in 40 minutes with a flow rate of 1.5 ml/min. The column was heated at 45°C to obtain better separation of MRL-1 and its metabolites. The column eluate was directed into a β-Ram radiometric detector (In/Us Systems, Brandon, FL) for online radioactivity profiling. When the sample total radioactivity was too low for online detection, the column eluate was collected on 96-well scintillation plates, dried under nitrogen, and then read by a 1450 LSC and Luminescence counter (MicroBeta TriLux; PerkinElmer).
For metabolite identification in dog/rat plasma and at the injection site, an Acquity BEH C18 column (2.1 × 100 mm, 1.7 μm particle size) (Waters Corporation, Milford, MA) was used at a flow rate of 0.15 ml/min and heated at 40°C. Mobile phases A and B consisted of water and acetonitrile, respectively, and contained 0.1% of formic acid. The gradient was 0–35% B from 5 to 55 minutes. The effluent from the LC column was introduced into the ion source of the LTQ-Orbitrap mass spectrometer operated in positive mode. The MS method consisted of a full scan mass analysis over the range m/z 250 to 2000 at a resolving power of 15,000 followed by three FT MS2 scans for the three most intense ions from scan one at a resolving power of 7500. The LC-MS/MS data were acquired with Xcalibur 2.0 (Thermo Fisher Scientific, Waltham, MA) and processed using SEQUEST search in Bioworks 3.3, and Protein Calculator (Thermo Electron Scientific, Waltham, MA). By adding the sequence of the peptide of interest into an existing database, a user-defined database was built and used to search against to identify any related peptide metabolites.
Results
Lymphatic Uptake in Dogs after Subcutaneous and Intravenous Administration of [3H]MRL-1.
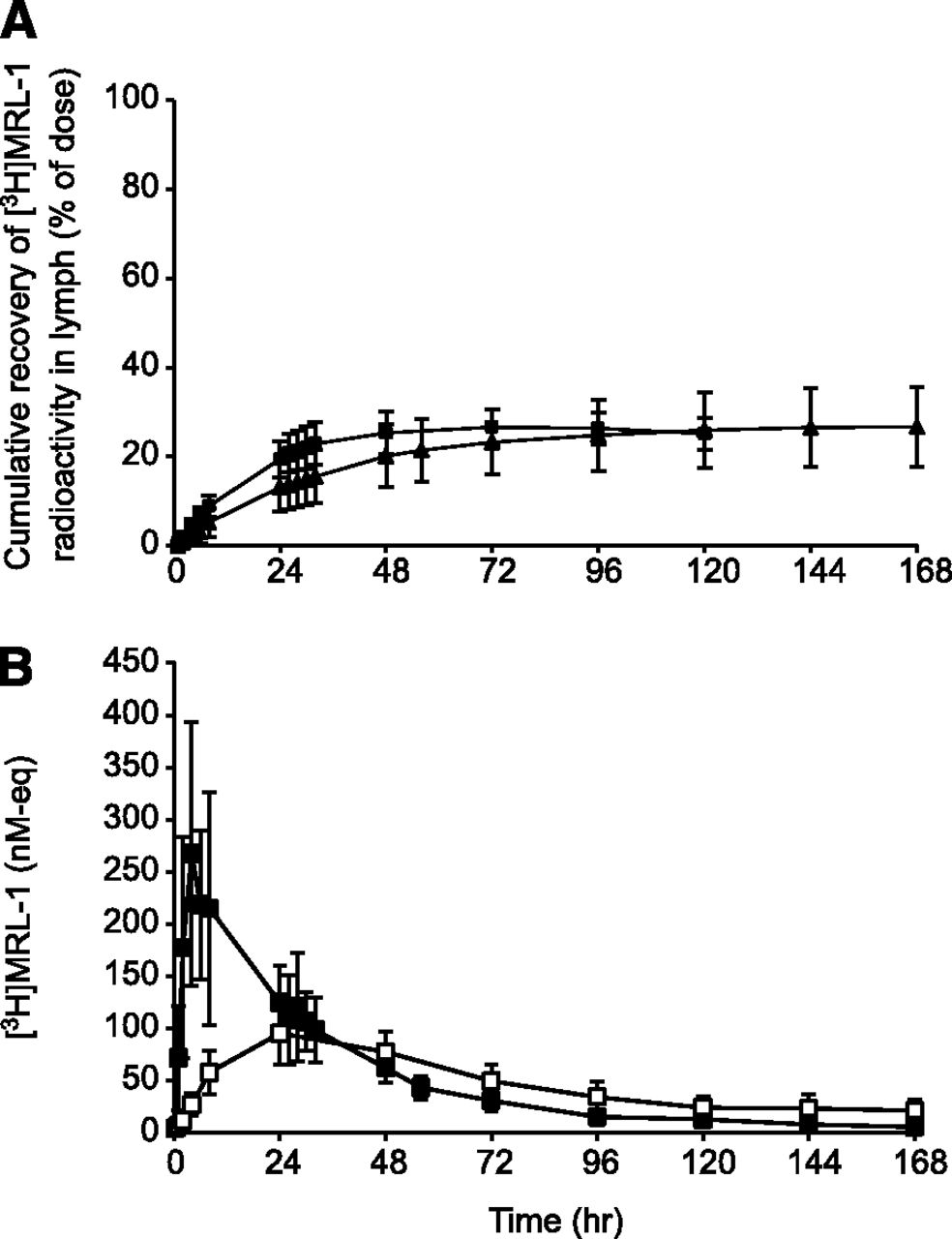
Figure 2 shows the cumulative recovery of radioactivity in lymph from LDC dogs after s.c. or i.v. dosing of [3H]MRL-1. Approximately 73% and 20% of the administered dose was recovered in lymph after s.c. and i.v. dosing, respectively (Fig. 2A and Table 2). Majority of the radioactivity was collected in lymph within 24–48 hours after s.c. and i.v. administration. Figure 2B shows the concentration of [3H]MRL-1 related radioactivity (nM equivalents) in lymph and plasma after s.c. dosing in LDC dogs. The radioactivity concentration in lymph during the absorption phase (within 24 hours) was much higher compared with that in plasma, suggesting that the s.c. administered radioactivity was absorbed directly into the lymphatic system. Figure 3 shows the mean plasma radioactivity concentration-time profiles after s.c. and i.v. dosing in LDC dogs, and the exposure (area under the concentration-time curve, AUC) data are shown in Table 3.

Cumulative (A) recovery and (B) concentration of [3H]MRL-1–related radioactivity in dog lymph. LDC dogs were dosed with [3H]MRL-1 at 3 mg/kg s.c. (▴) or 1 mg/kg i.v. (▪), and the radioactivity in the collected lymph (▪) and plasma (□) was measured using a liquid scintillation analyzer as described in the text.
Cumulative recovery of [3H]MRL-1 related radioactivity (percentage of dose) from lymph, urine, and feces in rats and dogs.
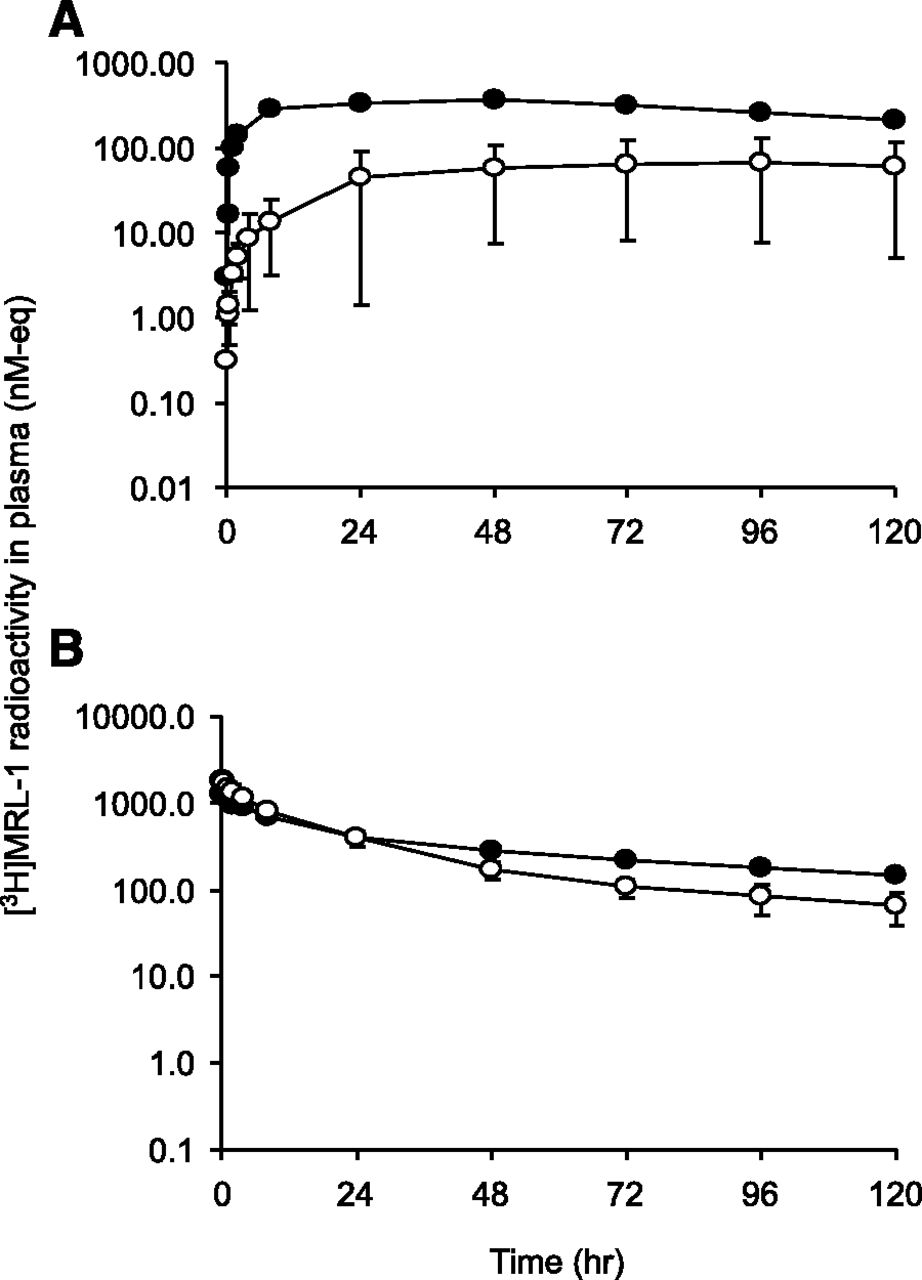
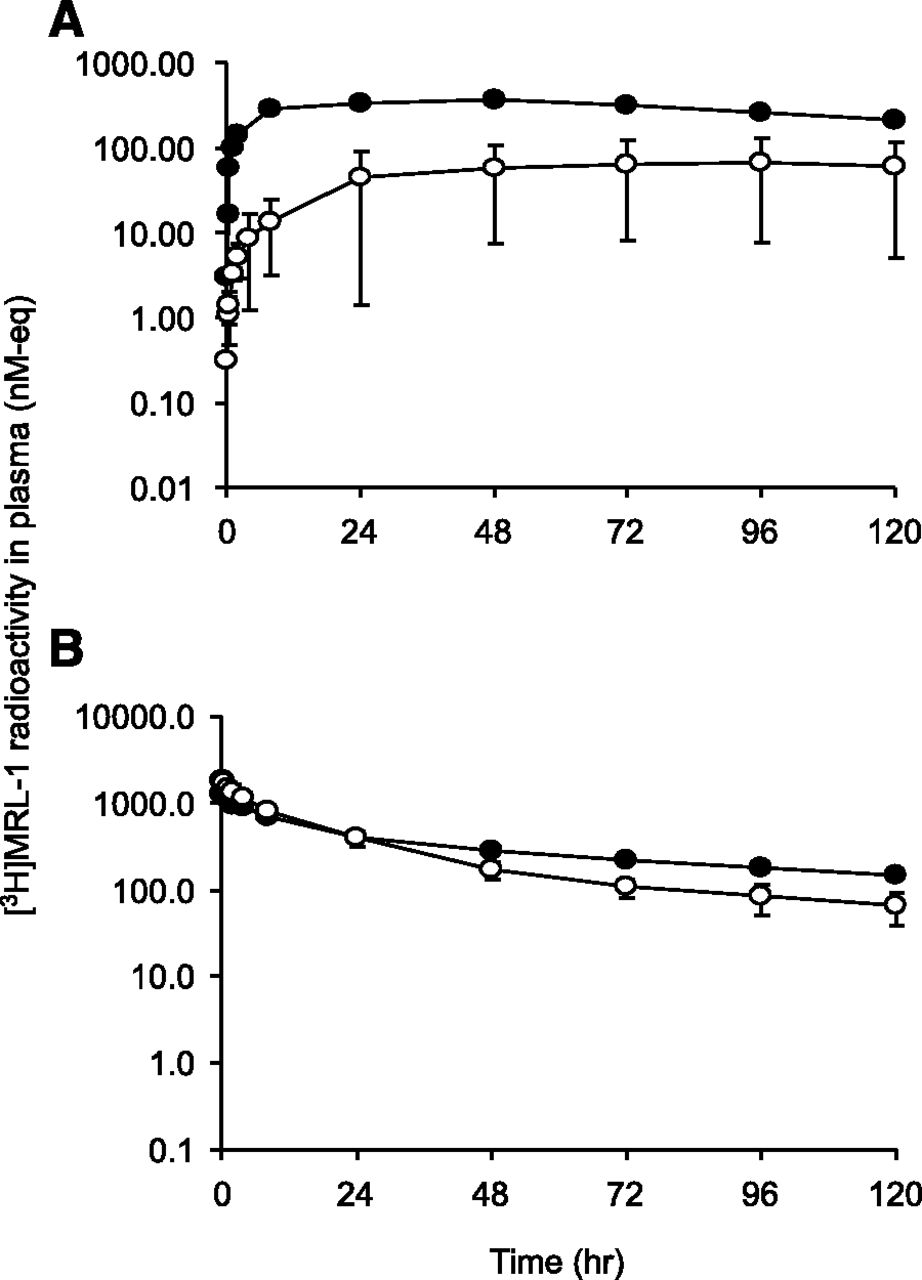
Mean plasma radioactivity concentration versus time profiles in LDC (○) and noncannulated control dogs (●) after administration of [3H]MRL-1 at (A) 3 mg/kg s.c. or (B) 1 mg/kg i.v. Radioactivity in plasma was measured using a liquid scintillation analyzer as described in the text.
Plasma exposure of [3H]MRL-1 radioactivity in LDC rats and dogs
Consistent with lymphatic absorption, the systemic exposure (AUC0–120 h) to [3H]MRL-1 related radioactivity was reduced by 82% when compared with that in sham-operated control dogs after s.c. dosing (Fig. 3A). After i.v. dosing, the AUC0–120 h to radioactivity was reduced by only 16% in LDC dogs compared with that in sham-operated control dogs (Fig. 3B).
Lymphatic Uptake in Rats after Subcutaneous and Intravenous Administration of [3H]MRL-1.
Figure 4 shows the cumulative recovery of radioactivity in lymph from LDC rats after s.c. or i.v. dosing of [3H]MRL-1. Approximately 27% and 26% of the administered radioactivity was recovered in lymph after s.c. and i.v. administration, respectively (Fig. 4A and Table 2). The majority of the radioactivity collected in lymph was recovered within 24–48 hours after s.c. and i.v. administration. Figure 4B shows the concentration of [3H]MRL-1 related radioactivity (nM equivalents) in lymph and plasma after s.c. dosing.

Cumulative (A) recovery and (B) concentration of [3H]MRL-1–related radioactivity in rat lymph. LDC rats were dosed with [3H]MRL-1 at 3 mg/kg s.c. (▴) or 1 mg/kg i.v. (▪), and the radioactivity in the collected lymph (▪) and plasma (□) was measured using a liquid scintillation analyzer as described in the text.
The peak concentration in lymph was only about 2-fold higher than that in plasma, suggesting that the lymphatic uptake in rats was not as significant as that observed in dogs after s.c. dosing. The plasma radioactivity concentration-time profiles after s.c. or i.v. dosing of [3H]MRL-1 in LDC rats are shown in Fig. 5, and the AUC data are summarized in Table 3.

Mean plasma radioactivity concentration versus time profiles in LDC rats (○) and noncannulated control rats (●) after administration of [3H]MRL-1 at (A) 3 mg/kg s.c. or (B) 1 mg/kg i.v. Radioactivity in plasma was measured using a liquid scintillation analyzer as described in the text.
After s.c. dosing, in agreement with the lymphatic recovery of radioactivity, the systemic exposure (AUC0–168 h) to [3H]MRL-1–related radioactivity was 44% of that observed in the sham-operated control rats (Fig. 5A). After i.v. dosing, the exposure to radioactivity was 70% of that observed in the control rats (Fig. 5B), suggesting some recirculation of systemic radioactivity into the lymphatic system.
Metabolism Profiles of [3H]MRL-1 in Plasma and at the Injection Site in Dog and Rat after Subcutaneous Dosing.
The metabolism profile of [3H]MRL-1 was evaluated in intact dogs and rats after s.c. dosing. Figure 6, A and B, depicts representative radiochromatograms of plasma at 48 hours from dogs and rats, respectively. The major circulating components in dog and rat plasma were the parent peptide [3H]MRL-1 and a metabolite M1 at 30 minutes. The retention time of M1 matched with that of an authentic sample of PEG-linker [3H]MRL-2 (Fig. 1). Also, base hydrolysis (see below under Identification of Metabolites) of the isolated peak fraction of M1 did not produce any detectable amino acids (data not shown). Based on these data, we concluded that structure of M1 is identical or similar to [3H]MRL-2. However, minor changes to the linker or PEG (for example, hydrolysis of maleimide) are difficult to confirm.
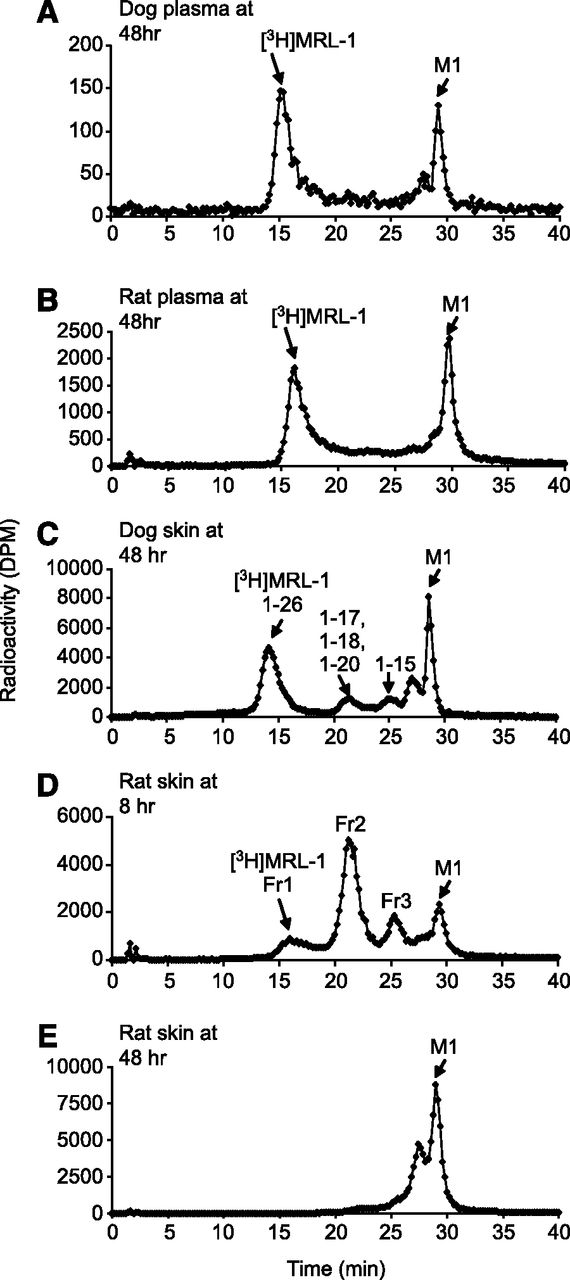
Representative HPLC radio chromatograms of plasma and injection site skin extracts from dogs and rats after s.c. administration of [3H]MRL-1. (A) Dog plasma at 48 hours. (B) Rat plasma at 48 hours. (C) Injection site in dog at 48 hours. (D) Injection site in rat at 8 hours. (E) Injection site in rat at 48 hours. Dogs and rats were dosed s.c. with [3H]MRL-1 at 3 mg/kg, and plasma and injection-site skin tissues (3–5 inch diameter) were extracted and analyzed as described in the text.
Figure 6, C–E, depicts representative radiochromatograms of extracts of the s.c. injection site in dogs at 48 hours and in rats at 8 and 48 hours after dosing, respectively. The metabolite profile in dogs at 48 hours was qualitatively similar to that in rats at 8 hours. As we will describe, there were total of eight metabolites detected at the injection site in dogs and rats. These metabolites, along with [3H]MRL-1 and [3H]MRL-2, were resolved into four broad peaks on HPLC analysis.
As shown in Fig. 6, C and D, although metabolite profiles in dog and rat injection sites were qualitatively similar, they were significantly different quantitatively. Metabolism of [3H]MRL-1 at the injection site in rats was much faster compared with that in dogs; only a small amount of intact [3H]MRL-1 remained at 8 hours, and mostly PEG-linker [3H]MRL-2 remained at 48 hours. Analysis of the skin at the s.c. injection site in rats showed that 62% and 17% of the administered radioactivity remained at 24 and 432 hours after dosing, respectively. Similar studies in dogs showed that 15% of the s.c. administered dose remained at the injection site 24 hours after dosing. In agreement with the extensive metabolism that occurred at the injection site in dogs and rats, the absolute bioavailability of MRL-1 was found to be low (<30%) in both species after s.c. dosing (data not shown).
Identification of Metabolites.
Due to the high mass and polydispersity of 40 kDa PEG, it is difficult to determine the accurate mass of the PEGylated derivatives (Webster et al., 2007). Therefore, base catalyzed cleavage of peptide from PEG followed by identification of the cleaved peptide by LC-MS/MS provided a convenient method for the structural determination of metabolites of [3H]MRL-1. The tritium label on the linker was stable, and the tritium loss was not observed in any reaction.
As described in Fig. 7, retro-Michael reaction of MRL-1 produced a dehydroalanine derivative of the full-length peptide 1-26, suggesting that the truncated metabolites of MRL-1 can also be analyzed using this procedure. The peak fractions (fractions 1–4; Fig. 6, C and D) purified using HPLC were treated with base, and the resulting peptides were analyzed by LC-MS/MS as described in Materials and Methods. Molecular weights of the metabolites detected under each peak are summarized in Table 4, and the LC-MS profiles are shown in Fig. 8.
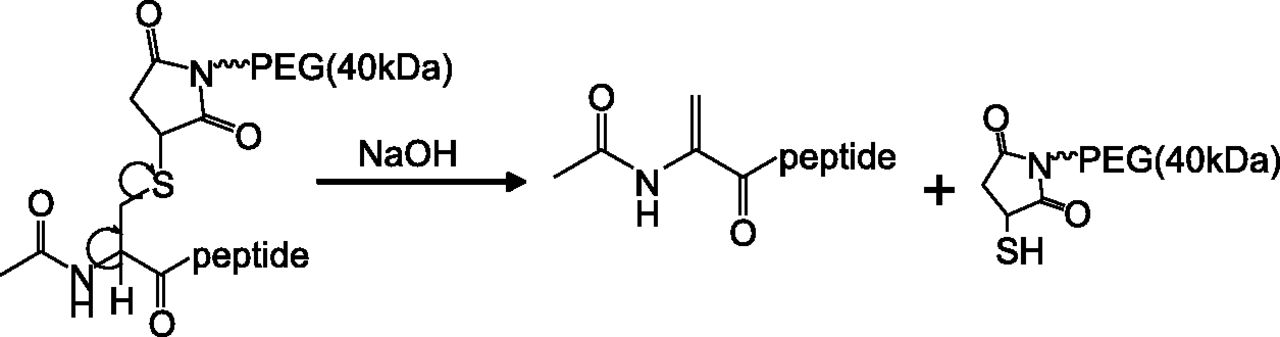
Retro-Michael reaction of MRL-1 resulting in the formation of dehydroalanine peptide derivative.
Accurate masses of truncated (pegylated) peptide metabolites of MRL-1 identified in vivo
Truncated peptide was cleaved from PEG in each metabolite using a chemical procedure and analyzed by LC-MS.

Extracted ion chromatograms of metabolites formed at the injection site in rats at 8 hours after s.c. dosing of [3H]MRL-1. Fractions 1–3 shown in Fig. 6D were purified, and treated with NaOH to cleave PEG. The cleaved peptides were then extracted and analyzed by LC-MS/MS as described in the text.
Fraction 1 (Fig. 8A) primarily consisted of intact PEGylated peptide MRL-1, as the HRMS (High Resolution Mass Spectrometry) data agreed well with full length 1-26 peptide. Metabolite with 1-25 sequence was also detected in trace amounts. Figure 8B shows that fraction 2 was a mixture of three metabolites, peptides 1-17, 1-18, and 1-20. As shown in Fig. 8C, there were two major metabolites under fraction 3: 1-12, and 1-15. In addition, 1-13 and 1-16 were identified in trace amounts under fraction 3. The masses of the metabolites determined by LC-MS matched well with the corresponding theoretical mass for each sequence (Table 4). A summary of the metabolic cleavage sites of the peptide MRL-1 is shown in Fig. 9.

Metabolic sites of [3H]MRL-1.
Excretion of [3H]MRL-1 in Dogs and Rats after Subcutaneous and Intravenous Dosing.
The amount of [3H]MRL-1–related radioactivity collected in the lymph and excreted in the urine and feces is summarized in Table 2. In both species, the majority of the excreted radioactivity was recovered in the urine. After i.v. dosing in LDC dogs, 48% and 20% of the administered radioactivity was recovered in 120 hours in urine and lymph, respectively. After s.c. dosing in LDC dogs, a total of 80% of the radioactivity was recovered in 120 hours: 73% in the lymph and 7% in the urine. The radioactivity in the dog feces was not determined.
After i.v. dosing in rats, 57% and 8% of the administered radioactivity was recovered in 144 hours in urine and feces, respectively. However, after s.c. dosing, only 30% and 9% of the administered radioactivity was recovered in 144 hours in urine and feces, respectively. Figure 10A depicts the representative radiochromatogram of the rat urine extract, and Fig. 10B shows the radiochromatogram of authentic standards of [3H]MRL-1 and [3H]MRL-2. Figure 10A shows that [3H]MRL-1 was not excreted intact and there were primarily two peaks, M1 and M2. On the basis of comparison of M1 retention time with that of an authentic standard (Fig. 10B), the structure of M1 was tentatively identified as MRL-2 (Fig. 1). The structure of M2, which is likely to be a truncated PEGylated peptide, was not further explored.

(A) Representative radiochromatogram of extract of urine from rats after i.v. dosing of [3H]MRL-1. Rats were dosed i.v. with [3H]MRL-1 at 3 mg/kg, and urine was collected up to 168 hours. Urine was extracted and analyzed by HPLC with fraction collection as described in the text. (B) Radiochromatogram of authentic standards of [3H]MRL-1 and [3H]MRL-2.
Discussion
We used a PEGylated peptide [3H]MRL-1 to determine the lymphatic uptake in LDC dogs and rats. MRL-1 is a 26 amino-acid peptide covalently attached to 40 kDa PEG using a maleimide linker. Linker region was selected for tritium labeling because in the event the peptide portion is truncated or completely cleaved at the site of injection, the label remains on the PEG linker (molecular weight ∼42 kDa) whose uptake can still be monitored by observing the radioactivity.
Results obtained in LDC dogs showed that majority of the s.c. administered radioactivity was absorbed via the lymphatic system. Consistent with these data, the radioactivity concentration in the collected lymph was far greater than that detected in plasma; the plasma AUC decreased proportionately when compared with that in sham-operated control dogs. These results demonstrate that predominantly lymphatic absorption of a 46 kDa molecule in LDC dogs agrees well with what has been previously reported in LDC sheep and dogs (Supersaxo et al., 1990; Charman et al., 2000; McLennan et al., 2005; Kota et al., 2007; Wang et al., 2012).
The results obtained from LDC dogs after i.v. dosing suggest that large molecules can travel from systemic circulation to lymphatic system through interstitial space to a limited extent, as observed elsewhere with pegylated polylysine dendrimers (22–68 kDa) in a rat model (Kaminskas et al., 2009). These results also likely reflect a process during which protein and peptide drugs transport from blood circulation to the site of action in target tissues to exert their pharmacologic activity. Macromolecules in blood can be pushed out across vascular walls along with fluid because of pressure and concentration gradients, and then can distribute into the interstitium. Protein drug molecules can then pass through the extracellular matrix by diffusion to the cell surface target, or they may be reabsorbed back into the lymph circulation (Audus and Raub, 1993; Wiig et al., 2008).
Although [3H]MRL-1 was primarily absorbed by lymphatic system in dogs after s.c. dosing, the data obtained in rats suggest that there could be s.c. anatomic differences between these species. Only about 27% of the s.c. dose administered was collected in thoracic lymph in LDC rats. This apparent discrepancy led us to conduct further analyses of the dose remaining at the injection site in rats to estimate the extent of absorption. The results suggested that only about 50% of the s.c. dose was absorbed during the lymph collection period of 168 hours in rats. Therefore, 27% of the s.c. administered dose (or ∼54% of the absorbed dose) recovery in thoracic lymph agrees well with the corresponding reduction in AUC (∼56%) in LDC rats. These data suggest that lymphatic uptake and vascular absorption are equally feasible in the s.c. absorption of MRL-1 in rats.
Similar results were reported for polylysine dendrimers in LDC rats (Kaminskas et al., 2009). However, in contrast with these results, lymphatic uptake was a minor component in the absorption of human tumor necrosis factor (17 kDa) and human erythropoietin alfa (30.4 kDa) in LDC rats (Kojima et al., 1988; Kagan et al., 2007). Evidently, the experiments conducted in LDC rats by various research groups have produced contradictory results on the role of lymphatic system in the s.c. absorption of large molecules. Additional studies are required to understand the anatomic differences between the lymphatic system of rodents and that of higher species, and to determine whether 16 kDa molecular mass cut off, suggested based on the sheep model, is also applicable in rodents for predominant lymphatic uptake.
Subcutaneous bioavailability of biologic drugs varies significantly (Tang et al., 2004; Richter et al., 2012). Metabolism in the subcutaneous space and during lymphatic transport have been suggested to be among the reasons for <100% bioavailability. The results obtained in our study provide direct evidence for the loss of drug in the subcutaneous space in dogs and rats. It is interesting to note that [3H]MRL-1 was metabolized at a significantly higher rate at the injection site in rats compared with dogs. On the basis of the amino acids present at the cleavage sites, we can speculate that trypsin, chymotrypsin, and other endopeptidases may be responsible for metabolism.
Currently, there is a lack of reliable information on the expression, distribution, and substrate specificity of these enzymes in the s.c. space or on the species differences in any of these attributes (Woodley, 1994; Olsen et al., 2004). The absorption and metabolism data we have described seem to suggest that [3H]MRL-1 is absorbed at a faster rate (in dogs) than the metabolites (in rats), which mainly constituted the PEG-linker with or without truncated peptide. It is possible that absorption of PEG is influenced by physicochemical properties (such as charge, size, etc.) of the attached protein (Richter et al., 2012).
Our data also suggest that the extent of metabolism at the injection site may influence the overall absorption rate and route (lymphatic versus vascular uptake) of large molecules. A limitation of our work is the inability to assess the bioavailability of MRL-1 based on total radioactivity due to metabolism and other factors. However, in agreement with these findings, a separate pharmacokinetics study indicated low absolute bioavailability of MRL-1 (<30%) in dogs and rats after s.c. dosing, and the bioavailability in rats tended to be even lower than in dogs (Zou et al., unpublished data).
Allometric scaling of preclinical pharmacokinetics is often used to estimate human pharmacokinetics and the dose of biologic drugs without consideration of their metabolism in the s.c. space. Potential species-specific metabolism in the s.c. space underscores the uncertainty in these extrapolations from preclinical data. Therefore, information obtained from in vitro metabolism studies in tissue preparations such as skin homogenate, hepatocytes, and kidney cell membranes from preclinical species and human may be used for cross-species comparison. Drug loss within the lymphatic system also has been reported. Based on the experiments conducted in peripheral and central duct-cannulated sheep, it was concluded that loss of human growth hormone within the lymphatics contributed significantly to its reduced bioavailability after s.c. administration (Charman et al., 2000). However, our study found that the metabolism profiles in lymph and plasma in comparison with those from the injection site suggested that MRL-1 clearance within the lymphatics is not significant.
PEGylation is one of the main approaches explored for improving the plasma half-life of biotherapeutics (Harris et al., 2001; Veronese and Pasut, 2005; Veronese and Mero, 2008). Although currently there are several marketed protein drugs that exploit the PEGylation strategy, only limited data are available on the mechanism of their elimination. Excretion of [3H]MRL-1–related radioactivity was slow in dogs and rats. However, it was evident in rats that elimination of [3H]MRL-1 was mainly through metabolism, followed by excretion of metabolites in the urine. It can be predicted based on the metabolites we have described that the peptide portion of MRL-1 was the main target for the hydrolytic enzymes, ultimately leading to the excretion of 40 kDa PEG. It is difficult to determine whether PEG itself was a target for metabolizing enzymes or whether any minor changes occurred on the PEG scaffold before excretion (Herold et al., 1989; Veronese and Pasut, 2005).
It is well recognized that glomerulus in the kidney filters substances mainly based on their molecular size, shape, and charge. Large molecules with molecular mass >70 kDa or molecular sizes >40 Å are mostly restricted from passing through glomerulus and are eliminated by other pathways, such as liver uptake, proteolytic digestion, and the immune system (Brenner et al., 1978; Takakura et al., 1987; Yamaoka et al., 1994; Hamidi et al., 2008). Because peptide in MRL-1 contains only 26 amino acids, 40 kDa PEG was thought to play a dominant role in the distribution and elimination of the conjugate. Because of extensive hydration, the hydrodynamic volume of 40 kDa is expected to be much higher than that of an equal size globular protein. Nevertheless, high polymer flexibility and deformity, together with its rod-like conformation allow for glomerular filtration, albeit at low rate. After glomerular filtration, MRL-1 is likely degraded through hydrolysis by brush border enzymes located on the luminal membrane or by endocytosis and subsequent hydrolysis.
Excretion details for few PEGylated biologics have been reported. Limited data from SDS-PAGE analysis have indicated that only 40 kDa material is present in rat urine after 400 mg/kg s.c. dosing of certolizumab, a 40 kDa PEGylated TNF-α antibody. Similarly, after i.v. dosing of [14C]IFNα-2a (Pegasys) in rats, the majority of excreted radioactivity was detected in the urine (51% of the dose), which mostly contained degraded interferon fragments and 40 kDa PEG (Modi et al., 2000). In contrast, PEGylated peptide-based erythropoiesis-stimulating agent peginesatide (45 kDa) was excreted mostly intact in the urine in rats and monkeys (Woodburn et al., 2012, 2013).
In summary, our study describes the significance of lymphatic system in s.c. absorption of a 40 kDa PEGylated peptide MRL-1 in dog and rat. Our data show the loss of the drug at the injection site and identify potential species differences in enzymatic activity in the s.c. space. The MRL-1 peptide was metabolized while still attached to PEG and was excreted in the urine as metabolites.
Authorship Contributions
Participated in research design: Reddy, Zou, Bateman, Adreani, Chris Johnson, Cunningham, Shen.
Conducted experiments: Zou, Cunningham, Trinh, Shen, Wang, Hong, Christine, Nunes.
Contributed new reagents or analytic tools: Reddy, Zou, Bateman, Zhang, Braun, Staskiewicz.
Performed data analysis: Reddy, Zou, Bateman, Kumar.
Wrote or contributed to the writing of the manuscript: Reddy, Zou, Bateman, Kumar, Adreani, Shen.
Footnotes
Abbreviations
- AUC
- area under the concentration-time curve
- EDTA
- ethylenediaminetetraacetic acid
- HPLC
- high-performance liquid chromatography
- LC
- liquid chromatography
- LC-MS/MS
- liquid chromatography coupled to tandem mass spectrometry
- LDC
- lymph duct-cannulation
- LSC
- liquid scintillation counting
- MS
- mass spectrometry
- PEG
- poly(ethylene glycol)
- Copyright © 2013 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}