


Abstract
Pyrrolobenzodiazepine (PBD)-dimer is a DNA minor groove alkylator, and its CD22 THIOMAB antibody drug conjugate (ADC) demonstrated, through a disulfide linker, an efficacy in tumor reduction for more than 7 weeks with minimal body weight loss in xenograft mice after a single 0.5–1 mg/kg i.v. dose. The DNA alkylation was investigated here in tumors and healthy organs of mice to understand the sustained efficacy and tolerability. The experimental procedures included the collection of tumors and organ tissues of xenograft mice treated with the ADC followed by DNA isolation/hydrolysis/quantitation and payload recovery from reversible DNA alkylation. PBD-dimer formed a considerable amount of adducts with tissue DNA, representing approximately 98% (at 24 hours), and 99% (at 96 hours) of the total PBD-dimer in tumors, and 78–89% in liver and lung tissues, suggesting highly efficient covalent binding of the released PBD-dimer to tissue DNA. The amount of PBD-DNA adducts in tumor tissues was approximately 24-fold (at 24 hours) and 70-fold (at 96 hours) greater than the corresponding amount of adducts in liver and lung tissues. In addition, the DNA alkylation levels increased 3-fold to 4-fold from 24 to 96 hours in tumors [41/106 base pairs (bp) at 96 hours] but remained at the same level (1/106 bp) in livers and lungs. These results support the typical target-mediated cumulative uptake of ADC into tumors and payload release that offers an explanation for its sustained antitumor efficacy. In addition, the low level of DNA alkylation in normal tissues is consistent with the tolerability observed in mice.
Introduction
Pyrrolobenzodiazepine (PBD)-dimers are DNA alkylators that exhibit extremely potent cytotoxic activities against multiple cancer cell lines (Gregson et al., 2001). These molecules function by covalently binding to the 2-amino groups of guanine in the minor grooves of cellular DNA, thereby forming DNA adducts that disrupt normal cell functions (Jenkins et al., 1994; Thurston and Bose, 1994; Gregson et al., 2001). We previously reported that an antibody drug conjugate (ADC) that combines a PBD-dimer payload, an anti-CD22 THIOMAB antibody, and a cyclobutyl-containing disulfide linker (1) with a drug-to-antibody ratio of 2 showed tumor reduction against CD22-expressing xenograft tumor models compared with the corresponding inactive ADC that utilizes a cyclopropyl-containing disulfide linker (2) in such assessments (Zhang et al., 2016a). In these experiments, a much lower level of the free PBD-dimer 3 released from 1 was observed in tumors relative to the amount of the free cyclopropyl thiol 4, a nonimmolation catabolite, produced via intracellular catabolism of 2 (severalfold; Fig. 1, A and B; Table 1) (Zhang et al., 2016a). We suspected that the measured quantity of the released PBD-dimer 3 in tumors might be underestimated given the known ability of PBD-dimer 3 to covalently bind to DNA and became “trapped” in a form that was not easily quantifiable (Zhang et al., 2016a). In contrast, cyclopropyl thiol 4 did not meaningfully bind to DNA, and the higher intratumor levels noted for this analyte relative to PBD-dimer 3 presumably reflect the difference in their DNA binding (Zhang et al., 2016b). In this study, the quantity of PBD-dimer in tumors and normal tissues was determined via DNA isolation and then the release of PBD-dimer from the PBD-DNA adducts. We envisioned that the use of this new assay would provide a less biased evaluation of the intracellular PBD-dimer distribution differences between tumors and normal organs after ADC administration. The amount of PBD-DNA adducts inside tumor cells should provide a better indicator of efficacy than the amount of free PBD-dimer (biomarker). Moreover, the characterization of the covalently bound drug may provide a better correlation of DNA alkylator concentrations with pharmacological/toxicological effects to support ADC preclinical studies.

(A), (B) The catabolism and payload release of anti-CD22 cyclobutyl- and cyclopropyl-containing disulfide PBD-dimer ADCs (1 and 2) resulting in PBD-dimer 3 that covalently binds to DNA but cyclopropyl thiol 4 that does not bind to DNA. (C) DNA digestion by nuclease P1 and release of PBD-dimer 3 from PBD-DNA adducts.
The concentration of free PBD-dimer 3 cyclopopyl-thiol 4 in tumor and major organs of WSU-DLCL2 xenograft mice after a single 5 mg/kg i.v. dose of anti-CD22 cyclobutyl–containing disulfide PBD-dimer and anti-CD22 cyclopropyl–containing disulfide PBD-dimer ADCs (1 and 2) (n = 2)
Results
DNA was isolated from the homogenates of tumor and major organ tissues of xenograft mice treated with ADCs 1 and 2. Digestion by nuclease P1 converted the isolated DNA to single nucleotides, and thus abrogated the interaction and binding affinity between PBD-dimer 3 and the minor groove of DNA (Fig. 1C) (Shimelis and Giese, 2006; Rahman et al., 2011). The covalent bonds between PBD-dimer 3 and the 2-amino groups of guanine were subsequently cleaved by heating of the sample after nuclease digestion (Rahman et al., 2011). Under these sample-processing conditions, PBD-dimer 3 was stable. PBD-dimer 3 was consequently recovered quantitatively from DNA adducts and then accurately quantitated by LC-MS/MS. As shown in Table 2, very high levels of PBD-dimer 3 were recovered from isolated tumor DNA after ADC 1 dosing to xenograft mice. At 24- and 96-hour time points, these values respectively represent 98% and 99% of total PBD-dimer in tumor tissues [total = free 3 (Table 1) plus DNA-bound 3 (Table 2)]. In other words, the amount of DNA-bound 3 in tumors was approximately 40-fold higher than the corresponding free levels at 24 hours, and about 120-fold higher when measured at 96 hours. Somewhat reduced bound/free ratios of 3 (3-fold to 10-fold) were observed in liver and lung after treatment with ADC 1, but the majority of PBD-dimer quantified in these tissues was still detected in the DNA-bound form (representing 78–89%) (Tables 1 and 2). Collectively, these data indicate that most of the PBD-dimer 3 payload released from ADC molecules in tumors and other tissues is sequestered via covalent binding to DNA, which should be considered as a disposition mechanism of PBD-dimer 3. PBD-dimer is an excellent agent that rapidly alkylates DNA in vitro (Thurston and Bose, 1994; Zhang et al., 2016b); its predominant distribution in DNA (forming covalent adducts) indicated that the same reaction occurred efficiently in vivo.
The DNA isolation and PBD-dimer recovery in tumor and major organs of WSU-DLCL2 xenograft mice after a single 5 mg/kg i.v. dose of anti-CD22 cyclobutyl-containing disulfide PBD-dimer and anti-CD22 cyclopropyl-containing disulfide PBD-dimer ADCs (1 and 2) (n = 2)
In contrast to the corresponding intratumor levels of free 3, the amount of DNA-bound 3 detected in tumors after the administration of ADC 1 increased with time. Specifically, the measured DNA-bound levels of PBD-dimer 3 were 3-fold to 4-fold higher at 96 hours relative to the level detected at 24 hours (192–277 versus 64–84 pmol/g tumor tissue) (Table 2; also compare with free levels in Table 1). The data suggest that there is an accumulation of PBD-dimer in tumor DNA during this period of time, and that further accumulation of PBD-dimer in tumors after 96 hours can be expected, with an assumption that the tumor antigen is not downregulated upon ADC treatment. Interestingly, a related, time-dependent increase in the quantity of DNA-bound 3 was not observed in liver or lung tissues, and the absolute values of detected 3 (free and DNA-bound) at both 24 and 96 hours were much lower than those noted in tumors (Table 2). We speculate that a combination of factors may limit PBD-dimer accumulation in normal healthy tissues during these experiments including: (1) a reduced uptake of ADC 1 into non–antigen-containing organs; (2) preferential disposition of the released payload drug 3 in healthy organs, such as liver and kidneys, that may contain additional drug metabolism enzymes and transporters for drug clearance from tumors; and (3) possible active DNA repair in the healthy organs (Drabløs et al., 2004; Fu et al., 2012; Kraynov et al., 2016). For unknown reasons, the amounts of DNA-bound PBD-dimer 3 observed in kidney tissues was significantly lower (below the lower limit of quantitation) than those observed in the liver and lung.
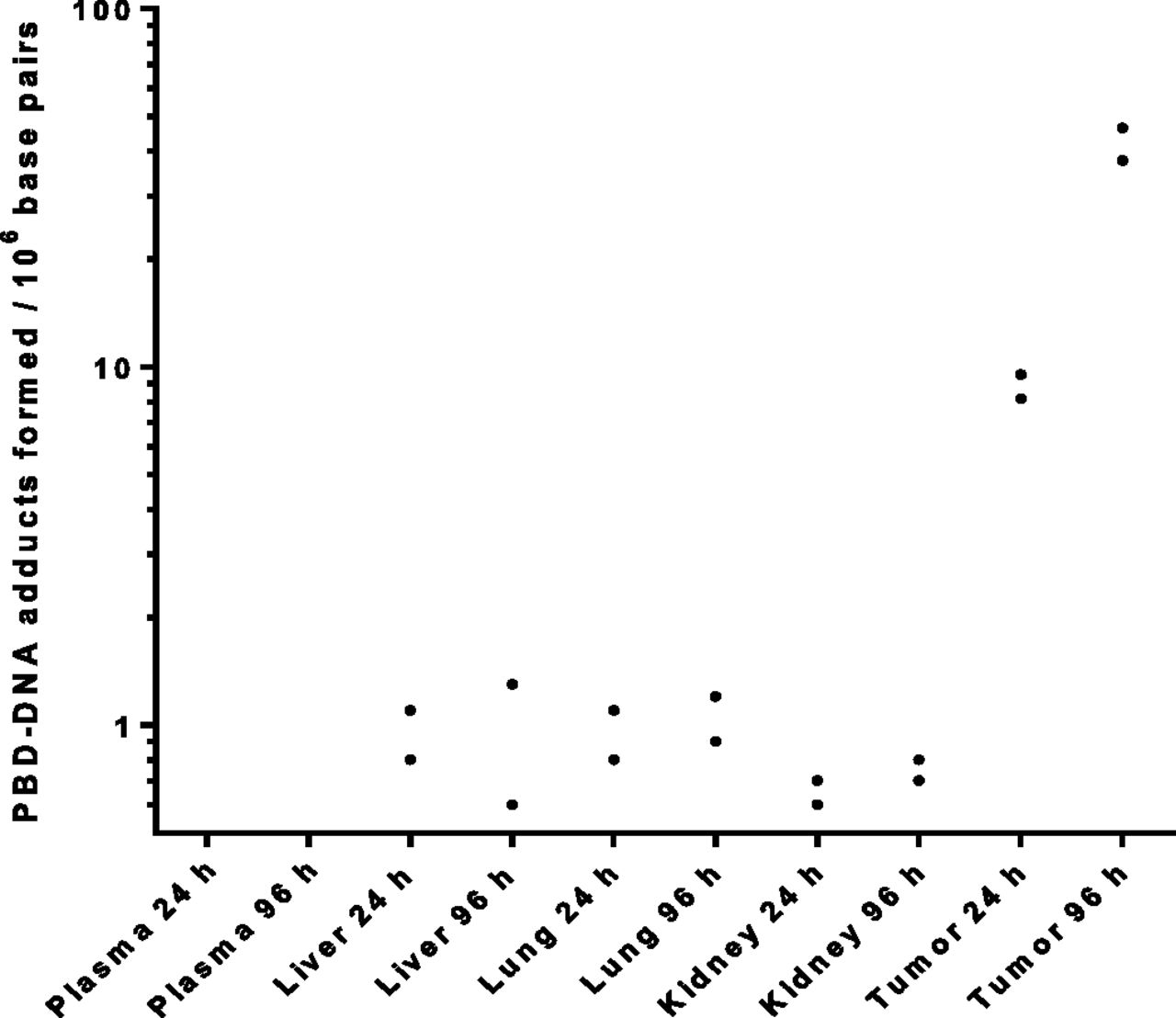
As shown in Table 2, a greater amount of DNA was isolated from tumors relative to the normal tissues that were analyzed (on the basis of milligrams per gram of tissue). These differences may be due to genomic duplications and amplifications in the cancer cells that are not present in the normal cells (Habano et al., 2007; Mwakigonja et al., 2010). The yield of isolated DNA also likely depends on how well the tissues were homogenized and the related experimental operations that may be difficult to perfectly control (Aldous et al., 2005; Triant and Whitehead, 2009). To better account for the observed variation in isolated DNA values, we calculate the number of PBD-dimer adducts formed per unit length of DNA using recovered PBD-dimer 3 and digested nucleotides from the same isolated DNA samples (Fig. 2; Table 2). Using this method, the amount of PBD-dimer 3 adducts bound to tumor DNA (PBD-DNA adducts) was calculated to be approximately 10 per million bp at 24 hours after a single dose of ADC 1 administered to xenograft-bearing mice (Fig. 2). The value increased to about 41 million PBD-DNA adducts (Fig. 2). In contrast, the corresponding amounts of PBD-DNA adducts calculated for lung and liver tissues were approximately 1 PBD-dimer 3 per million bp at both 24 and 96 hours (Fig. 2). Related values for kidney tissues were even lower. Therefore, the PBD-dimer level per unit weight of DNA (PBD-DNA adducts) may be a more appropriate indicator of DNA damage caused by PBD-dimer binding. The tolerability of this ADC in mice was thus corroborated, although the quantitative correlation is yet to be established between the adduct number and the triggering of cell apoptosis events (Pepper et al., 2004).

The number of PBD-DNA adducts formed per million bp in DNA of tumor and major organs at 24 or 96 hours after a single 5 mg/kg i.v. dose of anti-CD22 cyclobutyl- and cyclopropyl-containing disulfide PBD-dimer ADCs (1 and 2) (n = 2). No PBD-DNA adducts were detected in plasma samples.
Interestingly, a low level of DNA-bound PBD-dimer 3 was detected in tumors obtained from xenograft mice dosed with the cyclopropyl-containing ADC 2 despite poor release of the active payload 3 from this ADC (Zhang et al., 2016a,b). It is unclear whether this observed level of DNA-bound 3 resulted from a simple chemical degradation of ADC 2 (Fig. 1B) or from any enzymatic cleavage that was involved in the linker cleavage. In any event, the low levels of the PBD-DNA adducts observed in tumors after treatment of ADC 2 were significantly lower than those from ADC 1. These intratumor values of PBD-DNA adducts also correlated well with their distinct efficacy profiles for ADC 1 and 2. The DNA isolation and PBD-dimer recovery analysis unambiguously confirmed that this ADC had no efficacy in xenograft mice due to minimal DNA alkylation by the active payload that was released in tumor cells.
Discussion
The accurate quantitation of isolated DNA and recovered PBD-dimer facilitated the direct comparison of DNA alkylation levels in tumor and organ samples. The marked difference in formation of PBD-DNA adducts between the tumor and organs could support a wide therapeutic window by the ADC in mice. Slower trafficking of the ADC molecules into cells of organ tissues via pinocytosis (Jedema et al., 2004; Kovtun and Goldmacher, 2007) compared with the antigen-antibody recognition–mediated ADC uptake in tumor cells likely contributes to the observed differences in tissue distribution.
Achieving strong in vivo efficacy results with ADCs requires a given conjugate to deliver enough cytotoxic payloads into tumor cells in a reasonably short period of time compared with the long half-life of ADCs in circulation. In the previous study (Zhang et al., 2016a), we have demonstrated that the chemical structure and concentrations of intratumor catabolites ultimately determine the efficacy of an ADC, and that systemic exposures could not predict ADC efficacy. In the current study, the detailed quantitation of PBD-DNA adducts revealed that most of PBD-dimer payload 3 released intracellularly in tumors and organs by ADC 1 covalently bound to DNA. In addition, the determination of the PBD-dimer level per unit length of DNA allowed the direct comparison of DNA alkylation levels between tumor and various organ samples. Assuming 3 billion bp for genomic DNA, the DNA adduct level of 41 PBD-dimers/106 bp determined at 96 hours postdose suggests that approximately 12,000 PBD-dimers per cell (41*3 × 109/106/10, considering dose difference) are required to sustain the observed efficacy of 7-week tumor reduction after a 0.5–1 mg/kg dose of this ADC (Zhang et al., 2016a). Therefore, an equivalent to 42 ng of PBD-dimer at 96 hours after ADC administration would need to be delivered to a 0.3-g tumor (the actual tumor size used in the xenograft models) to support the observed efficacy. The 42 ng PBD-dimer represented 21% and 42% of the 1 and 0.5 mg/kg ADC 1 doses, respectively, for a 25-g mouse. Based on the total antibody concentration and free PBD-dimer concentration that were determined previously (Zhang et al., 2016a), and on the DNA-bound PBD-dimer level that is reported in this study, it was estimated that approximately 80% of ADC molecules that entered tumors were covalently bound to DNA at 96 hours postdose.
Once released at the site of action after ADC administration, a DNA alkylator like PBD-dimer 3 is “trapped” through covalent binding to DNA. The DNA adduct accumulation in tumors and tissues, once it reaches certain thresholds, may trigger cell apoptosis events and subsequent tumor killing. Therefore, a DNA alkylkator ADC, such as the one described in this study, could show an extended efficacy period even after the ADC was cleared from systemic circulation. In contrast, MMAE (maleimide-monomethyl auristatin E) and DM1, the reversible inhibitors that can diffuse away from the site of action (“diffusible” payloads), which may require the presence of certain levels of their ADCs to be in circulation for constant payload supply. It is not surprising that MMAE and DM1 ADCs need to be dosed every 3 weeks to support their long-term efficacy (prescribing information for KADCYLA, ado-trastuzumab emtansine, Genentech, 2013; and Adcetris, Brentuximab Vedotin, Seattle Genetics, 2011). Therefore, the sustained period of DNA alkylation that correlated with the efficacy may support less frequent dosing of a DNA alkylator ADC than the normal every 3 weeks. This less frequent dosing should allow for longer time between dose cycles to facilitate normal tissue repair through DNA repair mechanisms (McHugh et al., 2001; Clingen et al., 2005) and the disposition of payload drugs. Consistent with our findings and logic, an analog PBD-ADC is currently dosed every 6 weeks in a clinical trial (Pietanza et al., 2015).
In conclusion, the isolation of DNA from tumors and normal tissues, and the recovery of PBD-dimer 3 from PBD-DNA adducts provided new insights into the mechanism of action of PBD-dimer–containing ADCs. The marked difference in the levels of DNA adduct formation may suggest a potential of this ADC to widen the therapeutic window of highly cytotoxic agents like a DNA alkylator. Future preclinical studies can be conducted to help ensure that an ADC delivers adequate DNA alkylators to achieve tumor remission while simultaneously maintaining a safe level of DNA alkylation in normal organs. This example demonstrates that a thorough characterization of DNA alkylation, representing a disposition pathway of a DNA alkylator, is required for understanding the mechanism of action of an ADC carrying a DNA alkylator payload. In this study, a robust procedure for the quantification of DNA-PBD adducts and drug release was developed and applied to better understand the efficacy and tolerability of an ADC carrying a DNA-alkylating payload.
Experimental
Materials and Methods
Materials.
Calf thymus DNA was purchased from Rockland Immunochemicals Inc. (Pottstown, PA). Nuclease P1 was purchased from Sigma-Aldrich (St. Louis, MO). DNeasy Blood & Tissue Kit was purchased from Qiagen (Valencia, CA). All the other materials used were the same as described in the previous study (Zhang et al., 2016a).
Animal Study.
The procedures in animal study were described in detail previously, including the establishment of the subcutaneous xenograft mouse model; the administration of anti-CD22 THIOMAB antibody cyclobutyl-containing disulfide PBD-dimer and anti-CD22 cyclopropyl-containing disulfide PBD-dimer ADCs (1 and 2); and the collection of plasma, tumors, livers, lungs, and kidneys at 24 and 96 hours postdose (Zhang et al., 2016a). All the samples were kept frozen at −80°C until analysis.
DNA Extraction from Mouse Tissues and Plasma.
Mouse tissues (tumor, liver, lung, and kidney) were weighed and homogenized in 4× ice-cold phosphate-buffered saline by weight. The plasma samples were diluted with 4 volumes of ice-cold phosphate-buffered saline. DNA from 75-µl homogenates or 15-µl plasma was isolated by the DNeasy Blood & Tissue Kit (Qiagen) following the instructions with the following modifications. After the first loading of tissue lysate, the flow through was loaded two more times to ensure the best column binding of DNA. The columns were washed sequentially by two wash buffers containing 60–70% ethanol. In the last step, DNA on the column was eluted with 200 µl of water twice, and the eluates were combined.
DNA Digestion and Heating to Release PBD-Dimer 3.
Aliquots of 200 µl of DNA eluant were digested with 0.001 units of nuclease P1 at 37°C for 1 hour and then heated at 90°C for 30 minutes to release PBD-dimer. The postheating samples were aliquoted for the separate quantitation of PBD-dimer and DNA. The stability of PBD-dimer in the absence of DNA was also tested, and PBD-dimer 3 was found to be stable.
DNA and PBD-Dimer Quantitation.
Isolated DNA was quantitated by measuring the deoxyadenosine monophosphate generated from nuclease P1 digestion of calf thymus DNA as a standard by an LC-MS/MS method (unpublished data). Calf thymus DNA was dissolved in water at various concentrations, and the linear range was 5–5000 ng/ml for of DNA quantitation. The DNA isolated from tissue samples was diluted 200-fold before quantitation. The digestion was complete, as judged by the observation that no more deoxyadenosine monophosphate was produced with additional nuclease P1. The PBD-dimer 3 stock solutions were spiked into pure water or 100 µg/ml calf thymus solution to make standard curve samples to contain 0.2–7.8 nM PBD-dimer 3. The standard curve samples were incubated at 37°C for 1 hour to ensure the completion of DNA alkylation (Zhang et al., 2016a) before digestion and heating. The LC-MS/MS analysis showed that the presence of DNA did not impact the recovery and quantitation of PBD-dimer since two standard curves in water and DNA solutions were nearly superimposed. Quantitative recovery of PBD-dimer 3 from tissue DNA samples can be expected after the digestion and heating process.
Calculation of PBD-DNA Adducts.
The conversion of mass of nucleotides as determined by LC-MS/MS to the molarity of DNA was based on the average molecular weight of a DNA bp (of 650 Da). The final results were shown as the adduct numbers per million DNA bp. PBD-DNA adducts = PBD concentration/DNA concentration/650*106: that is, at 96 hours, PBD-DNA adducts = 192/(2.73/650*109)*106 = 45.7 PBD/106 bp.
Acknowledgments
The authors thank Melissa Schutten, Keyang Xu, Luna Liu, Geoffrey Del Rosario, Jintang He, Jack Sadowsky, Ben Shen, and Isabel Figueroa Amenabar for their technical contributions.
Authorship Contributions
Participated in research design: Ma, Khojasteh, Hop, and Zhang
Conducted experiments: Ma, Yu, and Zhang
Contributed new reagents or analytic tools: Erickson, Pillow, and Dragovich
Performed data analysis: Ma, Khojasteh, Hop, Polson, Wang, and Zhang
Wrote or contributed to the writing of the manuscript: Ma, Erickson, Polson, Pillow, Dragovich, and Zhang
Footnotes
- Received August 8, 2016.
- Accepted September 26, 2016.
Abbreviations
- ADC
- antibody drug conjugate
- bp
- base pair
- LC-MS/MS
- liquid chromatography tandem mass spectrometry
- MMAE
- maleimide-monomethyl auristatin E
- PBD
- pyrrolo[2,1-c][1,4]benzodiazepine
- Copyright © 2016 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}