


Abstract
Troglitazone (TRO) is an insulin sensitizer used in the treatment of type II diabetes. TRO is known to increase the activity of cytochrome P-450 (CYP) 3A in vivo. We have investigated the effect of TRO on CYP3A protein content and the activity of CYP3A (as measured by the formation of 6β-hydroxytestosterone formation) in primary cultures of human hepatocytes in comparison with rifampicin (RIF). Hepatocytes were isolated from four human livers by perfusion with collagenase, plated on collagen-coated plates, and maintained in William’s E medium. After 48 h in culture, cells were exposed to RIF (10 μM) or TRO (0–50 μM) twice, each over a period of 24 h, and the activity of CYP3A was measured. TRO increased the activity of CYP3A in a concentration-dependent manner, reaching a maximal response at 5 μM. Pretreatment of the hepatocytes with 10 μM TRO or 10 μM RIF resulted in a 4- to 15-fold increase in the activity of CYP3A. Maximum increase in CYP3A protein was observed at 5 μM TRO. There was a significant correlation (R2 = 0.89) between the content of immunoreactive CYP3A protein in the hepatocytes and the rate of formation of 6β-hydroxytestosterone. These results indicate that TRO is a potent inducer of CYP3A and is similar to RIF in inducing CYP3A in human hepatocytes. At concentrations of 25 μM and above, TRO was toxic to the cells, as determined by a decrease in the activity of CYP3A, a reduction in the amount of immunoreactive protein, and changes in the morphology of the cells.
Troglitazone (TRO)1 (Rezulin; Warner-Lambert Company) is an orally active insulin sensitizer of the thiazolidinedione class of compounds, indicated for the treatment of type II diabetes. TRO acts primarily by decreasing insulin resistance. It improves sensitivity to insulin in muscle and adipose tissue and inhibits hepatic gluconeogenesis. TRO is metabolized to sulfate and glucuronide conjugates and a quinone metabolite (Izumi et al., 1996). In vitro studies using expressed cytochrome P-450 (CYP) enzymes have shown that TRO is not metabolized by CYP3A (Rezulin package insert, 1998). However, TRO has been reported to increase the urinary excretion of 6β-hydroxycortisol and the ratio of 6β-hydroxycortisol to cortisol in the urine of healthy human subjects (Koup et al., 1998). This strongly suggests that TRO is an inducer of CYP3A.
Primary cultures of human hepatocytes represent a unique in vitro system to evaluate the induction of phase I and phase II enzymes involved in drug metabolism. Several investigators have successfully used human hepatocyte cultures to investigate the effect of various agents on the induction of CYP enzymes (Kostrubsky et al., 1995; Strom et al., 1996; Chang et al., 1997; Kostrubsky et al., 1998; reviewed inMaurel, 1996; Li, 1997).
The objective of the present study was to evaluate the effect of TRO on CYP3A protein content and the activity of CYP3A in primary cultures of human hepatocytes and to compare its effect to rifampicin (RIF), a well known inducer of CYP3A.
Materials and Methods
Chemicals.
William’s E medium and media supplements, dexamethasone and insulin, were obtained from BioWhittaker (Walkersville, MD). Gentamicin was obtained from Life Technologies Laboratories (Grand Island, NY). TRO was a gift from Parke-Davis (Ann Arbor, MI). Testosterone and 6β-hydroxytestosterone were obtained from Steraloids Inc. (Wilton, NH). Falcon culture dishes (60 mm) were obtained from Becton Labware (Franklin Lakes, NJ). Nitro blue tetrazolium/5-bromo-4-choloro-3-indolyl phosphate color-developing reagent and alkaline phosphatase-conjugated anti-rabbit antibody were purchased from Bio-Rad (Richmond, CA). All solvents and other chemicals used were of HPLC grade or the highest purity available.
Hepatocyte Cultures and Treatment Protocol.
Hepatocytes were isolated from four human liver donors. Table1 summarizes the medical history of the donors and the drugs to which the donors were exposed before organ donation. Hepatocytes were isolated by a three-step collagenase perfusion technique as described previously (Strom et al., 1996, 1998). The viability of the cells obtained, as measured by trypan blue exclusion test, ranged from 79 to 89%. In one case [human hepatocyte donor number (HH) 681], the hepatocytes were subjected to density gradient separation using Percoll to remove nonviable cells (Table 1). All the hepatocytes were plated in William’s E medium supplemented with 0.1 μM dexamethasone, 0.1 μM insulin, 0.05% gentamicin, and 10% bovine calf serum. Hepatocytes (3 × 106) were plated on 60-mm culture plates previously coated with rat tail collagen. Cells were allowed to attach for 4 to 6 h, at which time the medium was replaced by a serum-free medium containing all the supplements listed above. Medium was changed every 24 h. At 48 and 72 h, the cells were treated with fresh medium containing RIF (10 μM) or different concentrations of TRO (0–50 μM; n = 3 at each concentration) dissolved in dimethyl sulfoxide (DMSO). The final concentration of DMSO in the medium was 0.1%. The cells were maintained in culture at 37°C in a humid atmosphere containing 5% CO2 and 95% air throughout the experiment.
Donor information
Measurement of CYP3A Activity.
After 96 h in culture, the hepatocytes were washed with fresh medium and incubated for an hour and then exposed to a medium containing testosterone (100 μM) in acetonitrile (final concentration in the media was 0.4%) for 30 min. At this time point, the medium was harvested and stored at −20°C. 6β-hydroxytestosterone in the medium was measured by an HPLC method as described by Waxman et al. (1983) and Crespi and Pennman (1997), with the following modifications. One hundred microliters of the culture medium was injected into a LiChrospher 100 RP-18 column (4.6 × 250 mm, 5 μm). A mobile phase of methanol/water (60:40, v/v) at a flow rate of 1.2 ml/min was used to separate various components. 6β-hydroxytestosterone was measured at 254 nM by comparing the absorbance to a standard curve of 6β-hydroxytestosterone in William’s E medium. Because all the inducers were dissolved in DMSO, we also evaluated the potential effect of exposing the cells to DMSO (0.1%) for 48 h on CYP3A activity.
Measurement of the Expression of CYP3A4/5 Protein.
At the end of the sampling period, the cells were harvested in phosphate buffer (0.1 M, pH 7.4) and stored at −20°C for protein determination (Lowry et al., 1951) and analysis of immunochemically reactive CYP3A4/5. Immunochemical analysis of CYP3A4/5 protein was performed as described previously (Kostrubsky et al., 1995) using 15 μg of pooled sonicated proteins. CYP3A4/5 was detected with a rabbit antihuman CYP3A antibody, which detects both CYP3A4 and CYP3A5 proteins (Kostrubsky et al., 1995, 1998). An alkaline phosphatase-conjugated antirabbit antibody and Nitro blue tetrazolium/5-bromo-4-choloro-3-indolyl phosphate-developing reagents were used to visualize the blots.
Statistical Analysis.
The data were analyzed using ANOVA followed by post hoc Duncan test for group-wise comparison. The results were considered statistically significant if p ≤ .05.
Results
Increase in the Activity of CYP3A by TRO and RIF.
The effect of RIF (10 μM) and TRO (0–25 μM) on CYP3A activity as measured by the rate of formation of 6β-hydroxytestosterone in four cultures of human hepatocytes are shown in Figs.1, A to D.

Effect of TRO on CYP3A activity as measured by the formation of 6β- hydroxytestosterone (pmol/min/mg protein).
Hepatocytes prepared from human donors were treated for 48 h with 0 to 50 μM TRO or 10 μM RIF. At the end of this time period, the medium was changed and testosterone (100 μM) was added to the cells. After 30 min of incubation, aliquots of the medium were removed and the amount of 6β-hydroxytestosterone was determined as described in Materials and Methods. Each value represents the mean ± S.D. of triplicate treatments. There was no difference between untreated cells and cells treated with 0.05 to 0.1 μM TRO. ∗, Significantly different from untreated cells and 0.05, 0.1 μM TRO with p < .05. ∗∗, Significantly different from cells treated with 0.5 and 1 μM TRO with p< .05. ∗∗∗, Significantly different from 10 μM TRO withp < .05.
In HH673 (Fig. 1A), the rate of formation of 6β-hydroxytestosterone in the untreated cells was 0.15 ± 0.02 nmol/min/mg protein. Treatment with TRO at 1 to 25 μM resulted in a 1.3- to 3.9-fold increase (Table 2) in the formation rate of 6β- hydroxytestosterone compared with the untreated cells. In the same culture, RIF (10 μM) caused a 3.5-fold increase in the formation rate of 6β-hydroxytestosterone as compared with the untreated cells. There were no significant differences between the effect of TRO (1–10 μM) and RIF (10 μM) on the formation of 6β-hydroxytestosterone.
Effect of TRO and rifampicin on the activity of CYP3A
Because the treatment with 1 μM TRO resulted in an almost maximum increase in testosterone 6β-hydroxylase activity in culture HH673, we investigated the effect of lower concentrations of TRO on the activity of CYP3A in hepatocyte cultures from three additional donors. In HH676 (Fig. 1B), HH677 (Fig. 1C), and HH681 (Fig. 1D), the rates of formation of 6β-hydroxytestosterone in the untreated cells were 0.05 ± 0.02, 0.09 ± 0.01, and 0.13 ± 0.03 nmol/min/mg protein, respectively. Treatment with 0.05 and 0.1 μM TRO did not cause significant increases in the activity of CYP3A in any of the cultures (Table 2) as compared with the untreated cells. However, treatment with 0.5 to 25 μM TRO caused 7- to 13.4-, 5.1- to 9.4-, and 2.8- to 6.5-fold increases in the formation of 6β-hydroxytestosterone in HH676, HH677, and HH681, respectively. The formation rate of 6β-hydroxytestosterone after treatment with TRO (0.5–25 μM) or RIF (10 μM) was significantly higher than that observed in untreated cells. There were no significant differences in the formation rates of 6β-hydroxytestosterone in cells treated with TRO (1–10 μM) or RIF (10 μM).
In all of the cultures studied, treatment with TRO at 25 μM resulted in a decrease in the activity of CYP3A and the content of immunoreactive CYP3A protein. Exposure of the cells to 50 μM TRO was lethal to the hepatocytes within 24 h as judged by microscopic examination of the hepatocytes. CYP3A activity could not be determined in these cells.
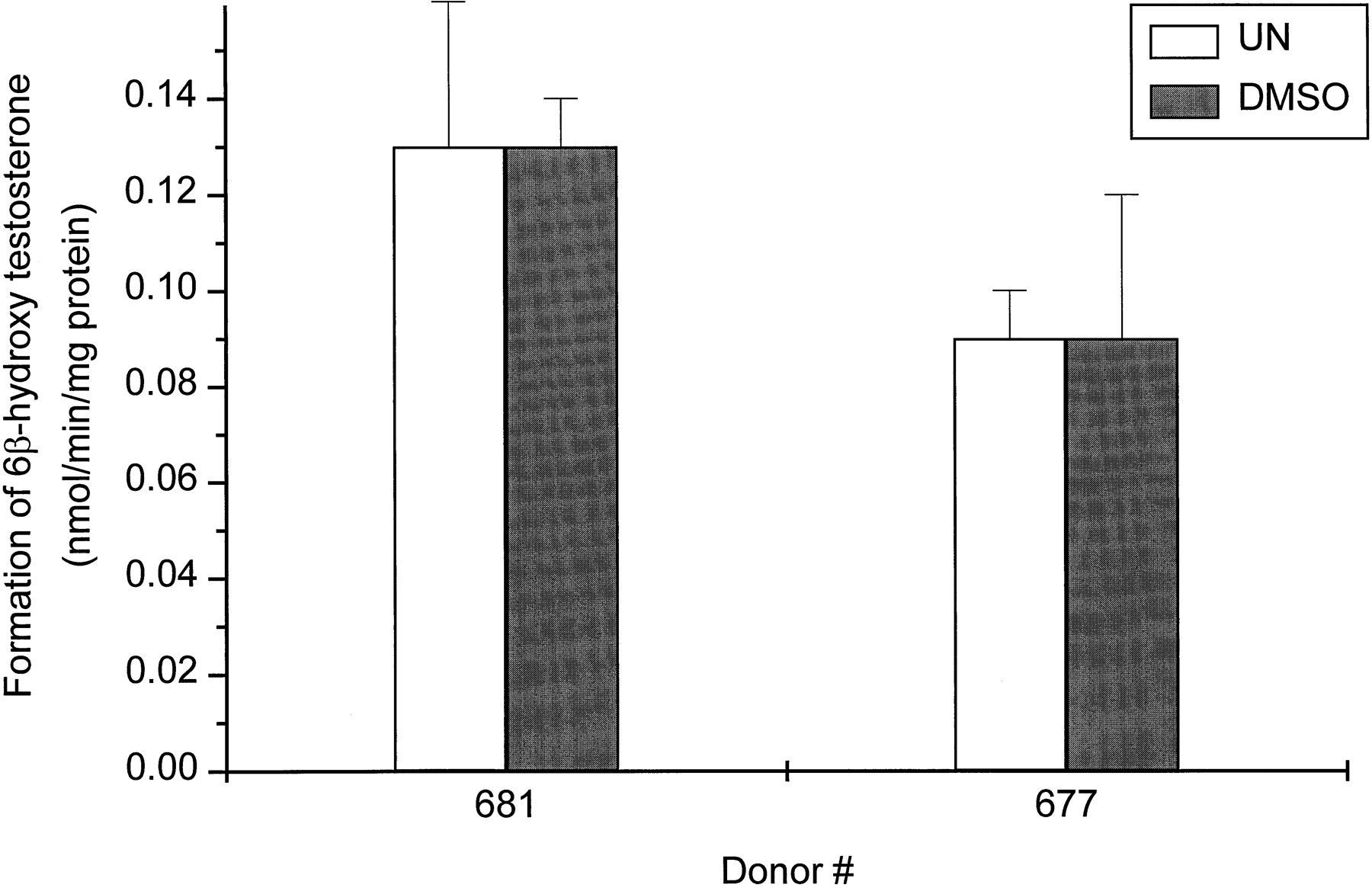
Figure 2 shows the effect of DMSO on testosterone 6β-hydroxylase activity in two cultures of human hepatocytes. There were no significant differences in formation rates of 6β-hydroxytestosterone between cells exposed to 0.1% DMSO for 48 h and cells that were not exposed to any DMSO.
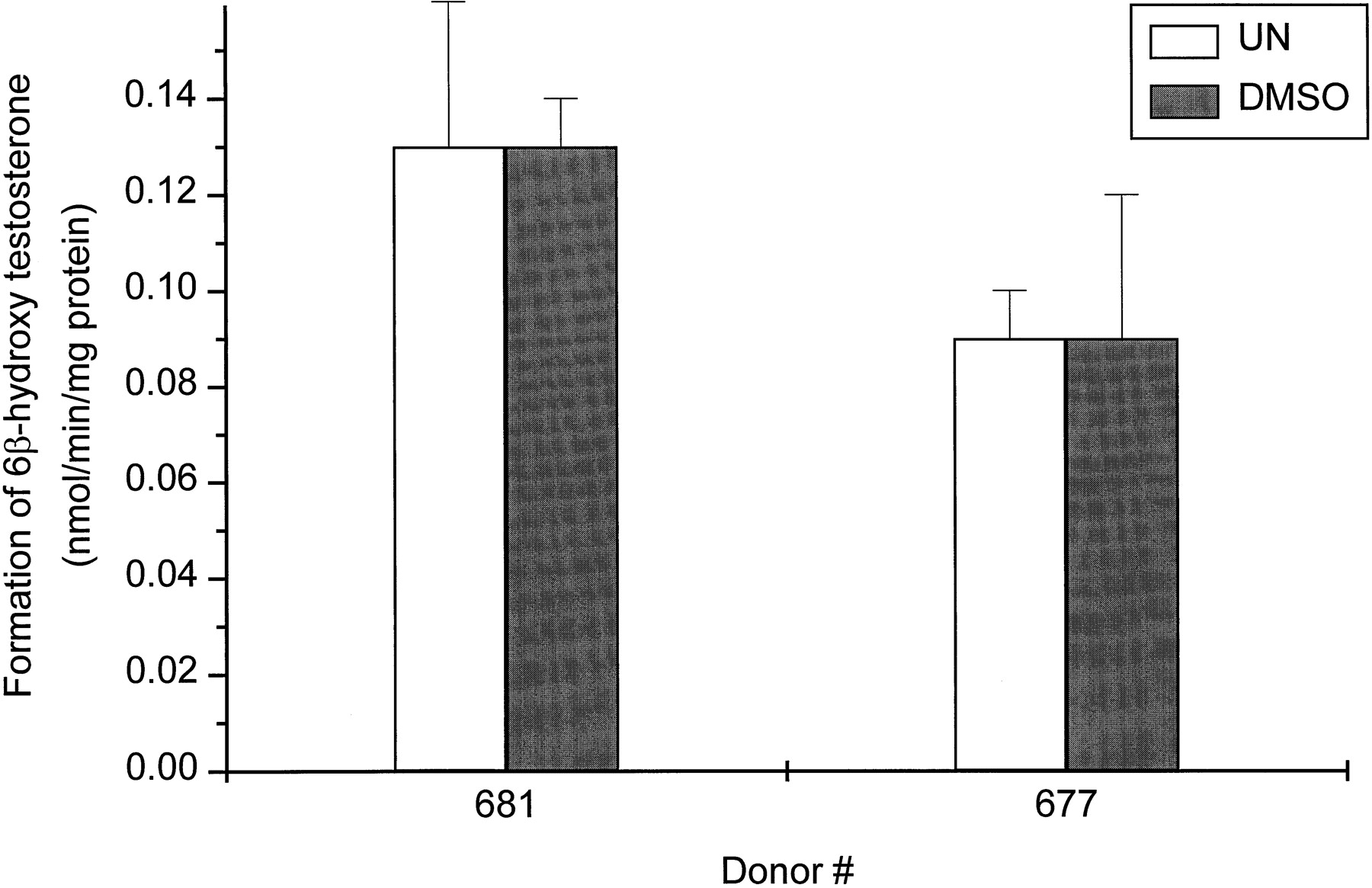
Effect of DMSO on the CYP3A activity as measured by the formation of 6β-hydroxytestosterone (pmol/min/mg protein).
Hepatocytes from two donors (HH677 and HH681) were treated for 48 h with 0.1% DMSO. At the end of this time period, the medium was changed and testosterone (100 μM) was added to the cells. After 30 min of incubation, aliquots of the medium were removed and the amount of 6β-hydroxytestosterone was determined as described inMaterials and Methods. Each value represents the mean ± S.D. of triplicate treatments. There was no significant difference between untreated (UN) and DMSO-treated hepatocytes.
Immunochemical Analysis of CYP3A4/5 Protein.
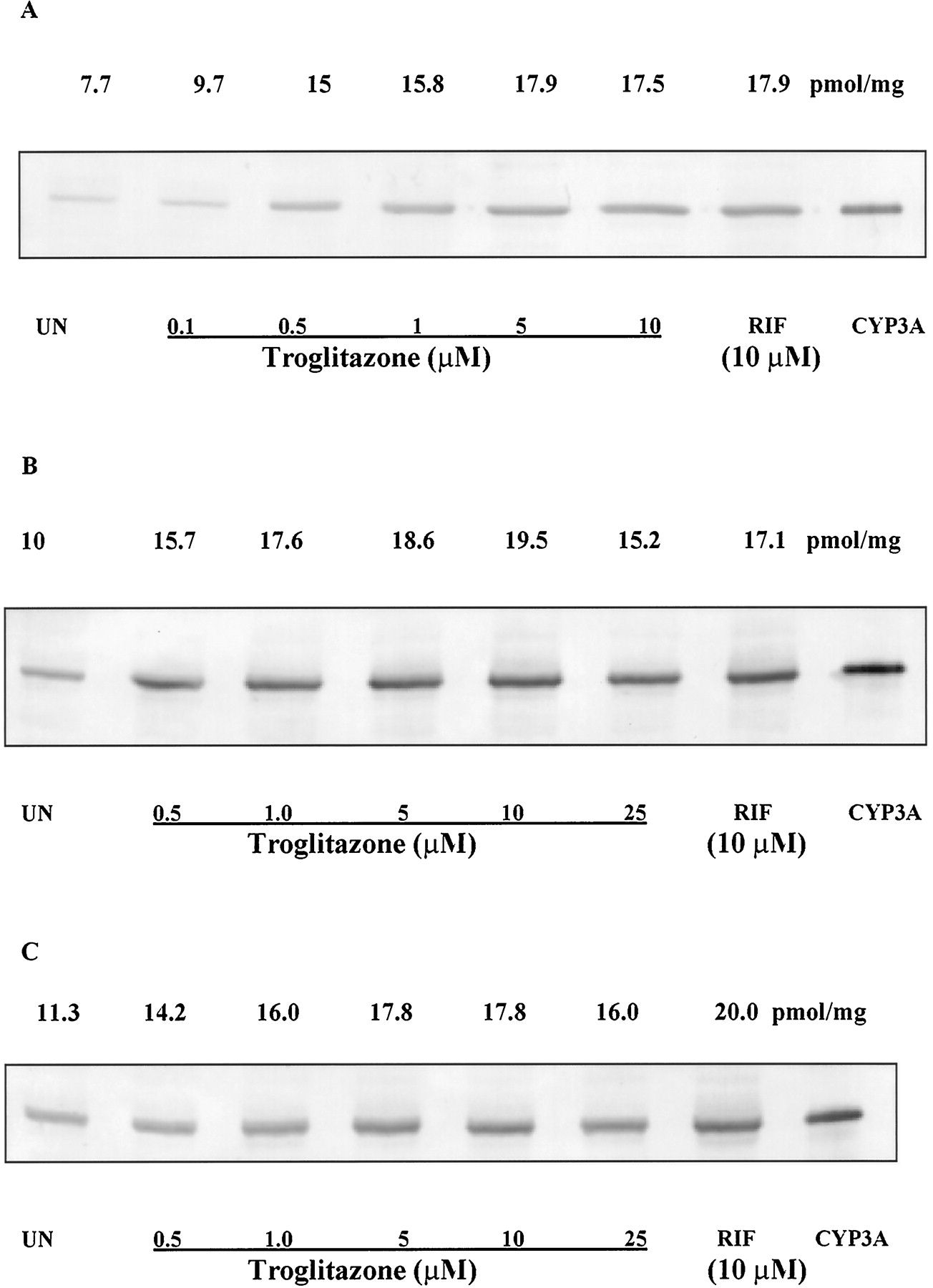
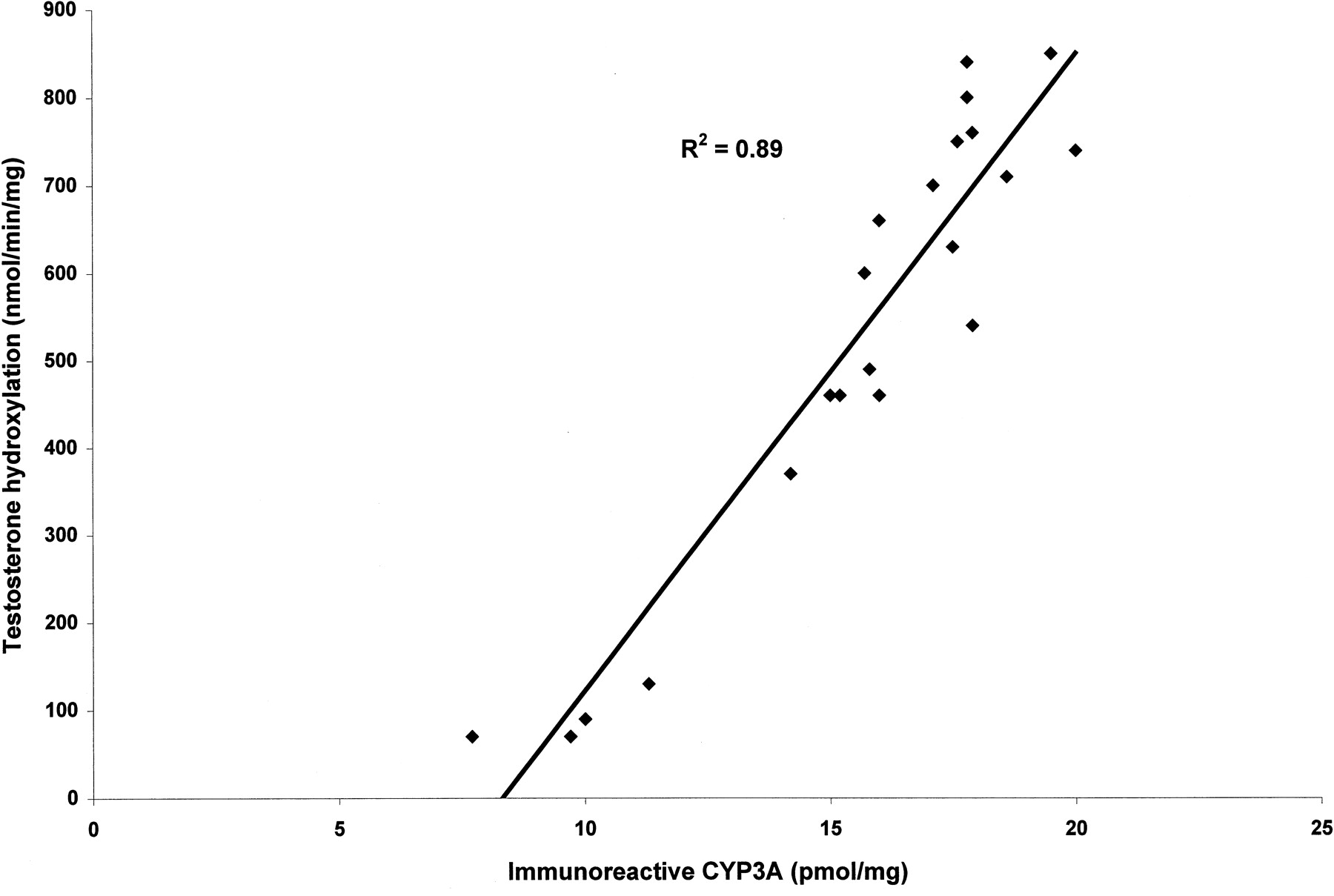
We examined the effect of TRO and RIF on the immunoreactive CYP3A4/5 protein from three separate human hepatocyte cultures. Cells from three wells for each culture were pooled and analyzed for immunoreactive CYP3A4/5. TRO (0.1–10 μM) increased the amount of immunoreactive CYP3A protein in a concentration-dependent manner (Fig.3). The maximum amount of CYP3A protein achieved with TRO (5 μM) treatment was comparable with the amount of CYP3A protein in the wells treated with 10 μM RIF. There was a good correlation (R2 = 0.89) between the amount of immunoreactive CYP3A in human hepatocytes and the activity of CYP3A (Fig. 4). At a concentration of 25 μM, TRO decreased the amount of immunoreactive CYP3A protein in all of the cultures.

Effect of TRO on the amount of immunochemical CYP3A from three donors.
Hepatocytes were treated for 48 h with the concentration of TRO indicated in the figure, and immunodetectable CYP3A levels were analyzed in sonicates of the whole cell as described inMaterials and Methods. Sonicated protein (15 μg) was applied per well. Expressed CYP3A at 0.2 pmol was used as control. A, HH676; B, HH677; C, HH681. The numbers on the top of each panel are the amounts of CYP3A protein (pmol/mg protein) after each treatment.
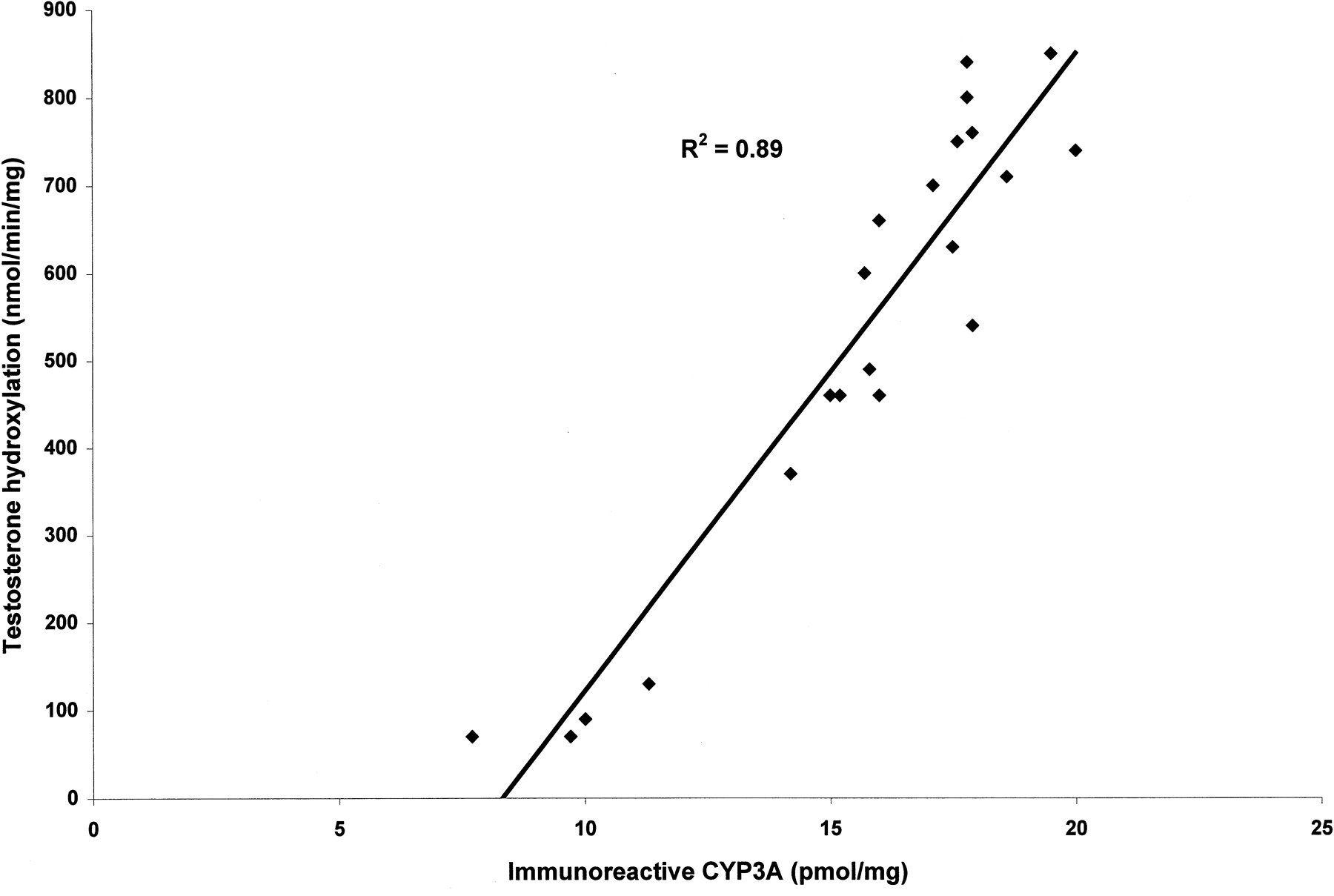
Correlation between the amount of immunoreactive CYP3A protein (pmol/mg protein) and CYP3A activity (pmol/min/mg protein) in three donors (R2 = 0.89).
The activity of CYP3A was determined by the formation of 6β-hydroxytestosterone in untreated and treated hepatocytes; the amount of immunoreactive protein was measured in the pooled samples of each treatment as quantitated by Western blot.
Discussion
Human hepatocyte cultures are useful in evaluating the effect of a chemical or drug on the mRNA expression, amount of immunoreactive proteins, and the activity of CYP enzymes. RIF is one of the most potent inducers of CYP3A (Venkatesan, 1992) and has been used as a positive control in studies that evaluate the induction of CYP3A in human hepatocytes (Pichard et al., 1990; Schuetz et al., 1993;Kostrubsky et al., 1995, 1998; Li, 1997; Li et al., 1997). In primary cultures of human hepatocytes, treatment with RIF (10 μM) for 48 h results in maximal increase in the activity of CYP3A, as measured by the formation of 6β-hydroxytestosterone (Li, 1997). In the present study, RIF (10 μM) treatment resulted in a 4- to 15-fold increase in CYP3A activity, which is in agreement with the data published in the literature.
In all of the cultures tested, TRO was a potent inducer of CYP3A-mediated testosterone 6β-hydroxylation. TRO-mediated increases in the activity of CYP3A and the immunoreactive CYP3A protein levels were similar to that observed with RIF (10 μM) treatment. Donor hepatocytes that exhibited the maximum induction by RIF also showed the greatest induction with TRO (HH676), and hepatocytes with the lowest response to RIF showed the lowest induction with TRO (HH673).
On day 5, the activities of CYP3A in the untreated cells were different between the four cultures tested. Untreated hepatocytes from donors HH673, HH681, and HH677 were exposed to CYP3A inducers (phenytoin and dexamethasone) before the liver donation. In the untreated cultures, hepatocytes from HH673 showed the highest CYP3A activity, followed by HH681 and HH677. There was a 4- to 13-fold increase in the formation of 6β-hydroxytestosterone in these cultures after treatment with TRO. This increase in enzymatic activity corresponded to a 2.3-fold increase in immunoreactive CYP3A protein. The increase in the CYP3A protein amount was not of the same magnitude as that of the activity. The exact reason(s) for such an observation is not clear at this time. It may relate to the fact that there could have been an underestimation of the CYP3A protein content in the cells due to an interference in the assay used because of the presence of other cellular components. It may reflect increased activity of other enzymes that produce 6β-hydroxytestosterone that are also induced by rifampin. Additional studies are essential to completely characterize this phenomenon.
Our observations suggest that TRO will increase the in vivo intrinsic clearance of other substrates of CYP3A. Concentrations of TRO that cause induction in the primary hepatocyte cultures are similar to the plasma concentrations observed in patients on TRO therapy (Young et al., 1998). Our observations are consistent with published studies that reported a 2-fold increase in the urinary excretion of 6β-hydroxycortisol and the ratio of 6β-hydroxycortisol to total cortisol, a measure of CYP3A activity, in healthy human subjects treated with TRO (Koup et al., 1998). TRO has also been shown to decrease the trough concentrations of cyclosporine (a substrate for CYP3A) in stable renal transplant patients (Kaplan et al., 1998). TRO has been reported to decrease the plasma concentrations of terfenadine and oral contraceptives by 30% (Rezulin package insert, 1998).
We have observed a more pronounced increase (4- to 13-fold) in the activity of CYP3A in vitro than what is reported in most of the in vivo studies. TRO is highly bound to plasma proteins (>99%) and the high plasma protein binding of TRO may partially be responsible for the lower magnitude of induction (due to a lower amount of unbound drug) observed in vivo. TRO is highly distributed into a number of tissues including the liver in rats, mice, and dogs (Kawai et al., 1997). Factors such as the concentration of TRO in the liver tissues, the plasma protein binding of TRO, and the possible effect of TRO on the hepatic blood flow may also contribute to the observed differences in in vitro and in vivo studies.
The effect of TRO on other CYP or phase II enzymes is not completely characterized at this time. TRO appears to have no effect on prothrombin time when coadministered to patients on warfarin, suggesting that TRO has no effect on CYP2C9. TRO has also been shown not to alter the pharmacokinetics of acetaminophen, suggesting a lack of effect of TRO on phase II pathways (Rezulin package insert, 1998).
Use of TRO has been associated with severe hepatic dysfunction leading to hepatic failure in few patients (Watkins and Whitcomb, 1998; Gitlin et al., 1998; Neuschwander-Tetri et al., 1998). Hepatotoxicity can be evaluated in primary cultures of human hepatocytes by various end points including morphological changes, biochemical changes, or changes in enzymatic activity (Guillouzo et al., 1997). In the present study, 50 μM TRO was lethal to the cells (morphological). At 25 μM TRO, toxicity to the cells was evident as noted by a reduction in the rate of formation of 6β-hydroxytestosterone and amount of CYP3A protein. We have reported that a decrease in the inducible level of the immunoreactive CYP3A protein and testosterone 6β hydroxylase activity after exposure to increasing concentrations of Taxol correlates well with toxicity to the hepatocytes as measured by the incorporation of C14-labeled leucine into newly synthesized protein (Kostrubsky et al., 1999). Therefore, the data presented here indicates that concentrations greater than 25 μM TRO are toxic to hepatocytes in culture.
In summary, we have demonstrated that TRO is a potent inducer of CYP3A. The increases in the immunoreactive CYP3A protein and the activity of CYP3A are comparable to that caused by RIF. Primary cultures of human hepatocytes are useful to evaluate induction of hepatic CYP3A and to determine the hepatotoxic potential of new chemicals.
Acknowledgments
We thank Dr. Adedayo A. Adedoyin of Zeneca Pharmaceuticals and Dr. Ram P. Kapil of Forest Laboratories Inc. for their comments on the manuscript. We also thank the Liver Tissue Procurement and Distribution System (LTPADS) for the support for tissue and cells.
Footnotes
-
Send reprint requests to: Raman Venkataramanan Ph.D., 718 Salk Hall, School of Pharmacy, University of Pittsburgh, Pittsburgh, PA 15261. E-mail: rv+{at}pitt.edu
-
This work was supported in part by summer fellowships from the University of Pittsburgh Pharmacy Associates (V.R., B.J.K.), a Postdoctoral Fellowship from Lilly Research Laboratories (V.E.K), Anatomic Gift Foundation (K.D., J.E.E., S.C.S), Merck Scholar Program Award (B.J.K.), and National Institutes of Health N01 DK-92310 (S.C.S.).
- Abbreviations used are::
- TRO
- troglitazone
- CYP
- cytochrome P-450
- HH
- human hepatocyte donor number
- RIF
- rifampicin
- DMSO
- dimethyl sulfoxide
- Received March 26, 1999.
- Accepted June 28, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}