


Abstract
We previously demonstrated that multidrug resistance protein 3 (Mrp3/ABCC3) is induced in rat liver by phenobarbital (PB) and several other microsomal enzyme inducers that induce cytochrome P450 2B (CYP2B). CYP2B is induced by constitutive androstane receptor (CAR)-retinoid X receptor (RXR) heterodimer binding to a phenobarbital-responsive promoter element in the CYP2B promoter. Hepatic mRNA levels of CYP2B and Mrp3 were measured in three models of altered CAR activity to determine whether CAR is also involved in the induction of Mrp3. In Wistar Kyoto rats, where males express higher CAR protein levels than females, the induction of CYP2B1/2 was significantly higher in males than in females by PB, diallyl sulfide, and trans-stilbene oxide but not oltipraz. Mrp3 was induced by each of these treatments, but in contrast to CYP2B1/2, to a similar magnitude in males and females. In male hepatocyte-specific RXRα-/- mice, CYP2B10 was not induced by diallyl sulfide or oltipraz but remained inducible by PB and trans-stilbene oxide after considering the decrease in basal CYP2B10 expression. Mrp3, however, was induced by PB, diallyl sulfide, trans-stilbene oxide and oltipraz in both wild-type and RXRα-/- mice. Additionally, constitutive expression of Mrp3 was significantly reduced in RXRα-/- mice. In CAR-/- mice, the robust induction of CYP2B10 by PB was completely absent. However, Mrp3 was equally induced both in wild-type and CAR-/- mice by PB. These data clearly demonstrate that induction of hepatic Mrp3 by PB and other microsomal enzyme inducers is CAR-independent and implies a role for RXRα in the constitutive expression of Mrp3.
Multidrug resistance protein 3 (Mrp31) is an ATP-dependent xenobiotic transporter that is predominantly expressed in the intestine, where it is thought to be involved in bile acid absorption (Cherrington et al., 2002; Rost et al., 2002). In liver, Mrp3 expression is normally very low and localized to the basolateral or sinusoidal membrane of hepatocytes where it serves to export a wide range of organic anions from the liver, back to the blood, thereby decreasing hepatic exposure and toxicity to liver (Donner and Keppler, 2001; Soroka et al., 2001).
The ability of certain compounds, termed microsomal enzyme inducers, to increase specific enzyme systems that result in an increased metabolic capacity has been widely studied. Many Phase I (oxidative and reductive) and Phase II (conjugative) enzymes have been identified as being transcriptionally regulated by these inducers. Along with Phase I and Phase II enzyme induction, pretreatment with several microsomal enzyme inducers have been shown to alter the excretion of xenobiotics, which implies that transport processes may also be similarly regulated (Klaassen and Plaa, 1968; Klaassen, 1970, 1974). Determining whether these microsomal enzyme inducers coordinately regulate transporter genes would add to our understanding of the disposition of xenobiotics.
One mechanism of transcriptional regulation that has been well studied is the activation of the constitutive androstane receptor (CAR). CAR is a member of a family of nuclear orphan receptors and has been shown to bind specific promoter sequences (called phenobarbital-responsive enhancer modules), and activate transcription (Honkakoski et al., 1998a; Waxman, 1999). Specifically, CAR has been shown to heterodimerize with retinoid X receptor (RXR) and bind to a phenobarbital-enhancer element on the CYP2B gene and increase transcription (Honkakoski et al., 1998b). Because of its function as the obligate heterodimeric partner, RXR is essential for CAR activity. The predominant phenobarbital-inducible CYP2B genes in mouse and rat are CYP2B10 and CYP2B1/2, respectively, and therefore commonly used as a positive indicator for CAR activation. CAR is highly expressed in liver and activates gene transcription constitutively in cultured cell lines (Forman et al., 1998). In hepatocytes, however, CAR protein is normally retained in the cytoplasm and is unable to activate expression of CYP2B10 or other target genes (Kawamoto et al., 1999), unless PB or other structurally unrelated microsomal enzyme inducers induce translocation of CAR to the nucleus (Sueyoshi and Negishi, 2001). CAR has been described as a cellular sensor that is capable of responding to chemical exposure (Honkakoski et al., 1998b; Kawamoto et al., 1999; Sueyoshi et al., 1999). By combining the coordinate regulation of both Phase I and II metabolism in liver with the transport by Mrp3, an organism may be greatly advantaged by reducing hepatotoxicity during chemical exposure.
We have previously shown that rat hepatic Mrp3 is induced by a diverse group of chemicals including PB, diallyl sulfide, oltipraz, and trans-stilbene oxide, which all share a common trait as potent CYP2B inducers (Cherrington et al., 2002). Therefore, the aim of this study was to determine the role of CAR in the induction of Mrp3 by the microsomal enzyme inducers that are known to induce CYP2B. Three separate models have been used to compare the CAR-mediated activation of Mrp3 and CYP2B: a CAR-deficient Wistar Kyoto (WKY) rat strain model, a hepatocyte-specific RXRα-/- model, and a CAR-/- model. A gender-selective protein level of CAR has been implicated in the sexually dimorphic induction of CYP2B by phenobarbital in the livers of WKY rats (Yoshinari et al., 2001). In this strain, male rats express much higher levels of cytosolic and nuclear CAR protein, and CYP2B1 is fully inducible by PB, whereas female CYP2B1 levels are only slightly induced because they are CAR deficient. The RXRα-/- and CAR-/- models were genetically engineered to lack the specific protein required for CAR-RXRα heterodimerization. The CAR-/- mice are a general knockout, whereas the RXRα-/- mice are devoid of RXRα only in hepatocytes.
Materials and Methods
Chemicals. Oltipraz was a gift from Dr. Ronald Lubet (National Cancer Institute, Bethesda, MD). Trans-stilbene oxide was purchased from Aldrich Chemical Co. (Milwaukee, WI). All other chemicals were purchased from Sigma-Aldrich (St. Louis, MO).
Animals. Five male or female Wistar Kyoto rats per group (200-250 g; Harlan, Indianapolis, IN) were acclimated to the housing facility (2-3 rats/cage, 50% relative humidity, 12-h light/dark cycle) for 1 week and fed Teklad 8604 rodent chow (Harlan Labs, Madison, WI). Treatment groups were as follows: PB (80 mg/kg, i.p. in saline), diallyl sulfide (DAS, 500 mg/kg, i.p. in corn oil), trans-stilbene oxide (TSO, 200 mg/kg, i.p. in corn oil, twice daily), ethoxyquin (EQ, 150 mg/kg, p.o. in corn oil), oltipraz (OPZ, 150 mg/kg, p.o. in corn oil), corn oil (i.p.), corn oil (p.o.), and saline (i.p.). All animals were treated for 4 days with an injection volume of 5 ml/kg. Male hepatocyte-specific RXRα-/- mice (Wan et al., 2000) or corresponding age-matched controls were treated as follows: PB (80 mg/kg, i.p. in saline), DAS (200 mg/kg, i.p. in corn oil, 3 days), TSO (200 mg/kg, i.p. in corn oil, twice daily/3 days), OPZ (150 mg/kg, p.o. in corn oil), corn oil (i.p.), corn oil (p.o.), or saline (i.p.). Unless otherwise noted, all animals were treated for 4 days with an injection volume of 5 ml/kg. All livers were excised 24 h after the final treatment, snap-frozen in liquid nitrogen, and stored at -80°C until use. Male CAR-/- mice (Wei et al., 2000) or C57BL/6 mice were treated with either corn oil or PB (100 mg/kg, i.p. in saline, 24 h), and total RNA was prepared from livers.
Total RNA was isolated using RNAzol B reagent (Tel-Test Inc., Friends-wood, TX) per the manufacturer's protocol. RNA concentration was determined by UV spectrophotometry, and its integrity was examined by ethidium bromide staining after agarose gel electrophoresis.
Development of Specific Oligonucleotide Probe Sets for bDNA Analysis. Rat CYP2B1/2 and Mrp3 probes were used as previously described by Hartley and Klaassen (2000) and Cherrington et al. (2002), respectively. The mouse CYP2B10 probe set was designed against GenBank accession number NM_009998 and has some cross-reactivity with CYP2B9, 2B13, and 2B19. The mouse Mrp3 probe set was designed against GenBank accession number BI330408, an EST, which displays 91% homology to the rat Mrp3 sequence. These target sequences (Table 1) were analyzed by ProbeDesigner Software Version 1.0 (Bayer Corp., Emeryville, CA). All oligonucleotide probes were designed with a Tm of approximately 63°C enabling hybridization conditions to be held constant (i.e., 53°C) during each hybridization step and for each oligonucleotide probe set. Every probe developed in ProbeDesigner was submitted to the National Center for Biotechnological Information (NCBI) for nucleotide comparison by the basic logarithmic alignment search tool (BLASTn) to ensure minimal cross-reactivity with other known mouse sequences and ESTs.
List of oligonucleotide probes generated for analysis of mRNA expression
Branched DNA Assay. Specific CYP2B or Mrp3 oligonucleotide probes were diluted in lysis buffer supplied in the QuantiGene HV signal amplification kit (Bayer Corp.-Diagnostics Div., Tarrytown, NY). All reagents for analysis (i.e., lysis buffer, capture hybridization buffer, amplifier/label probe buffer, and substrate solution) were supplied in the QuantiGene HV signal amplification kit. Total RNA (1 μg/μl, 10 μl) was added to each well of a 96-well plate containing 50 μl of capture hybridization buffer and 50 μl of a diluted probe set. Total RNA was allowed to hybridize to each probe set overnight at 53°C. Subsequent hybridization steps were carried out according to the manufacturer's protocol, and luminescence was measured with a Quantiplex 320 bDNA luminometer interfaced with Quantiplex data management software version 5.02 (Bayer Corp.-Diagnostics Div.) for analysis of luminescence from 96-well plates.
Statistical Analysis. Data are expressed as mean ± standard error. For multiple comparisons, analysis of variance was performed followed by Duncan's multiple range test. Gender and constitutive differences were determined using Student's t test. The level of significance was set at p < 0.05.
Results
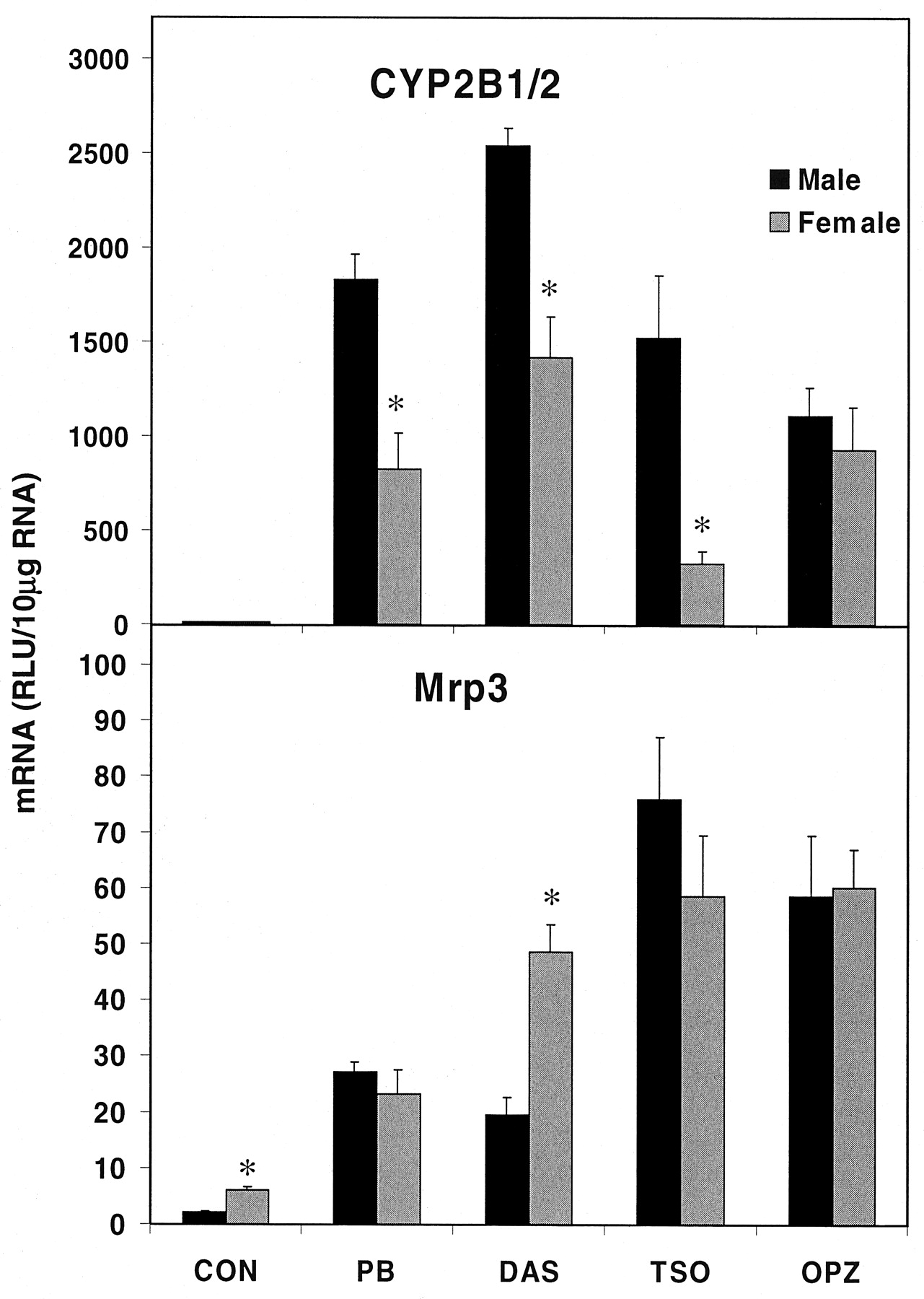
Mrp3 Expression in the Sexually Dimorphic Wistar Kyoto Rat. The expression levels of hepatic CYP2B1/2 and Mrp3 were determined in male and female Wistar Kyoto rats treated with phenobarbital, diallyl sulfide, trans-stilbene oxide, and oltipraz. Wistar Kyoto rats have been shown to be sexually dimorphic in their induction of CYP2B1 mRNA in liver by phenobarbital and the levels of hepatic CAR protein, with male rats having higher CAR protein and greater CYP2B1 induction than female rats (Yoshinari et al., 2001). In the present study, CYP2B1/2 mRNA levels were significantly increased by all four treatments in both male and female rats (Fig. 1). As anticipated, CYP2B1/2 induction was significantly higher in male Wistar Kyoto rats than in female rats treated with phenobarbital (122%), diallyl sulfide (79%), and trans-stilbene oxide (369%). No gender difference was observed in the CYP2B1/2 induction by oltipraz.

Induction of hepatic CYP2B1/2 and Mrp3 mRNA in male and female WKY rats.
Hepatic CYP2B1/2 or Mrp3 mRNA levels were measured in male and WKY rats treated with the following inducers: controls corn oil (i.p. and p.o.) and saline (CON, grouped together), PB, DAS, TSO, and OPZ. Units are expressed as relative light units (RLU) ± S.E.M. with RNA-independent “background” subtracted.
Similarly, Mrp3 mRNA levels were also significantly increased by each of the treatments (Fig. 1). However, unlike CYP2B1/2, there were no differences observed between genders in the increased expression of Mrp3 except for diallyl sulfide, where females actually have a higher level. Also, within the control animals, female constitutive expression of Mrp3 was significantly higher than in males.
Mrp3 Expression in RXRα-Deficient Mice. The expression levels of hepatic CYP2B10 and Mrp3 were also measured in hepatocyte-specific RXRα-/- and wild-type mice in response to phenobarbital, diallyl sulfide, trans-stilbene oxide, and oltipraz administration (Fig. 2). RXRα is thought to be the most important RXR subtype required for efficient heterodimerization and CAR-mediated induction of CYP2B. Hepatic CYP2B10 levels were significantly induced by each treatment in the wild-type mice. However, the induction of CYP2B10 by diallyl sulfide and oltipraz was absent in the RXRα-/- mice, but CYP2B10 remained inducible in the RXRα-/- mice by phenobarbital and trans-stilbene oxide, although to a lesser degree than in wild-type mice. This observation corresponds to previously published data showing that CYP2B10 remains inducible by the potent CAR activator TCPOBOP in the absence of RXRα-/- (Cai et al., 2002). Constitutive CYP2B10 expression was also significantly lower in vehicle-treated RXRα-/- mice.

Induction of CYP2B10 and Mrp3 in RXRα-/-mice.
Hepatic CYP2B1/2 or Mrp3 mRNA levels were measured in wild-type and RXRα-/- mice treated with the following inducers: controls corn oil (i.p. and p.o.) and saline (CON, grouped together, n = 6), PB, DAS, TSO, and OPZ (n = 4). Units are expressed as relative light units (RLU) ± S.E.M. with RNA-independent “background” subtracted. *, significantly different from vehicle; †, significantly different from wild-type vehicle.
Interestingly, mRNA levels of Mrp3 were reduced more in the vehicle-treated RXRα-/- mice than in the corresponding wild-type mice, suggesting a possible role for RXRα in the constitutive expression of Mrp3. Also, Mrp3 mRNA levels were significantly increased by PB, diallyl sulfide, trans-stilbene oxide, and oltipraz in both wild-type (182, 94, 133, and 144%) and RXRα-/- mice (185, 174, 187, and 268%), respectively.
Mrp3 Expression in CAR-Deficient Mice. As a positive control for CAR-mediated induction, treatment of wild-type mice with phenobarbital resulted in more than an 8000% increase in CYP2B10 mRNA (Fig. 3). This robust induction was completely absent in the CAR-/- animals treated with phenobarbital, as has been previously noted (Wei et al., 2000).
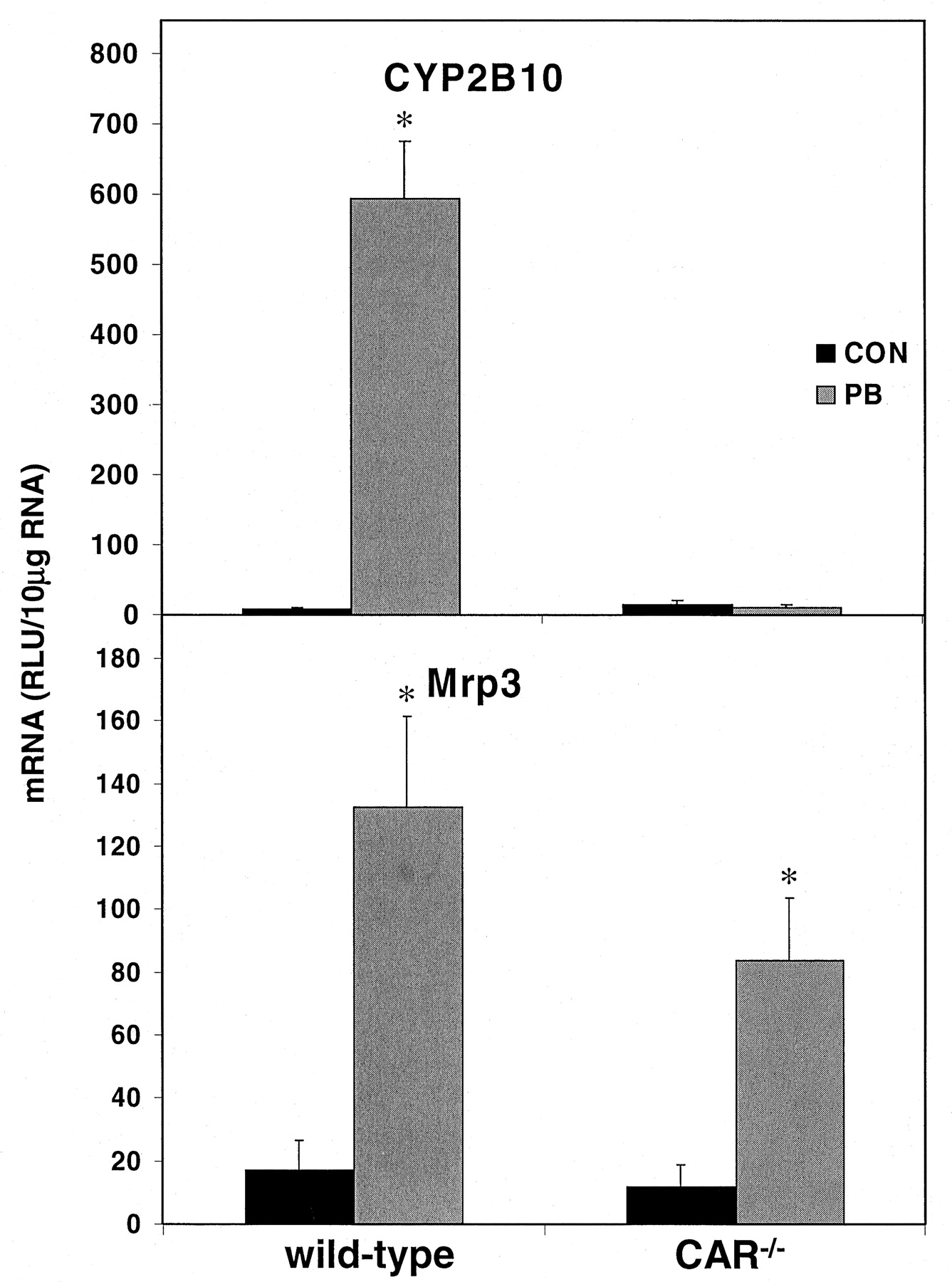
Induction of CYP2B10 and Mrp3 in CAR-/-mice.
Total RNA from male wild-type and CAR-/- mice (n = 5) treated with saline or PB was subjected to the bDNA assay in the presence of CYP2B10 or Mrp3 probe sets, respectively. Units are expressed as relative light units (RLU) ± S.E.M. with RNA-independent “background” subtracted. *, significantly different from vehicle.
Similarly, Mrp3 was also induced by phenobarbital in the wild-type mice (670%). However, unlike CYP2B10, Mrp3 remained inducible by phenobarbital in the CAR-/- mice (610%). Constitutive expression of Mrp3 was not significantly altered in the vehicle-treated CAR-/- mice when compared with the wild-type animals.
Discussion
The current study indicates that induction of hepatic Mrp3 by PB and other prototypical CYP2B inducers is independent of CAR. Our results in three separate models where the presence or activity of CAR is altered indicate that the induction of CYP2B by PB is mediated by CAR, consistent with previously published reports (Wei et al., 2000; Yoshinari et al., 2001). However, unlike CYP2B, Mrp3 remains inducible in each model in which CAR levels or function are defective.
In the WKY rat model, CAR protein expression is much lower in untreated female liver than in untreated male liver, as well as in nuclear extracts of female rats treated with PB (Yoshinari et al., 2001). The result of this sexually dimorphic expression of CAR is putatively the cause for the gender-selective induction of CYP2B1 mRNA in WKY rats, where females are only slightly induced by PB, but male induction is robust. Therefore, each gene that is transcriptionally activated by CAR should be induced more in male than in female WKY rats. Additionally, all microsomal enzyme inducers that activate CAR would result in the gender-selective induction of CYP2B1. The current data suggest that PB, diallyl sulfide, and trans-stilbene oxide induce CYP2B1/2 through the common mechanism of CAR activation. However, oltipraz, which also causes Nrf2-mediated activation of the antioxidant response element (Ramos-Gomez et al., 2001), may induce CYP2B1/2 by a mechanism other than CAR activation.
We have previously demonstrated that hepatic Mrp3 mRNA levels are induced by each of these chemical treatments in male Sprague-Dawley rats (Cherrington et al., 2002), which is mirrored here in male WKY rats. Strikingly, none of the treatments produced a male-selective induction of Mrp3 mRNA levels as for CYP2B1/2, suggesting a mechanism of Mrp3 induction that is independent of CAR. A comparison of the fold induction of Mrp3 in male and female WKY rats is somewhat complicated by the increased basal expression in female rats. However, these results, together with a recent report showing PB induction of Mrp3 mRNA in HepG2 cells with and without stable expression of mouse CAR and PB producing a gender-independent induction of Mrp3 in WKY rats (Xiong et al., 2002), provide strong correlative evidence for a CAR-independent mechanism of Mrp3 induction by PB and PB-like microsomal enzyme inducers.
The second model, male RXRα-/- mice, was used to study the role of CAR activity in the induction of Mrp3. RXR, the obligate binding partner of CAR, is necessary for DNA binding (Baes et al., 1994). There are three separate RXR genes, encoding RXRα, -β, and -γ, that are each capable of heterodimerizing with nuclear orphan receptors (Chiba et al., 1997). Whereas lack of RXRα in a conventional knockout mouse is embryo lethal (Kastner et al., 1994; Sucov et al., 1994), mice lacking both RXRβ and RXRγ and having only one copy of RXRα are viable and are not known to suffer from any impairment of liver function (Krezel et al., 1996). In contrast, the RXRα-/- mice in this study used a cre/lox-mediated recombination to selectively mutate the RXRα gene in adult hepatocytes. These mice have established an integral role for RXRα, in which heterodimerization with several nuclear orphan receptors including CAR, is compromised (Wan et al., 2000). In that study, expression of CYP2B10 mRNA was reduced below 10% of wild-type mice, suggesting that the RXRα-CAR heterodimer is the major activator of at least basal transcription of CYP2B10.
In the present study, Mrp3 induction by each of the microsomal enzyme inducers is independent of the presence or absence of RXRα, thus implying that Mrp3 induction is CAR-independent. However, this does not rule out the possibility that RXRβ or RXRγ heterodimerize with CAR to activate transcription of Mrp3. Alternatively, the lack of induction of CYP2B10 by both oltipraz and diallyl sulfide in RXRα-/- mice suggests either a possible decreased binding affinity of the alternate RXRs with CAR to the PB-responsive element or a separate RXRα-independent mechanism for the induction of CYP2B10 (for a discussion of other mechanisms, see Jarukamjorn et al., 2001). Additionally, we are aware of the potential gender differences involved in the use of male RXRα mice and are pursuing these possible implications.
The CAR-/- model was used to definitively determine whether CAR is necessary for the induction of Mrp3. In these mice, the DNA-binding domain of the CAR gene was replaced by the β-galactosidase gene. These animals are unable to express CAR mRNA or induce CYP2B10 in response to PB (Wei et al., 2000). As expected, PB produced a robust induction of hepatic CYP2B10 in wild-type mice that was completely absent in the CAR-/- mice. Similarly, PB induced Mrp3 in wild-type mice. However, unlike CYP2B10, PB induced Mrp3 in CAR-/- mice, although to a slightly smaller degree. This result clearly shows induction of Mrp3 by PB in the absence of CAR. In contrast, TCPOBOP (a potent CYP2B inducer) was recently shown to induce Mrp3 90% in a CAR-dependent manner (Maglich et al., 2002). It should be noted that TCPOBOP activates CAR-mediated transcription by directly binding to CAR, causing nuclear translocation (Moore et al., 2000; Tzameli et al., 2000), whereas PB does not bind to CAR but is rather a CAR activator, which is putatively mediated by nuclear phosphorylation, resulting in increased CAR activity (Zelko and Negishi, 2000). Thus, whereas both TCPOBOP and PB can activate CAR-mediated transcription and Mrp3 expression, we conclude that PB and other PB-like inducers are capable of inducing Mrp3 in a CAR-independent manner.
The present data comparing vehicle-treated controls of wild-type and knockout strains provide insight into the mechanism(s) of constitutive expression of both CYP2B and Mrp3. The present data confirm previous reports showing lower constitutive CYP2B mRNA levels in RXR-/- mice compared with wild-type mice (Wan et al., 2000; Wei et al., 2000; Yoshinari et al., 2001). Although CAR-/- mice had similar levels of Mrp3 mRNA compared with the wild-type mice, female WKY rats show a significantly higher level of Mrp3 in the vehicle-treated control groups, indicating that CAR may play a role in regulating other physiological factors important in the basal transcription of Mrp3. Interestingly, Mrp3 levels are greatly reduced in the RXRα-/- mice where several signaling pathways are defective, indicating that RXRα, in one of its many roles, is critical for the constitutive expression of hepatic Mrp3. To date, the complete mechanism(s) for constitutive expression of Mrp3 has not been elucidated, although SP1 and SP3 have been implicated in the basal transcription in a rat intestinal cell line (Tzeng and Huang, 2002).
In summary, the present data clearly demonstrate that PB induction of hepatic CYP2B mRNA is CAR-dependent, as expected. CAR is also required for CYP2B induction by diallyl sulfide and trans- stilbene oxide, but not by oltipraz. Additionally, RXRα is required for the maximal induction of CYP2B10 by these inducers. Most importantly, we conclude that hepatic Mrp3 mRNA induction is both CAR- and RXRα-independent by PB and other PB-like microsomal enzyme inducers. The basis for this response remains to be determined.
Footnotes
-
↵1 Abbreviations used are: Mrp3, multidrug resistance protein 3; CAR, constitutive androstane receptor; PB, phenobarbital; RXR, retinoid X receptor; WKY, Wistar Kyoto; DAS, diallyl sulfide; TSO, trans-stilbene oxide; OPZ, oltipraz; EST, expressed sequence tag; TCPOBOP, 1,4-bis[2-(3,5-dichloropyridyloxy)]benzene.
-
This work was supported by National Institutes of Health Grants ES-09716 and ES-03192 (to C.D.K.), CA53596 and AA014147 (to Y.-J.Y.W.) and DK46546 (to D.D.M.).
- Received February 28, 2003.
- Accepted July 24, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}