


Abstract
The concurrent use of herbal medicinals with prescription and over-the-counter drugs carries a risk for unanticipated adverse drug-botanical pharmacokinetic interactions, particularly as a result of cytochrome P450 (P450) inhibition. Extracts of goldenseal (Hydrastis canadensis) containing approximately equal concentrations (∼17 mM) of two methylenedioxyphenyl alkaloids, berberine and hydrastine, inhibited with increasing potency (CYP2C9) diclofenac 4′-hydroxylation, (CYP2D6) bufuralol 1′-hydroxylation, and (CYP3A4) testosterone 6β-hydroxylation activities in human hepatic microsomes. The inhibition of testosterone 6β-hydroxylation activity was noncompetitive with an apparent Ki of 0.11% extract. Of the methylenedioxyphenyl alkaloids, berberine (IC50 = 45 μM) was the more inhibitory toward bufuralol 1′-hydroxylation and hydrastine (IC50 ∼350 μM for both isomers), toward diclofenac 4′-hydroxylation. For testosterone 6β-hydroxylation, berberine was the least inhibitory component (IC50 ∼400 μM). Hydrastine inhibited testosterone 6β-hydroxylation with IC50 values for the (+)- and (-)-isomers of 25 and 30 μM, respectively. For (-)-hydrastine, an apparent Ki value of 18 μM without preincubation and an NADPH-dependent mechanism-based inhibition with a kinactivation of 0.23 min-1 and a KI of ∼110 μM were determined. Cytochrome P450 metabolic-intermediate (MI) complex formation could be demonstrated for both hydrastine isomers. With expressed P450 isoforms, hydrastine formed a P450 MI complex with CYP2C9, CYP2D6, and CYP3A4. Coexpression of cytochrome b5 with the P450 isoforms enhanced the rate but not the extent of P450 MI complex formation.
Phytomedicinals are being used concomitantly with conventional prescription or over-the-counter drugs, a situation that carries the risk of unanticipated adverse drug-botanical pharmacokinetic interactions. A likely important locus of interactions is phase I drug metabolism, where alterations by both inhibition and induction can occur. Goldenseal (Hydrastis canadensis) is a popular immunostimulant, and goldenseal-containing herbal supplements rank high among botanical products sold (Eisenberg at al., 1998). Goldenseal extract contains isoquinoline alkaloids, including berberine, (+)- and (-)-hydrastine, and lesser amounts of hydrastinine (Fig. 1). Chemically, these three goldenseal alkaloids possess a methylenedioxyphenyl (MDP1) moiety which, in studies of cytochrome P450 (P450)-dependent drug metabolism, frequently give rise to inhibition. The inhibition often shows complex inhibition kinetics (Franklin, 1972). The complexity arises from the ability of the methylenic carbon in MDPs to undergo oxidization to a carbene, which then interacts with the heme iron of cytochrome P450 to produce a stable heme-adduct termed a cytochrome P450 metabolic-intermediate (MI) complex (Franklin, 1971; Philpot and Hodgson, 1971). Inhibition by P450 MI complexes is best characterized as “quasi-irreversible” (Ortiz de Montellano and Reich, 1986). The methylenedioxyphenyl P450 MI complexes show pH dependent dual absorbance maxima around 455 and 430 nm (Hodgson et al., 1973) with the former being the more readily identifiable at pH 7.4 (Franklin, 1971). Most investigations with P450 MI complex formation have been undertaken using laboratory animal liver microsomes; little has been undertaken using human enzymes. A recent report (Mathews et al., 2002) has recorded that methysticin and dihydromethysticin, two MDP components of kava extract, are able to form a P450 MI complex in human hepatic microsomes. P450 MI complex formation with goldenseal MDP components has not been reported in either animals or humans.
Chemical structures of methylenedioxyphenyl components of goldenseal extract.
In experiments with 21 commercial ethanolic extracts of botanicals available in Canada, goldenseal rhizome extract was shown to have the highest CYP3A4-inhibitory activity. Goldenseal extract-inhibitory activity was similar to that of St. John's wort extract, and these two were 1 to 2 orders of magnitude more inhibitory than all others (Budzinski et al., 2000). However, there is no information on the complexation of CYP3A4 or other human cytochrome P450 enzymes by goldenseal and its components. This study provides a mechanistic basis for the inhibition of CYP3A4 by goldenseal extract and its MDP components and extends the studies to other cytochromes P450, CYP2C9 and CYP2D6.
Materials and Methods
Chemicals and Reagents. Berberine, (1S,9R)-(+)-β-hydrastine [(+)-hydrastine], (1R,9S)-(-)-β-hydrastine [(-)-hydrastine], diclofenac sodium, mefenamic acid, testosterone, and NADPH were purchased from Sigma-Aldrich (St. Louis, MO). Propranolol hydrochloride was obtained from Wyeth-Ayerst (Princeton, NJ), and 4-hydroxydiclofenac, (±)-bufuralol hydrochloride, and 1′-hydroxybufuralol were obtained from Ultrafine Chemical Company (Manchester, UK). 6β-Hydroxytestosterone and 11β-hydroxytestosterone were obtained from Steraloids (Wilton, NH). In the interest of examining what people consume, three goldenseal root extracts were purchased: Nature's Way (Springville, UT), Wild Oats, (Boulder, CO), and Herb Pharm (Williams, OR). The contents of each were listed as containing 250 mg of dry root extract per ml, 200 mg herb weight per ml, and liquid extract, respectively. Human liver microsome samples selected for high activity toward the P450 isozyme-selective substrates of interest, HBI 102, 217, and 235 for testosterone 6β-hydroxylation, HBI 113, 229, and 233 for bufuralol 1′-hydroxylation, and HBI 101, 102, and 113 for diclofenac 4′-hydroxylation, were purchased from Human Biologics International (Scottsdale, AZ). Donor information and enzyme activity characteristics were obtained from the supplier. The individual microsomal samples used in each experiment are identified in the legends to the figures. Supersomes expressing a single P450 isoform (CYP2C9, CYP2D6, and CYP3A4) together with P450 reductase (and some with cytochrome b5 expressed in addition) were purchased from BD Gentest (Woburn, MA). All solvents were of reagent grade quality or better. HPLC-grade acetonitrile and methanol were purchased from Sigma-Aldrich.
Methylenedioxyphenyl Component Concentrations in Goldenseal Extracts. The contents of the two major MDP components in goldenseal extracts were determined from comparison with calibration curves of the pure alkaloids, berberine and (-)-hydrastine, as previously described (Abourashed and Khan, 2001). The alkaloids were separated by HPLC using CH3CN/0.1% TFA in H2O with UV detection at 300 nm and a flow rate of 1 ml/min on a C18 Supelco Discovery, 5-μm, 250 × 4.6 mm column. The compositions reported are the mean and standard error of three independent determinations of each extract.
Cytochrome P450 Enzyme-Selective Activities. The human microsomal metabolism of diclofenac (4-hydroxylation), bufuralol (1′-hydroxylation), and testosterone (6β-hydroxylation) were examined in the presence of goldenseal extract (Nature's Way) and concentrations of the goldenseal MDPs up to 1 mM. Goldenseal extract was evaluated at final incubation concentrations of 0.1 to 1% (v/v). The pure alkaloids were dissolved in methanol, the final concentration of which was 1% (v/v) in the incubation mixture. The reactions were initiated by the addition of NADPH (3 mM) and incubations were for time periods over which, in the absence of inhibitor, the reaction was linear. The substrates and metabolites were separated by HPLC at a flow rate of 1 ml/min on a C18 Supelco Discovery, 5-μm, 250 × 4.6 mm column. The isozyme-selective oxidative reactions utilized were based on previously described methods (Guengerich et al., 1986; Kronbach et al., 1987; Leemann et al., 1993). For CYP2C9 activity, human microsomes (HBI 101, 102, and 113) at 0.5 mg protein/ml were incubated with 100 μM diclofenac in Tris-chloride buffer (pH 7.6), for 30 min at 37°C. Diclofenac and its metabolite were extracted three times with ethyl acetate and separated by HPLC with CH3CN/0.1% TFA in H2O with UV detection at 268 nm. Mefenamic acid, added at the termination of the incubation, was used as an internal standard. For CYP2D6 activity, human liver microsomes (HBI 113, 229, and 233) at 1.0 mg protein/ml were incubated with 25 μM (±)-bufuralol in phosphate buffer (pH 7.6), for 30 min at 37°C. The reaction was terminated by the addition of 70% perchloric acid. The reaction mixture was centrifuged and the supernatant injected directly into the HPLC apparatus for analysis. The substrate and metabolites were separated using CH3CN/2 mM perchloric acid with fluorescence detection (excitation 285 nm, emission 310 nm). Propranolol, added at the termination of the incubation, was used as an internal standard. For CYP3A4 activity, human liver microsomes (HBI 102, 217, and 235) at 0.2 mg protein/ml were incubated with 200 μM testosterone in phosphate buffer (pH 7.4), for 15 min at 37°C. Testosterone and its 6β-hydroxylated metabolite were extracted with CH2Cl2 and separated by HPLC using CH3CN/0.1% TFA in H2O with UV detection at 236 nm. The internal standard, 11β-hydroxytestosterone, was added at the termination of the incubation. Data are reported as mean (± standard error) of the different microsomal samples utilized.
NADPH-Dependent Mechanism-Based Inactivation of Testosterone 6β-Hydroxylase (CYP3A4) Activity. To determine the NADPH-dependent mechanism-based inactivation of testosterone 6β-hydroxylase activity, concentrations of (-)-hydrastine (up to 300 μM) were preincubated for specific time intervals (0-3 min) with human liver microsomes (HBI 102 at 2.0 mg protein/ml) in the presence of 3 mM NADPH. After preincubation, an aliquot of the preincubation reaction mixture was diluted 10-fold into a secondary reaction mixture containing testosterone (200 μM) and 3 mM NADPH. The secondary (reporter) reaction was allowed to proceed for an additional 15 min and assayed for testosterone 6β-hydroxylase activity as previously described.
Cytochrome P450 Metabolic-Intermediate Complex Formation. The formation of P450 MI complexes was determined according to the method of Franklin (1981). Human liver microsomes (HBI 102 and 217 at 0.167 μM P450) were incubated with hydrastine or hydrastinine (up to 333 μM) in a masked microcuvette in an Aminco DW-2000 spectrophotometer, under aerobic conditions at 25°C in Tris-chloride buffer (pH 7.4). To determine the individual P450s responsible for the P450 MI complex formation, Supersomes [CYP2C9 (+b5 and -b5), CYP2D6, and CYP3A4 (+b5 and -b5)] at 0.167 μM P450 were incubated with MDP compounds (up to 333 μM). Interference in the Soret region thwarted determination of MI complex formation from berberine and whole goldenseal extract. For P450 MI complex identification, the formation was initiated by the addition of 1 mM NADPH, and the absorbance changes were monitored in split beam mode, by scanning at 1-min intervals. For determination of the initial rate (absorbance change in 0-30 s) and extent of P450 MI complex formation, absorbance changes at 455 nm relative to 490 nm were monitored in dual wavelength mode. After 3 min of equilibration, 1 mM NADPH was added to initiate the reaction and complex formation was continuously recorded. P450 MI complex formation is reported in Δ absorbance units since, in contrast to amine-derived P450 MI complexes, no reliable extinction coefficient has been established for human P450s and goldenseal MDPs (see Discussion).
Results
Content of Methylenedioxyphenyl Components in Goldenseal Extract. The amount of the two major MDP-containing alkaloids, berberine and hydrastine, in three commercial samples of goldenseal extract was determined by HPLC analysis (Table 1). All three samples contained berberine and hydrastine in the ratio of approximately 1 to 1.3, suggesting authentic plant sources, and at concentrations of 17.1 to 19.4 mM for berberine and 13.4 to 17.4 mM for hydrastine.
Content of methylenedioxyphenyl components in three commercially available goldenseal preparations
Effect of Goldenseal Extract on CYP2C9, CYP2D6, and CYP3A4 Activities. The ability of goldenseal extract to alter three microsomal P450-marker enzymatic activities, diclofenac 4-hydroxylation (CYP2C9), bufuralol 1′-hydroxylation (CYP2D6), and testosterone 6β-hydroxylation (CYP3A4) is shown in Fig. 2. Goldenseal extract inhibited diclofenac 4-hydroxylation, bufuralol 1′-hydroxylation, and testosterone 6β-hydroxylation activity with interpolated IC50 values of 0.98%, 0.66%, and 0.18%, respectively. Closer examination of testosterone 6β-hydroxylation, the activity most susceptible to inhibition, indicated that goldenseal extract inhibition was noncompetitive (Fig. 3A), with an apparent mean Ki of 0.11% (v/v) for the two human liver microsomal samples examined (Fig. 3B).

The effect of goldenseal extract on human hepatic microsomal CYP2C9, CYP2D6, and CYP3A4-dependent activities.
The effect of goldenseal extract (Nature's Way) on CYP2C9 (diclofenac 4′-hydroxylation), CYP2D6 (bufuralol 1′-hydroxylation), and CYP3A4 (testosterone 6β-hydroxylation) activities are shown. Each data point is the mean and standard error from three individual microsomal samples (HBI 102, 217, and 235 for testosterone 6β-hydroxylation; HBI 113, 229, and 233 for bufuralol-1′-hydroxylation; and HBI 101, 102, and 113 for diclofenac 4′-hydroxylation).
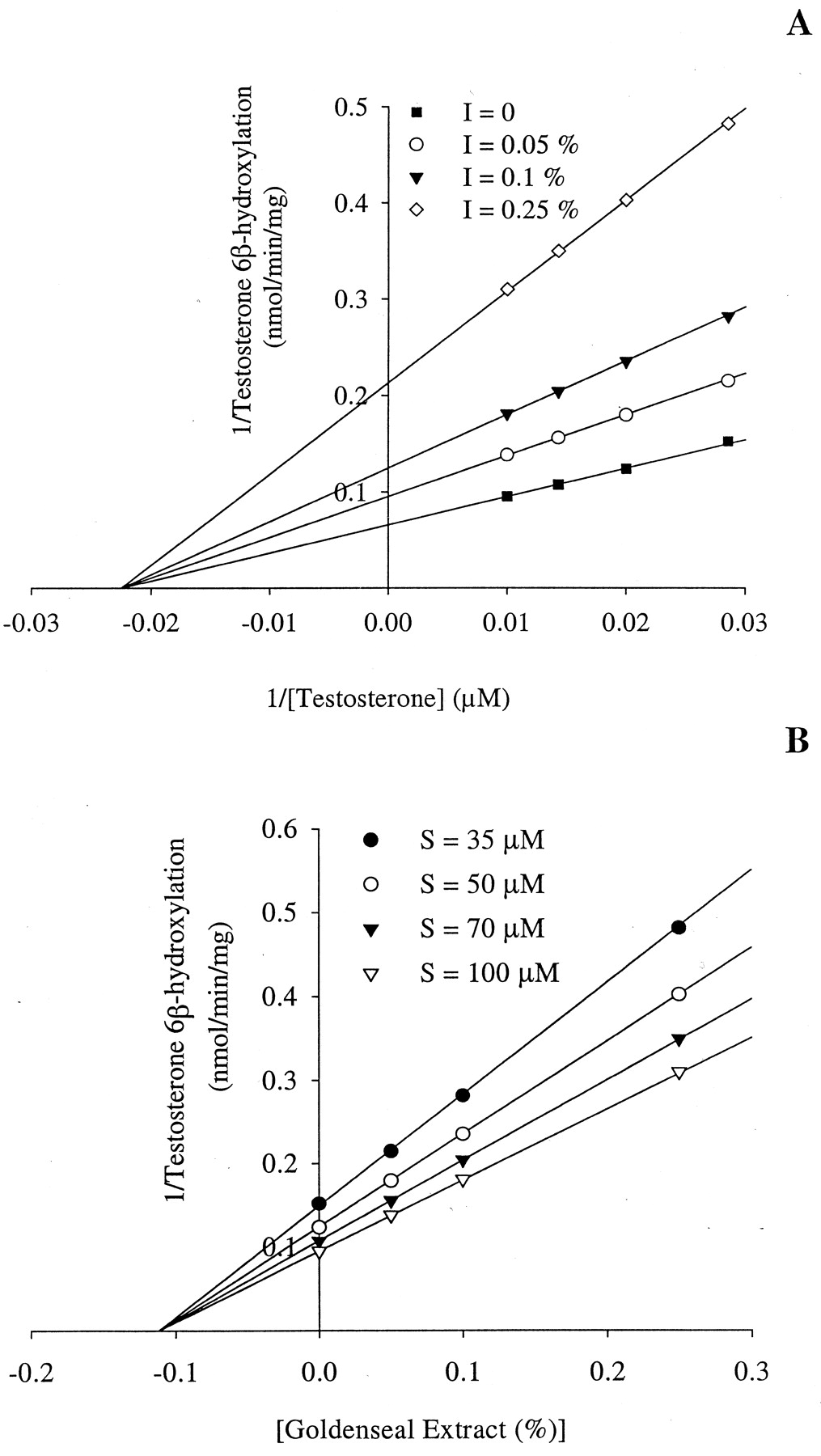
Enzyme kinetics plots of the inhibition of human hepatic microsomal testosterone 6β-hydroxylation activity by goldenseal extract.
Three goldenseal extract (inhibitor) (0.05-0.25%) and four testosterone (substrate) (35-100 μM) concentrations were examined. Data are presented as Lineweaver-Burk (A) and Dixon (B) plots. Each data point is the average from two human liver microsomal samples (HBI 217 and 235).
Effect of Individual Goldenseal Extract MDP Components on CYP2C9, CYP2D6, and CYP3A4 Activities. The relative abilities of two major goldenseal MDP components, berberine and hydrastine, to alter the three microsomal P450-marker enzymatic activities are shown in Fig. 4. Both isomers of hydrastine were evaluated because the isomeric composition of the extract was not known. Berberine inhibited testosterone 6β-hydroxylation with an interpolated IC50 value of 400 μM, whereas the (+)- and (-)-isomers of hydrastine were much more inhibitory, with interpolated IC50 values of 25 and 30 μM, respectively (Fig. 4A). Berberine also inhibited bufuralol 1′-hydroxylation (with an interpolated IC50 value of 45 μM; Fig. 4B) but was less inhibitory toward diclofenac 4-hydroxylation (Fig. 4C). Both isomers of hydrastine inhibited diclofenac 4-hydroxylation and bufuralol 1′-hydroxylation with interpolated IC50 values of 350 and 380 μM, and 500 and 1,000 μM, for the (+)- and (-)-isomers, respectively (Fig. 4, A and B). The hydrastine inhibition of testosterone 6β-hydroxylation activity was noncompetitive (Fig. 5A), for which an apparent Ki of 18 μM could be determined (Fig. 5B). The ability of hydrastine to inhibit testosterone 6β-hydroxylation activity in a mechanism-based manner was also assessed (Fig. 6). There was a rapid NADPH-dependent inactivation, seemingly complete within 1 min (Fig. 6A), for which a kinactivation of 0.23 min-1 could be determined (Fig. 6B). The KI for this mechanism-based inactivation was 110 μM.

The effect of methylenedioxyphenyl components on human hepatic microsomal CYP2C9-, CYP2D6-, and CYP3A4-dependent activities.
The effects of berberine and (+)- and (-)-hydrastine on CYP3A4 (testosterone 6β-hydroxylation) (panel A), CYP2D6 (bufuralol 1′-hydroxylation) (panel B), and CYP2C9 (diclofenac 4′-hydroxylation) (panel C) activities are shown. MDP concentrations used were 10, 100, and 500 μM for berberine and 10, 100, and 1,000 μM for the hydrastine isomers. Each data point is the mean and standard error from three individual microsomal samples (HBI 102, 217, and 235 for testosterone 6β-hydroxylation; HBI 113, 229, and 233 for bufuralol 1′-hydroxylation; and HBI 101, 102, and 113 for diclofenac 4′-hydroxylation).

Enzyme kinetics plots of the inhibition of human hepatic microsomal testosterone 6β-hydroxylation activity by (-)-hydrastine.
Three (-)-hydrastine (inhibitor) (1-100 μM) and five testosterone (substrate) (35-200 μM) concentrations were used. Data are presented as Lineweaver-Burk (A) and Dixon (B) plots. Each data point is the average from two human liver microsomal samples (HBI 217 and 235).

Time- and NADPH-dependent inhibition of human hepatic microsomal testosterone 6β-hydroxylation activity by (-)-hydrastine.
A, the (-)-hydrastine (inhibitor) concentrations indicated are those employed in the preincubation. Each data point is the average from duplicate incubations using microsomal sample HBI 102. The 0- to 1-min rate was used to derive the plot (B).
P450 MI Complex Formation by Goldenseal MDP Compounds. With both goldenseal extract and berberine, interference in the Soret region precluded detection of any P450 MI complex formation. Aerobic incubation of human liver microsomes with NADPH and hydrastinine, a minor goldenseal MDP component, showed no P450 MI complex formation within the concentration range examined (33-333 μM). Incubation with either (+)-hydrastine or (-)-hydrastine revealed P450 MI complex formation with a peak absorbance at 455 nm. The amount of P450 MI complex increased rapidly with time and reached a maximum by 4 min (Fig. 7A). P450 MI complex formation was concentration-dependent (Fig. 7B). Supplementation of 33 and 100 μM incubations with hydrastine and NADPH after a 5-min interval increased the extent of P450 MI complex formation, but only up to a maximum equal to that seen with 333 μM (absorbance traces not shown). The apparent Km values for the initial rate of P450 MI complex formation (Δabsorbance in the first 30 s) for (+)- and (-)-hydrastine were similar, at 115 and 105 μM, respectively. The addition of P450-selective substrates, testosterone (Fig. 7A) and diclofenac (not shown), subsequent to P450 MI complex formation did not dissociate the complex.

P450 Metabolic-intermediate complex formation by (-)-hydrastine in human liver microsomes.
A, microsomes (HBI 102 at 0.11 mg protein/ml) incubated with 333 μM (-)-hydrastine were scanned repetitively from 405 to 475 nm at 1-min intervals for 5 min after the addition of 1 mM NADPH. After 5 min, testosterone (333 μM) was added and the cuvette contents were rescanned. B, absorbance changes at 455 nm relative to 490 nm were continuously monitored (dual wavelength mode) in microsomes (HBI 102 at 0.11 mg protein/ml; 0.167 μM P450) containing 33, 100, or 333 μM (-)-hydrastine after the addition of 1 mM NADPH.
To delineate the individual P450s within human hepatic microsomes that might be responsible for the P450 MI complex formation seen with hydrastine, heterologously expressed P450 isozymes, CYP2C9 (+b5 and -b5), CYP2D6, and CYP3A4 (+b5 and -b5), were incubated with (+)- and (-)-hydrastine (Table 2). CYP3A4 Supersomes exhibited the highest rate of P450 MI complex formation. The rate of P450 MI complex formation was significantly enhanced in both of the Supersome preparations in which the P450s had been coexpressed with cytochrome b5 (Table 2). The enhancement was similar for both isoforms, 2.6- to 4-fold for CYP3A4, and 2.4- to 3-fold for CYP2C9.
Initial rate of cytochrome P450 metabolic-intermediate complex formation with (+)- and (−)-hydrastine (333 μM) in Supersomes containing CYP2C9, CYP2D6, and CYP3A4
Discussion
Goldenseal preparations are often used for the relief of cold and influenza symptoms and, because of their antibacterial properties (Abdel-Haq et al., 2000), in the treatment of dermatological, ocular, and urinary tract infections (Müller et al., 1995). In clinical trials, goldenseal extract was found to ameliorate the primary IgM response in test subjects (Rehman et al., 1999). Such studies prompted wide acceptance of goldenseal extract as an alternative influenza remedy. The human recommended daily dose of goldenseal extract is 10 to 30 drops, two to four times per day. Based on our analysis of three commercially available goldenseal preparations, all of which were very similar in composition, this equates to 15 to 30 mg of each alkaloid per day. Although whole-body distribution of this amount would produce very low concentrations, first-pass access to the liver after oral ingestion and the quasi-irreversible formation of inactive cytochrome P450 complexes during that first pass have the potential to affect the metabolism of other drugs. About 80% of the prescription and over-the-counter drugs currently available are metabolized, at least in part, by CYP2C9, CYP2D6, or CYP3A4. Of these, CYP3A4 (testosterone 6β-hydroxylation) proved the most susceptible to inhibition by goldenseal extract (Fig. 2). The Ki for this reaction was 0.11% extract (Fig. 3B). In terms of hydrastine and berberine concentrations (from Table 1), this would equate to 19 and 21 μM, respectively. Because the hydrastine isomer composition of the extract was not known, and the two isomers could be obtained in purified form, both isomers were evaluated for inhibitory effects but were found to differ little, with IC50 values of 25 and 30 μM (Fig. 4A). This indicates that any variations in the isomeric proportion in extracts would not markedly influence the degree of inhibition. The Ki for CYP3A4 inhibition by hydrastine [(-)-isomer] was 18 μM (Fig. 5A), suggesting that most of the CYP3A4 inhibition by extract could be due to the hydrastine it contains. Berberine, with an IC50 of ∼400 μM (Fig. 4A), would not be a major contributor to the observed CYP3A4 inhibition by extract.
The inhibitory effects of major MDP components of extract were reversed for CYP2D6 (bufuralol-1′-hydroxylation). Berberine, with an IC50 of 45 μM, was more inhibitory than the hydrastine isomers (IC50 values >500 μM) and would contribute more to the CYP2D6 inhibition seen with extract. However, 45 μM berberine would be present at 0.23% extract, a concentration at which inhibition was less than 50%, indicating that one or more components in the extract were in some manner preventing berberine from exerting its full inhibitory effect. For CYP2C9 (diclofenac 4-hydroxylation) inhibition, the extract IC50 value was 0.98%, which would represent 167 μM hydrastine and 190 μM berberine. With IC50 values of around 350 μM for the hydrastine isomers and around 500 μM for berberine, the extract inhibition could contain additive contributions of the two components but would not exclude contributions from other even more inhibitory components in the extract.
The noncompetitive nature of the inhibition of CYP3A4 metabolism by the extract, the most sensitive of the isozymes investigated, and a similar mechanism of inhibition by hydrastine, the more inhibitory of the two MDP components investigated, raised the possibility that inhibition via P450 MI complex formation may be occurring. Indeed, P450 MI complex formation from hydrastine could be detected in microsomes (Fig. 7) and with all three of the heterologously expressed enzymes examined (Table 2). A rapid formation of a P450 MI complex (complete within minutes) (Fig. 7) corresponds to the short time period over which an NADPH-dependent mechanism-based inactivation is seen (Fig. 6). Using the commonly adopted extinction coefficient of 75 mM-1 cm-1 for the complex, the maximum 455-nm absorbance seen for hydrastine (Fig. 7B) would account for 54% of the P450 present in the human hepatic microsomes, but the validity of this extinction coefficient for hydrastine and all P450 isoforms is open to question. Using displacement techniques, the extinction coefficient for the 455-nm absorbance of the isosafrole P450 MI complex in rats was 75 mM-1 cm-1 (Elcombe et al., 1976) and 73 mM-1cm-1 (Fennell and Bridges, 1979) in isosafrole-induced and 80 mM-1cm-1 in 3-methylcholanthrene-induced animals (Fennell et al., 1979), but only 38 mM-1 cm-1 in those induced with phenobarbital (Fennell et al., 1979). The extinction coefficients for two other MDPs, were 30 mM-1 cm-1 for safrole in safrole-induced animals and 33 mM-1 cm-1 for dihydrosafrole in dihydrosafrole-induced animals (Fennell and Bridges, 1979). Thorough studies to understand the reasons for these differences need to be undertaken before a more detailed balance sheet of human isoform contributions to observed absorbance changes at 455 nm can be meaningfully undertaken.
Without any preincubation, the hydrastine inhibition of testosterone 6β-hydroxylation activity was noncompetitive, with an apparent Ki of 18 μM (Fig. 5B). With preincubation with NADPH, there was a rapid mechanism-based inactivation (Fig. 6A) for which a kinactivation of 0.23 min-1 could be determined (Fig. 6B). As indicated above, this rapid inactivation is compatible with the rapid formation of a P450 MI complex. The KI for inactivation was 110 μM, a value very similar to the Km determined for P450 MI complex formation directly. The substrate concentration-dependent decrease in activity at zero time seen in Fig. 6B is interpreted as being due to the carryover of substrate into the secondary reporter reaction (where concentrations would range from 7.5 to 30 μM) to support MI complex formation during the reporter reaction (testosterone 6β-hydroxylation).
Overall, it is evident that at least three human isoforms are capable of forming MI complexes from hydrastine (Table 2), and the inhibition would be of the quasi-irreversible nature associated with P450 MI complex formation. For one isoform studied in depth (CYP3A4), the characteristics of the observed NADPH-dependent mechanism-based inhibition correlated well with the observed MI complex formation. Such observations support the concept that ingestion of goldenseal extract could potentially inhibit the metabolism of many drugs currently in therapeutic use.
Footnotes
-
↵1 Abbreviations used are: MDP, methylenedioxyphenyl; MI, metabolic-intermediate; P450, cytochrome P450; HPLC, high-performance liquid chromatography; TFA, trifluoroacetic acid.
- Received April 3, 2003.
- Accepted August 18, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}