


Abstract
Concentrative nucleoside transporters (Cnts) and equilibrative nucleoside transporters (Ents) have essential physiological functions and are important in disposition of anticancer and antiviral nucleoside analogs. Information on tissue distribution of Cnts and Ents in rodents is sparse. Thus, the present study aimed to determine the distribution of Cnt1-3 and Ent1-3 transcripts in 19 tissues of Sprague-Dawley rats and C57BL/6 mice of both genders. These six transcripts were quantified using the branched DNA signal amplification assay. Cnt1 transcripts were highest in small intestine, followed by kidney and testes, with similar expression in both species. Cnt2 mRNA was expressed highest in the small intestine of both rats and mice, intermediate in liver of rats but not in mice, and lower in thymus and spleen of both species. Cnt3 mRNA has marked species differences, with the highest expression in lung of rats but uterus of mice. Ent1 mRNA was most highly expressed in testes and lung of both species. Ent1 mRNA was highly expressed in liver and pituitary of mice, but not in rats. Ent2 mRNA was highly expressed in testes and brain of both species. Ent3 mRNA was highest in kidney, followed by testes, in both species. Significant gender differences were observed in kidney (mouse) and heart (rat). These studies demonstrate that in general, tissue distribution of Cnt and Ent is similar in rats and mice. However, a few important species and gender differences do exist, which could be responsible for related differences in efficacy and toxicity of substrates for these transporters.
Nucleoside transporters facilitate cellular uptake/efflux of various nucleosides and nucleobases, which are used for biosynthesis of nucleotides via the salvage pathway (Kinsella et al., 1997; Baldwin et al., 1999). Nucleotides are intermediates of many essential biosynthetic pathways, and are important in neurotransmission and as signaling molecules (Baldwin et al., 1999; Merighi et al., 2003). For example, adenosine is an important signaling molecule in brain, lung, kidney, heart, and immune systems (Baldwin et al., 1999; Fishman et al., 2002; Sitkovsky, 2003). Nucleoside transporters play an important role in terminating adenosine signaling by removing it from extracellular space, limiting its availability to cell-surface adenosine receptors.
Concentrative nucleoside transporters (CNTs) and equilibrative nucleoside transporters (ENTs) transport endogenous substrates such as adenosine, thymidine, cytidine, guanosine, uridine, inosine, and hypoxanthine (Ward et al., 2000; Ritzel et al., 2001a). CNTs facilitate Na+-dependent uptake of nucleosides into cells against a concentration gradient. ENTs transport nucleosides across plasma membrane bidirectionally in a Na+-independent manner, and are sensitive to certain inhibitors (Yao et al., 1997; Kiss et al., 2000; Ward et al., 2000). Various CNTs and ENTs have different, but overlapping substrate specificity. CNTs have high affinity for their natural substrates. CNT1 preferentially transports pyrimidine nucleosides, CNT2 preferentially transports purine nucleosides, and CNT3 transports both pyrimidine and purine nucleosides (Gray et al., 2004). ENT1 and 2 also transport both pyrimidine and purine nucleosides, but with lower affinity (Yao et al., 1997; Kiss et al., 2000; Ward et al., 2000). Among various CNTs and ENTs, ENT2 is unique in its high capacity for transporting nucleobases such as adenine (Yao et al., 2002).
Many anticancer and antiviral drugs are nucleoside or nucleobase analogs. Most nucleoside analogs are hydrophilic and cannot freely cross the plasma membrane. Pharmaceutically important drugs 5-fluorouridine, 5-fluoro-2′-deoxyuridine, arabinosylcytosine, zidovudine, cladribine, diethyldithiocarbamate, 2′,3′-dideoxyinosine, fludarabine, gemcitabine, melarsoprol, pentamidine, zalcitabine, and zebularine are substrates for nucleoside transporters (Hyde et al., 2001; Ritzel et al., 2001a; Damaraju et al., 2003; Gray et al., 2004). Expression and activity of nucleoside transporters is an important determinant in disposition, efficacy, and toxicity of these nucleoside analogs in vivo (Damaraju et al., 2003).
CNT1 is localized primarily in epithelia, such as intestine and kidney, whereas CNT2 and 3 have more broad distributions (Gray et al., 2004). ENT1, 2, and 3 are all widely distributed in mammalian tissues, and ENT2 is particularly abundant in human skeletal muscle (Baldwin et al., 2004). CNTs are located in the apical membrane, whereas ENTs are detected in the basolateral membrane of epithelial cells, suggesting a coordinate role of CNTs and ENTs in transepithelial nucleoside transport (Casado et al., 2002; Baldwin et al., 2004).
Despite extensive functional characterization of human and rodent nucleoside transporters (for review, see Gray et al., 2004; Baldwin et al., 2004), most knowledge on the tissue distribution of nucleoside transporters is obtained from humans, with little known about rats, and even less known about mice. Moreover, there are no published studies concerning gender-related differences in tissue distribution of nucleoside transporters, although gender differences in the disposition and toxicity of nucleoside analogs have been reported in humans (Burger et al., 1994; Pai and Nahata, 2000; Ofotokun and Pomeroy, 2003). In drug development, pharmacokinetics/toxicology (and frequently efficacy) studies in rats and mice are required before conducting clinical studies in humans. However, only ∼36% of the site-specific carcinogenic effects observed in one species (rats or mice) were also detected in the other species (Haseman and Lockhart, 1993). Many drug candidates fail in clinical trials due to undesirable absorption, distribution, metabolism, and excretion properties and toxicity (Lin et al., 2003). In the development of anticancer and antiviral nucleoside and nucleobase drugs, it is very important to know not only the similarities, but also the differences in tissue distribution of these nucleoside transporters among humans, rats, and mice, as well as the differences between males and females, to correctly allow for extrapolation of animal data to humans.
The purpose of this study was to assemble comprehensive information on tissue distribution patterns of Cnt1, 2, and 3 and Ent1, 2, and 3 mRNA in rats and mice of both genders. The mRNA expression of these six nucleoside transporters in 19 tissues of Sprague-Dawley rats and C57BL/6 mice was determined using the branched DNA (bDNA) signal amplification assay. Our data showed that in general, tissue distribution of Cnts and Ents is similar in rats and mice; however, a few important differences do exist. The similarities and differences among humans, rats, and mice, and the potential physiological importance of these observations are discussed.
Materials and Methods
Animals. Male and female Sprague-Dawley rats (8 weeks old) and C57BL/6 mice (8 weeks old) were purchased from Charles River Laboratories, Inc. (Wilmington, MA). Animals were acclimated to an Association for Assessment and Accreditation of Laboratory Animal Care-certified housing facility (70 ± 2°F, 50% relative humidity, 12-h light/dark cycle) for 1 week. Rats and mice were fed Teklad 8604 rodent chow (Harlan Teklad, Madison, WI) and laboratory mouse chow (Purina, St. Louis, MO), respectively, ad libitum. Animals were euthanized in a CO2 atmosphere. Tissues (heart, thymus, spleen, brain, pituitary, muscle, kidney, liver, blood vessel, duodenum, jejunum, ileum, large intestine, stomach, lung, testes, prostate, ovary, and uterus) were promptly removed, washed with ice-cold saline, and snap frozen in liquid nitrogen (intestinal epithelia from rats was obtained by scraping before freezing) and stored at -80°C.
RNA Extraction. Total tissue RNA was extracted using RNA-Bee reagent (Tel-Test Inc., Friendswood, TX) according to the manufacturer's protocol. For some minor tissues, such as ovary and pituitary, tissue samples from 10 animals were pooled to prepare total RNA. The RNA pellet was redissolved in 0.1 to 0.2 ml of diethyl pyrocarbonate-treated water. RNA concentrations were quantified by ultraviolet absorbance at 260 nm. RNA integrity was confirmed by agarose gel electrophoresis of 5 μg of total RNA and visualization of the intact 18S and 28S bands by ethidium bromide staining. The integrity of RNA was further confirmed by quantification of glyceraldehyde-3-phosphate dehydrogenase in these tissues by bDNA signal amplification assay (data not shown).
bDNA Signal Amplification Assay. Cnt1, 2, and 3 and Ent1, 2, and 3 mRNA was quantified using the bDNA assay (QuantiGene bDNA signal amplification kit; Genospectra, Fremont, CA) with modifications (Leazer and Klaassen, 2003). Sequences of target genes were accessed from GenBank. Multiple oligonucleotide probe sets (containing capture, label, and blocker probes) specific to a single mRNA transcript were designed using Probe Designer software v1.0 (Bayer Corp.-Diagnostics Div., Tarrytown, NY) with a mean temperature of approximately 63°C, enabling constant hybridization conditions (i.e., 53°C). Each probe developed in Probe Designer was submitted to the National Center for Biotechnology Information (Bethesda, MD) for nucleotide comparison by the basic local alignment search tool (BLASTn) to ensure minimal cross-reactivity with other known rat/mouse sequences. Oligonucleotides with a high degree of similarity (>80%) to other rat or mouse gene transcripts were eliminated from the probe set design. The design of probe sets for rat Cnt1 and 2 and Ent1 and 2 was reported previously (Leazer and Klaassen, 2003). The GenBank accession number, nucleotide sequence, and function for probes of mouse Cnt1-3, Ent1-3, and rat Cnt3 and Ent3 are given in Table 1. All reagents for analysis were supplied in the QuantiGene bDNA signal amplification kit. Total RNA (10 μg/well) was added to each well of a 96-well plate containing capture hybridization buffer (50 μl) and each diluted probe set (50 μl), and allowed to hybridize to the probe set overnight at 53°C. Subsequent washing and hybridization steps were carried out following the manufacturer's protocol. Luminescence of the 96-well plate was measured with a Quantiplex 320 bDNA luminometer (Bayer Corp.-Diagnostics Div.) interfaced with Quantiplex Data Management software version 5.02 for data analysis. The luminescence for each well is reported as relative light units (RLU) per 10 μg of total RNA. For each probe set, a standard curve was plotted using various amounts (2.5-20 μg) of total RNA from a tissue with moderate or high expression of the target gene; the linearity of the standard curve (R2) ranged from 0.98 to 0.99 (data not shown).
Oligonucleotide probes for analysis of rat and mouse Cnt and Ent Messenger RNA by bDNA signal amplification assay
Statistics. Data are presented as mean ± standard error of five animal tissues except the data on pituitary (rat and mouse) and ovary (mouse), which represent the mean of duplicate pooled samples from 10 animals. Differences between different groups were determined using Student's t test. Asterisks represent a statistical difference (p < 0.05) between genders.
Results
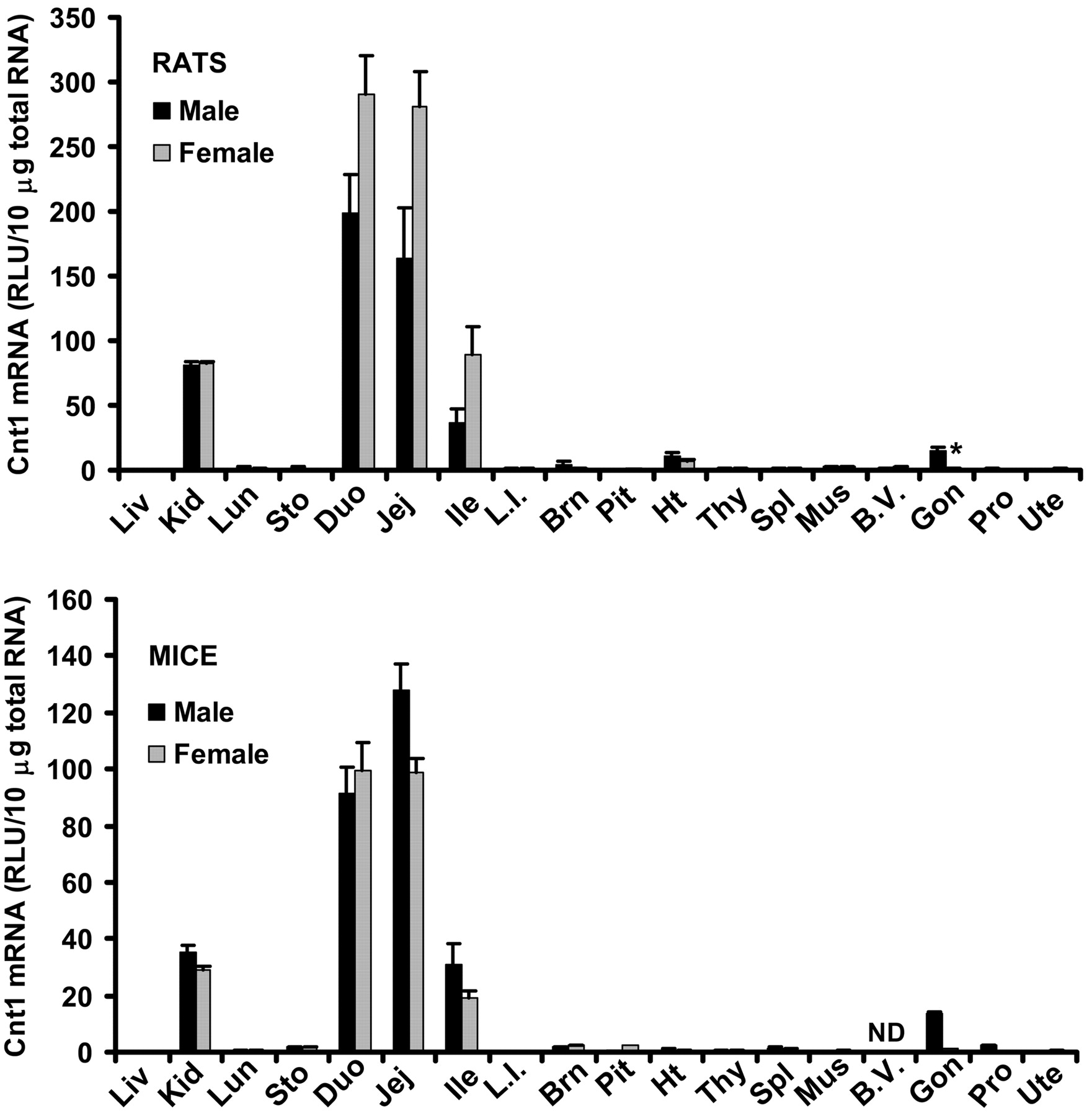
Tissue Distribution of Cnt1 mRNA in Male and Female Rats and Mice. Tissue distribution patterns of Cnt1 mRNA are very similar between rats and mice (Fig. 1). Cnt1 mRNA is highly expressed in the three sections of small intestine, with duodenum and jejunum being the highest, and ileum much lower. Moderate levels of Cnt1 mRNA were also detected in kidney and testes of both species. Low expression of Cnt1 mRNA was found in brain of both species, heart of rats, and stomach and prostate of mice, as well as female pituitary of mice. Cnt1 mRNA levels were very low in other tissues examined. Cnt1 mRNA in testes of rats and mice was 15- and 9-fold higher than that in ovary of rats and mice, respectively.

Tissue distribution of Cnt1 mRNA in male and female rats and mice. Ten micrograms of total RNA from various tissues of male and female Sprague-Dawley rats or C57BL/6 mice were analyzed with the Cnt1-specific probe set described under Materials and Methods. The y-axis represents the RLU per 10 μg of total tissue RNA. Data represent mean ± standard error of determinations from five samples per group, except the data on pituitary (rats and mice) and ovary (mice), which are the mean of duplicate pooled pituitary or ovary samples from 10 rats or mice. Liv, liver; Kid, kidney; Lun, lung; Sto, stomach; Duo, duodenum; Jej, jejunum; Ile, ileum; L.I., large intestine; Brn, brain; Pit, pituitary; Ht, heart; Thy, thymus; Spl, spleen; Mus, skeletal muscle; B.V., blood vessels; Gon, gonads; Pro, prostate; Ute, uterus. ND, not determined. ★, p < 0.05 compared with males.
Tissue Distribution of Cnt2 mRNA in Male and Female Rats and Mice. Tissue distribution patterns of Cnt2 mRNA are also similar between rats and mice (Fig. 2), with one exception, that Cnt2 mRNA was highly expressed in liver of rats but very low in liver of mice. In contrast to Cnt1 mRNA, which was expressed in only a few tissues, Cnt2 mRNA was expressed in many tissues, with the highest expression also in the small intestine, followed by spleen, thymus, heart, and lung of both species. Moderate to low expression of Cnt2 mRNA was found in all the other tissues examined. Mice had similar levels of Cnt2 mRNA in the three sections of small intestine, but rats had higher Cnt2 mRNA expression in duodenum than in jejunum and ileum. Female rats had ∼60% higher Cnt2 mRNA in spleen and blood vessels than did males. Rat and mouse testes had 74% and 3.1-fold higher Cnt2 mRNA than did rat and mouse ovary, respectively.

Tissue distribution of Cnt2 mRNA in male and female rats and mice. Ten micrograms of total RNA from various tissues of male and female Sprague-Dawley rats or C57BL/6 mice were analyzed with the Cnt2-specific probe set described under Materials and Methods. The y-axis represents the RLU per 10 μg of total tissue RNA. Data represent mean ± standard error of determinations from five samples per group, except the data on pituitary (rats and mice) and ovary (mice), which are the mean of duplicate pooled pituitary or ovary samples from 10 rats or mice. Liv, liver; Kid, kidney; Lun, lung; Sto, stomach; Duo, duodenum; Jej, jejunum; Ile, ileum; L.I., large intestine; Brn, brain; Pit, pituitary; Ht, heart; Thy, thymus; Spl, spleen; Mus, skeletal muscle; B.V., blood vessels; Gon, gonads; Pro, prostate; Ute, uterus. ND, not determined. ★, p < 0.05 compared with males.
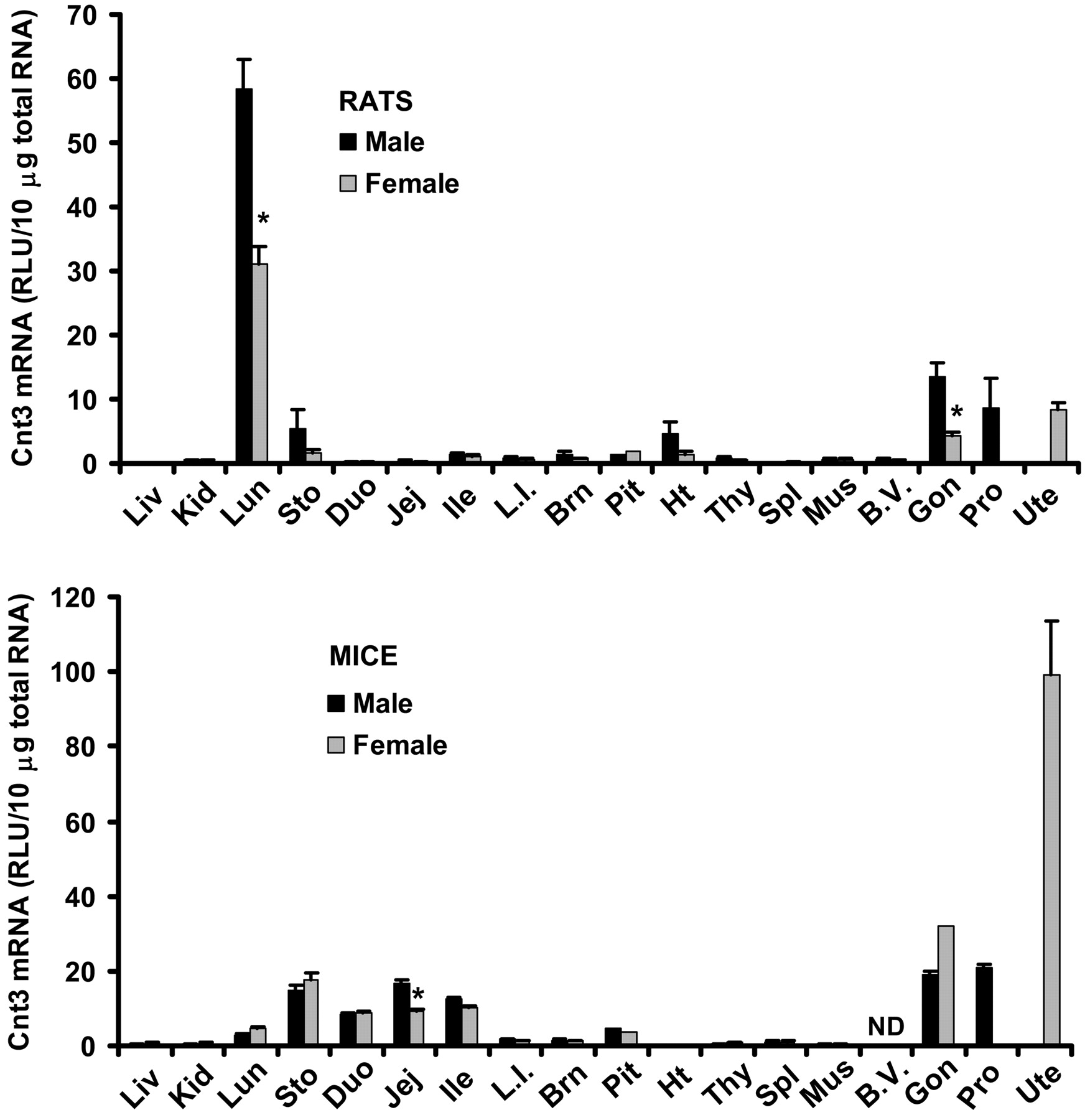
Tissue Distribution of Cnt3 mRNA in Male and Female Rats and Mice. Tissue distribution patterns of Cnt3 mRNA are markedly different between rats and mice (Fig. 3). In rats, Cnt3 mRNA was highest in lung with expression in males 87% higher than in females. In mice, Cnt3 mRNA was highest in uterus but low in lung. Moderate expression of Cnt3 was found in testes, prostate, ovary, brain, pituitary, and stomach of both species. In mice, Cnt3 mRNA was also moderately expressed in the three sections of small intestine, with a higher expression in male jejunum, but Cnt3 mRNA was hardly detectable in duodenum and jejunum of rats. Cnt3 mRNA was moderately expressed in heart of rats but not detectable in heart of mice. Cnt3 mRNA in all the other tissues examined was very low. Rats had 2.3-fold higher Cnt3 mRNA in testes than in ovary, but mice tended to have lower Cnt3 mRNA in testes than in ovary.

Tissue distribution of Cnt3 mRNA in male and female rats and mice. Ten micrograms of total RNA from various tissues of male and female Sprague-Dawley rats or C57BL/6 mice were analyzed with the Cnt3-specific probe set described under Materials and Methods. The y-axis represents the RLU per 10 μg of total tissue RNA. Data represent mean ± standard error of determinations from five samples per group, except the data on pituitary (rats and mice) and ovary (mice), which are the mean of duplicate pooled pituitary or ovary samples from 10 rats or mice. Liv, liver; Kid, kidney; Lun, lung; Sto, stomach; Duo, duodenum; Jej, jejunum; Ile, ileum; L.I., large intestine; Brn, brain; Pit, pituitary; Ht, heart; Thy, thymus; Spl, spleen; Mus, skeletal muscle; B.V., blood vessels; Gon, gonads; Pro, prostate; Ute, uterus. ND, not determined. ★, p < 0.05 compared with males.
Tissue Distribution of Ent1 mRNA in Male and Female Rats and Mice. Tissue distribution patterns of Ent1 mRNA are generally similar between rats and mice (Fig. 4), but certain significant species differences do exist. Ent1 mRNA was ubiquitously expressed in all the tissues examined. The highest Ent1 mRNA was found in testes of both species. Ent1 mRNA was also expressed at high to moderate levels in lung and liver of both species, but the lung to liver ratio was much higher in rats (∼5.0) than in mice (0.5-0.7). Mice had high Ent1 mRNA in pituitary, but rats did not. Significant expression of Ent1 mRNA was detected in blood vessels of rats. Rats, but not mice, displayed significant male-predominant expression of Ent1 mRNA in heart, brain, and skeletal muscle. Female mice had 47% higher Ent1 mRNA expression in kidney than males. Rat and mouse testes had 18.3- and 2.8-fold higher Ent1 mRNA than did rat and mouse ovary, respectively.

Tissue distribution of Ent1 mRNA in male and female rats and mice. Ten micrograms of total RNA from various tissues of male and female Sprague-Dawley rats or C57BL/6 mice were analyzed with the Ent1-specific probe set described under Materials and Methods. The y-axis represents the RLU per 10 μg of total tissue RNA. Data represent mean ± standard error of determinations from five samples per group, except the data on pituitary (rats and mice) and ovary (mice), which are the mean of duplicate pooled pituitary or ovary samples from 10 rats or mice. Liv, liver; Kid, kidney; Lun, lung; Sto, stomach; Duo, duodenum; Jej, jejunum; Ile, ileum; L.I., large intestine; Brn, brain; Pit, pituitary; Ht, heart; Thy, thymus; Spl, spleen; Mus, skeletal muscle; B.V., blood vessels; Gon, gonads; Pro, prostate; Ute, uterus. ND, not determined. ★, p < 0.05 compared with males.
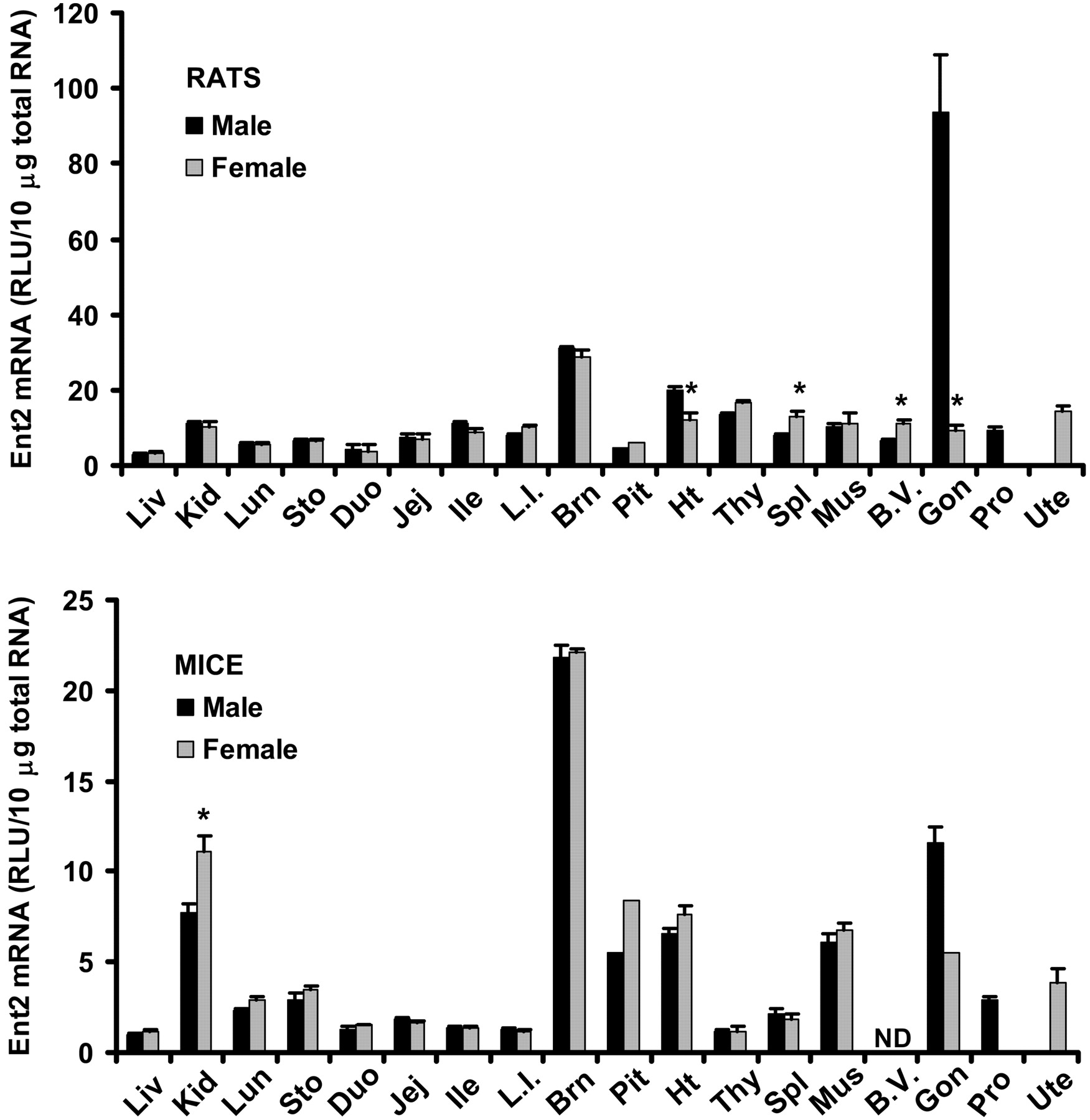
Tissue Distribution of Ent2 mRNA in Male and Female Rats and Mice. Tissue distribution patterns of Ent2 mRNA are similar between rats and mice (Fig. 5). Ent2 mRNA was ubiquitously expressed in all tissues examined. In both species, testes and brain were the two tissues with the highest expression of Ent2 mRNA; however, Ent2 mRNA was the highest in testes of rats, but highest in brain of mice. Rats, but not mice, displayed significant gender-predominant expression of Ent2 mRNA in spleen and blood vessels (63% and 83% higher in females than in males, respectively) as well as heart (67% higher in males than in females). Female mice had 44% higher Ent2 mRNA expression in kidney than males. Rat and mouse testes had 9.3- and 1.1-fold higher Ent2 mRNA than rat and mouse ovary, respectively.

Tissue distribution of Ent2 mRNA in male and female rats and mice. Ten micrograms of total RNA from various tissues of male and female Sprague-Dawley rats or C57BL/6 mice were analyzed with the Ent2-specific probe set described under Materials and Methods. The y-axis represents the RLU per 10 μg of total tissue RNA. Data represent mean ± standard error of determinations from five samples per group, except the data on pituitary (rats and mice) and ovary (mice), which are the mean of duplicate pooled pituitary or ovary samples from 10 rats or mice. Liv, liver; Kid, kidney; Lun, lung; Sto, stomach; Duo, duodenum; Jej, jejunum; Ile, ileum; L.I., large intestine; Brn, brain; Pit, pituitary; Ht, heart; Thy, thymus; Spl, spleen; Mus, skeletal muscle; B.V., blood vessels; Gon, gonads; Pro, prostate; Ute, uterus. ND, not determined. ★, p < 0.05 compared with males.
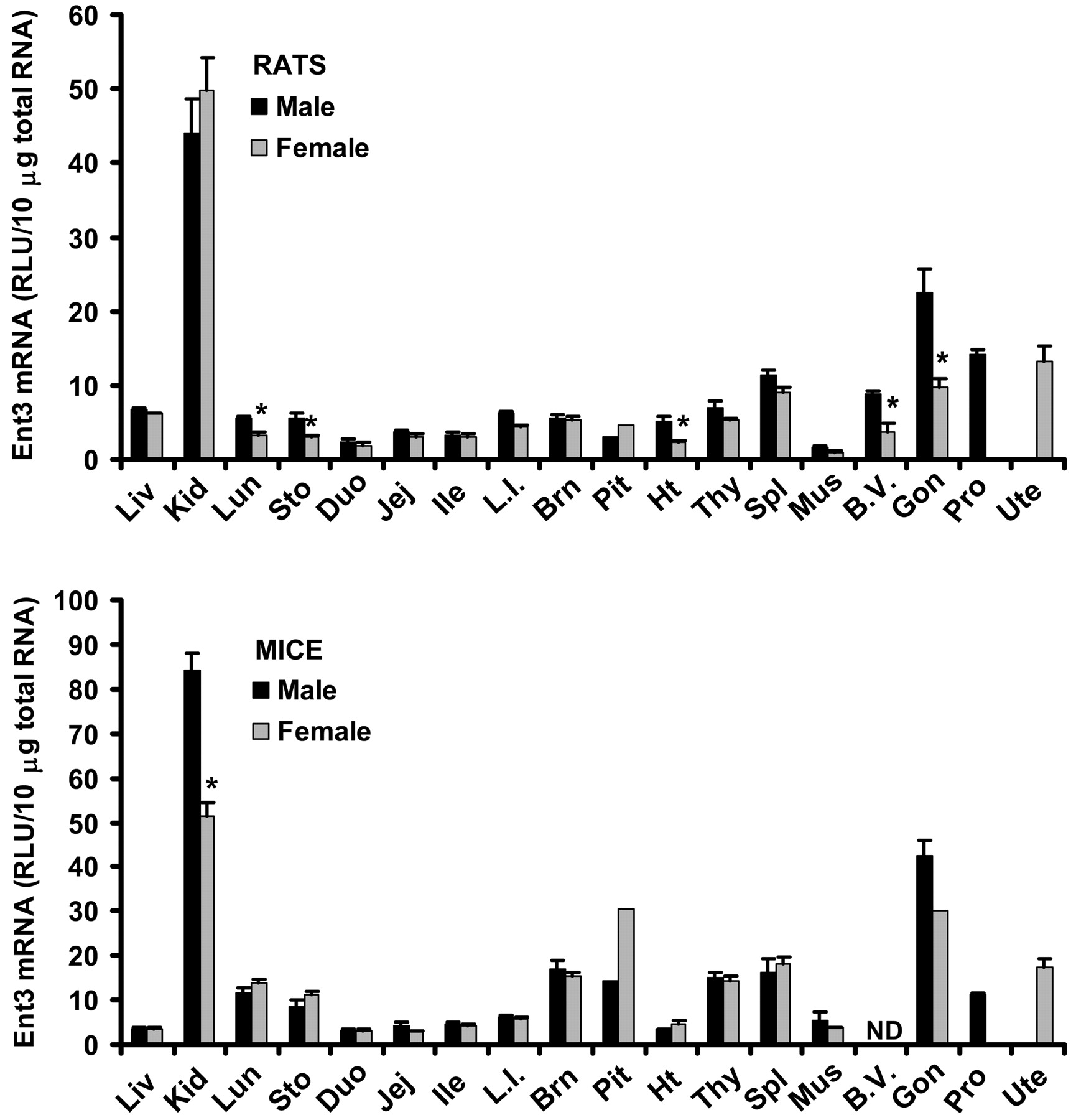
Tissue Distribution of Ent3 mRNA in Male and Female Rats and Mice. Tissue distribution patterns of Ent3 mRNA are very similar between rats and mice (Fig. 6). Similar to Ent1 and 2, Ent3 mRNA was also ubiquitously expressed. In both species, kidney had the highest expression of Ent3 mRNA, followed by testes. Rats exhibited significant male-predominant expression of Ent3 mRNA in lung, stomach, heart, and blood vessels (66, 90, 120, and 143% higher than females, respectively). Male mice had 65% higher Ent3 mRNA in kidney than females. Rat and mouse testes had 131% and 40% higher Ent3 mRNA than rat and mouse ovary, respectively.
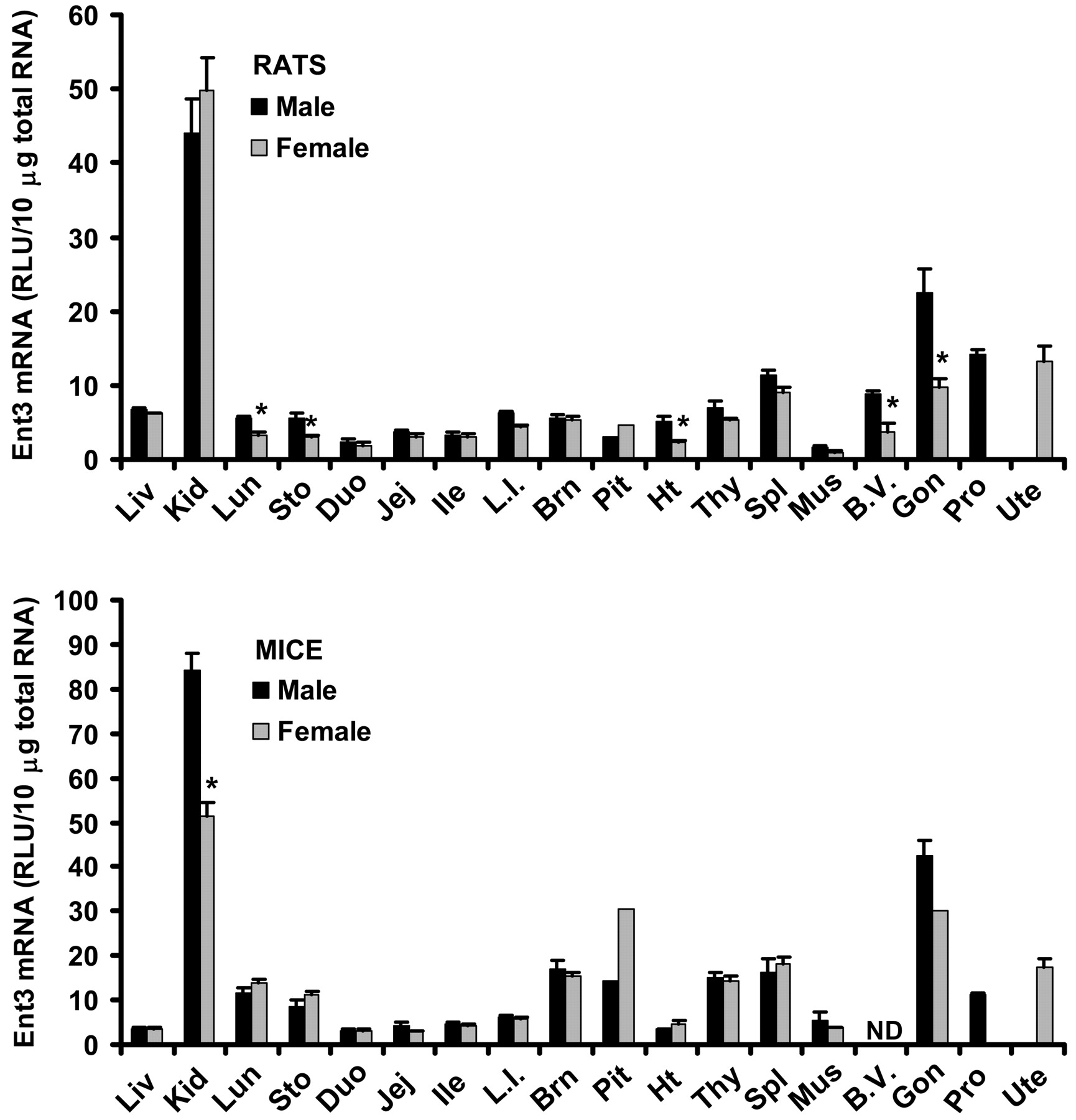
Tissue distribution of Ent3 mRNA in male and female rats and mice. Ten micrograms of total RNA from various tissues of male and female Sprague-Dawley rats or C57BL/6 mice were analyzed with the Ent3-specific probe set described under Materials and Methods. The y-axis represents the RLU per 10 μg of total tissue RNA. Data represent mean ± standard error of determinations from five samples per group, except the data on pituitary (rats and mice) and ovary (mice), which are the mean of duplicate pooled pituitary or ovary samples from 10 rats or mice. Liv, liver; Kid, kidney; Lun, lung; Sto, stomach; Duo, duodenum; Jej, jejunum; Ile, ileum; L.I., large intestine; Brn, brain; Pit, pituitary; Ht, heart; Thy, thymus; Spl, spleen; Mus, skeletal muscle; B.V., blood vessels; Gon, gonads; Pro, prostate; Ute, uterus. ND, not determined. ★, p < 0.05 compared with males.
Discussion
The present study demonstrates that in general, tissue distribution of Cnt and Ent is similar in rats and mice. However, a few marked species differences exist, such as Cnt2 in liver (high in rats; low in mice), Cnt3 in lung (high in rats; low in mice), and Ent1 in pituitary (low in rats; high in mice).
The mouse Cnt1 gene has not been identified in the published literature. However, a BLAST search of GenBank with the rat Cnt1 gene sequence identified a similar cDNA clone (MGC:74379) from mouse kidney (accession number BC061230). The deduced mouse Cnt1 protein contains 648 amino acids, the same length as rat Cnt1 protein, and shares 80% similarity with rat Cnt1 protein. Moreover, tissue distribution of Cnt1 mRNA (using probes against BC061230) in mice is very similar to that in rats (Fig. 1). These data indicate that sequence BC061230 is mouse Cnt1.
A considerable amount of Cnt1 protein has been detected in rat liver; however, using different methods, Cnt1 mRNA was either undetectable or detectable in rat liver (Huang et al., 1994; Felipe et al., 1998). Therefore, Cnt1 mRNA may be expressed at low levels, and undergo significant post-transcriptional regulation, in rat liver (Felipe et al., 1998). The present study shows that Cnt1 mRNA in rat and mouse liver is very low. In contrast, Pennycooke et al. (2001) reported a very high expression of CNT1 mRNA in human liver. Further study on protein expression is needed to draw conclusions on Cnt1 expression in mouse liver.
Cnt2 mRNA is highly expressed in the liver of rats but not in mice (Fig. 2). In contrast, Ent1 mRNA is highly expressed in mouse liver (Fig. 4), but only moderately expressed in rat liver (Fig. 4). Both Cnt2 and Ent1 efficiently transport adenosine, but Cnt2 mediates concentrative and Na+-dependent transport, whereas Ent1 mediates bidirectional and Na+-independent transport. Further study on adenosine uptake by mouse and rat liver may disclose the physiological significance of such a species-divergent expression of Cnt2 and Ent1 mRNA.
The present study demonstrates marked species differences in the tissue distribution of Cnt3 mRNA between rats and mice (Fig. 3). The deduced rat Cnt3 and mouse Cnt3 protein share 86% homology, and they are 72% homologous to human CNT3 protein. In humans, CNT3 transcripts are expressed at high levels in bone marrow, pancreas, trachea, and mammary gland, and at moderate levels in lung, liver, testes, prostate, uterus, and intestine (Ritzel et al., 2001b). However, Cnt3 mRNA is not expressed in liver of rats or mice (Fig. 3), and a separate study in this laboratory shows that Cnt3 is not expressed in bone marrow of rats (data not shown). Therefore, considerable species differences in tissue distribution of CNT3 exist among humans, mice, and rats. Human and mouse CNT3 efficiently transports both pyrimidine and purine anticancer nucleoside drugs (Ritzel et al., 2001b). The marked species differences in CNT3 distribution may have profound impact on evaluating CNT3 substrates during drug development.
The tissue expression pattern of mouse Ent1 (Fig. 4) is generally consistent with a previous report (Choi et al., 2000). Rat Ent1 and 2 have been functionally characterized, but their tissue distribution remained largely unknown (Yao et al., 1997, 2002). In skeletal muscle, Ent1 and 2 are only moderately expressed in rats and mice (Figs. 4 and 5), which is different from the very abundant expression of ENT2 in humans (Baldwin et al., 2004). ENT2 transports various nucleoside and nucleobase analogs, and may be important in transporting adenine and adenosine metabolites inosine and hypoxanthine (Ward et al., 2000), which are essential for energy metabolism in skeletal muscle. The apparent marked species difference between humans and rodents in skeletal muscular expression of Ent2 should be considered in development and clinical application of nucleoside and nucleobase drugs.
A preliminary report indicates that Ent3 has substrate specificity similar to that of Ent2 and may be predominantly localized to intracellular compartments (Baldwin et al., 2004). The present study shows that tissue expression patterns of Ent3 are very similar between rats and mice. The consistent high expression of Ent3 in kidney across species suggests that Ent3 may be important in the physiological functioning of kidney, probably facilitating the intracellular trafficking of nucleosides and nucleobases.
The present study demonstrates that Ent2 transcript is expressed at high levels, whereas Cnt1 and Cnt2 transcripts are expressed at low levels in the brain (Figs. 1, 2, and 5). In contrast, in choroid plexus, which locates in the ventricles of the brain, Ent2 is expressed at low levels, whereas Cnt1 and Cnt2 transcripts are at high levels (Choudhuri et al., 2003). Such regional difference in expression of nucleoside transporters may reflect their different physiological functions. For example, brain has high energy consumption, and Ent2 is unique in efficiently transporting nucleobases, important for energy metabolism (Yao et al., 2002).
Ent1 and 2 are broadly distributed in different regions of brain, and are expressed in cells rich in adenosine receptors (Anderson et al., 1999a,b). The hypothalamic-pituitary-adrenal axis plays a key role in regulation of endocrine and immune responses, in which the local concentration of adenosine in the pituitary may be of paramount importance (Rees et al., 2003). The present study provides the first evidence that various nucleoside transporters are expressed in pituitary. Through regulating local concentrations of adenosine, brain and pituitary ENTs may be essential in regulating endocrine and immune responses. The physiological significance of divergent expression of Ent1 (brain < pituitary) and Ent2 (brain > pituitary) in mouse brain and pituitary warrants further investigation.
Gender differences in expression patterns of transporters may have profound effects on basal physiology as well as drug efficacy and toxicity. The present study provides the first evidence of significant gender differences in expression of various nucleoside transporters in rats and mice. An intriguing observation is the gender-divergent expression of Ents in mouse kidney; males have lower expression of Ent1 (Fig. 4) and Ent2 (Fig. 5), but higher expression of Ent3 (Fig. 6) than females. In humans, adenine phosphoribosyltransferase deficiency causes a defect in AMP synthesis from adenine; as a result, adenine is oxidized to poorly water-soluble 2,8-dihydroxyadenine, which precipitates in the urine or kidney, causing nephrolithiasis and chronic renal failure. Interestingly, in an adenine phosphoribosyltransferase-null mouse model of nephrolithiasis, males have more pronounced kidney damage than females (Stockelman et al., 1998). The observed gender-divergent expression of Ents and the resultant gender difference in renal excretion and metabolism of adenine could be a major molecular mechanism underlying such a gender difference.
The present study shows that in heart, male and female rats express similar levels of Cnt1, 2, and 3 transcripts, but females have significantly lower levels of Ent1, 2, and 3 transcripts than males. In both humans and rats, males are known to be more vulnerable to cardiovascular diseases than females. Adenosine exerts its potent cardiovascular protective effect through increasing local blood flow (Lasley and Smart, 2001). Dipyridamole and nitrobenzylthioinosine, the two inhibitors of ENTs, protect the heart from ischemia/reperfusion-induced toxicity by increasing the concentration and efficacy of adenosine. Recently, Gamboa et al. (2003) reported that inhibitor-sensitive nucleoside transporters (ENTs) may function as an efflux barrier to prevent intravascular adenosine into the interstitium of heart or blood vessels. Currently, there is no report regarding gender-related information on expression of nucleoside transporters in human heart. Should women have lower expression of ENTs in heart (as observed in rats), this may result in a better uptake of adenosine and, thus, a significantly higher basal myocardial blood flow (Chareonthaitawee et al., 2001) and better protection against certain cardiovascular diseases than men. Interestingly, the present study also demonstrates significant gender differences in expression of Cnt2, Ent2, and Ent3 in blood vessels of rats. Further study on the putative gender differences in expression of ENTs and/or CNTs in the cardiovascular tissues of humans is warranted.
The present study shows that multiple Cnts and Ents are expressed in the sex tissues of rats and mice, such as testes, prostate, ovary, and uterus. Importantly, the considerable expression of all six types of nucleoside transporters in testes may contribute to the high sensitivity of testicular cancer to chemotherapy (Chaganti and Houldsworth, 2000) and the vulnerability of testes to nucleoside drug-induced toxicity. Moreover, previous study demonstrates that Cnt2, Ent1, and Ent2 transcripts are expressed in placenta at levels comparable to those in liver/kidney (Leazer and Klaassen, 2003). Thus, effects of nucleosides and nucleoside analogs on male and female reproduction necessitate scrutiny.
In conclusion, nucleoside transporters have essential physiological functions, and they are important in the disposition of anticancer and antiviral nucleoside analogs (Damaraju et al., 2003). The present study provides the most complete information to date on mRNA tissue expression patterns of all known Cnts and Ents in rats and mice. The observed species and gender differences could be responsible for some species and gender differences in disposition, efficacy, and toxicity of endobiotics and xenobiotics that are substrates for these transporters. Such knowledge is crucial for understanding the similarities and differences between humans and experimental animals regarding drug development and carcinogenesis.
Acknowledgments
We thank Drs. Susan Buist, Terrilyn Couch, Matthew Dieter, Tyra Leazer, Ning Li, and Angela Slitt, and Xingguo Cheng, Jonathan Maher, and Melinda Shelby for technical assistance.
Footnotes
-
Grant support: National Institutes of Health Grants ES-09649 and ES-09716.
-
doi:10.1124/dmd.104.001123.
-
ABBREVIATIONS: CNT/Cnt, concentrative nucleoside transporter; ENT/Ent, equilibrative nucleoside transporter; bDNA, branched DNA; RLU, relative light units.
- Received June 21, 2004.
- Accepted September 13, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}