


Abstract
Root extracts from kava-kava (Piper methysticum G. Forst) are clinically used for the treatment of anxiety and restlessness. Due to reported cases of liver toxicity, kava-kava extracts were withdrawn from the market in several countries in 2002. Because the efflux transporter P-glycoprotein (P-gp) is involved in the absorption, distribution, and excretion of many drugs and often participates in drug-drug interactions, we studied the effect of a crude kava extract and the main kavalactones kavain, dihydrokavain, methysticin, dihydromethysticin, yangonin, and desmethoxyyangonin on the P-gp-mediated efflux of calcein-acetoxymethylester in the P-gp-overexpressing cell line P388/dx and the corresponding cell line P388. The crude extract and the kavalactones showed a moderate to potent inhibitory activity with f2 (concentration needed to double baseline fluorescence) values of 170 μg/ml and 17 to 90 μM, respectively. The f2 value of yangonin could not be determined due to its higher lipophilicity. In conclusion, our results for the first time demonstrate P-gp-inhibitory activity of kava-kava and its components in vitro.
Extracts made from the root-stock of kava-kava (Piper methysticum G. Forst), a perennial shrub native to the South Pacific Islands, are traditionally used for their calming and relaxing effects. For over a decade, commercial kava-kava extracts have been marketed as dietary supplements for the treatment of stress, insomnia, and anxiety (Côté et al., 2004). The kavalactones (kavapyrones) kavain, dihydrokavain, methysticin, dihydromethysticin, yangonin, and desmethoxyyangonin are the main ingredients and were found to be the major pharmacologically active principle (Singh and Singh, 2002). Therefore, extracts are standardized to a certain amount of kavalactones. In 2002, kava-kava extracts were withdrawn from the market in Germany, United Kingdom, France, Switzerland, Australia, and Canada due to reported cases of liver toxicity (Teschke et al., 2003; Anke and Ramzan, 2004a,b; Clouatre, 2004). The reason for the observed liver toxicity is unknown. It has been suggested that the extraction process of the crude root powder results in chemical changes that may be related to hepatotoxicity in some preparations (Whitton et al., 2003), but interactions with other drugs or herbal supplements might also be the underlying cause.
Several human cytochrome P450 isoenzymes (P450s) are inhibited by kava-kava extracts and kavalactones in vitro (Unger et al., 2002; Côté et al., 2004; Unger and Frank, 2004; Zou et al., 2004) and in vivo (Gurley et al., 2005; Russmann et al., 2005). Hence, interactions at the level of P450s are conceivable. ABC-transporters, especially P-glycoprotein (P-gp), are another possible site of interaction. P-gp, the product of the MDR1/ABCB1 gene, is involved in the absorption, distribution, and excretion of many drugs (Schinkel and Jonker, 2003). Clinically relevant drug interactions are possible if the plasma or tissue concentrations of drugs with a narrow therapeutic range are increased or decreased as a consequence of the inhibition or induction of P-gp (Lin, 2003). However, the inhibition or induction of P-gp is not limited to synthetic drugs. Various natural products are known to influence the P-glycoprotein-mediated efflux of chemicals (Bhardwaj et al., 2002; Choi et al., 2004; Zhou et al., 2004). Recently, various St. John's wort constituents were screened for their inhibitory activity on P-gp, and hypericin and hyperforin were identified as potent inhibitors of the P-gp-mediated efflux of the substrates calcein-acetoxymethylester (calcein-AM) and daunorubicin in vitro (Wang et al., 2004). Furthermore, in vivo, extracts of St. John's wort caused a clinically relevant decrease of the plasma levels of P-gp and CYP3A4 substrates (Dresser et al., 2003; Izzo, 2004).
So far, nothing is known about possible interactions of kava-kava with P-gp. We therefore tested a crude methanolic (80% v/v) kava-kava root extract and the main kavalactones for their P-gp-inhibitory activity in the well established calcein assay (Weiss et al., 2003; Fröhlich et al., 2004) using the P-gp-overexpressing cell line P388/dx as well as the corresponding parental cell line P388.
Materials and Methods
Materials. Medium, medium supplements, and Hanks' balanced salt solution were purchased from Invitrogen (Karlsruhe, Germany), fetal calf serum was purchased from Biochrom AG (Berlin, Germany), and doxorubicin hydrochloride and verapamil were purchased from Sigma-Aldrich (Taufkirchen, Germany). DMSO and Triton X-100 were obtained from AppliChem (Darmstadt, Germany) and calcein-AM from MoBiTec (Göttingen, Germany). Kavain and quinidine were purchased from Roth (Karlsruhe, Germany). Kava-kava root powder (standardized to 2.5% kavalactones; batch 921552) was taken from Nature's Way Single Herbal capsules (Springville, UT). The kavalactones dihydrokavain, methysticin, dihydromethysticin, yangonin, and desmethoxyyangonin were obtained from Addipharma (Hamburg, Germany).
P388 and P388/dx Cells. As the model for P-gp we used the murine monocytic leukemia cell line P388 and the corresponding doxorubicin-resistant cell line P388/dx overexpressing mdr1a/1b (Boesch et al., 1991). Both cell lines were kindly provided by Dr. Dario Ballinari (Pharmacia and Upjohn, Milan, Italy). The culture conditions and the suitability of this cell system to evaluate P-gp-inhibitory effects have been published previously (Fröhlich et al., 2004).
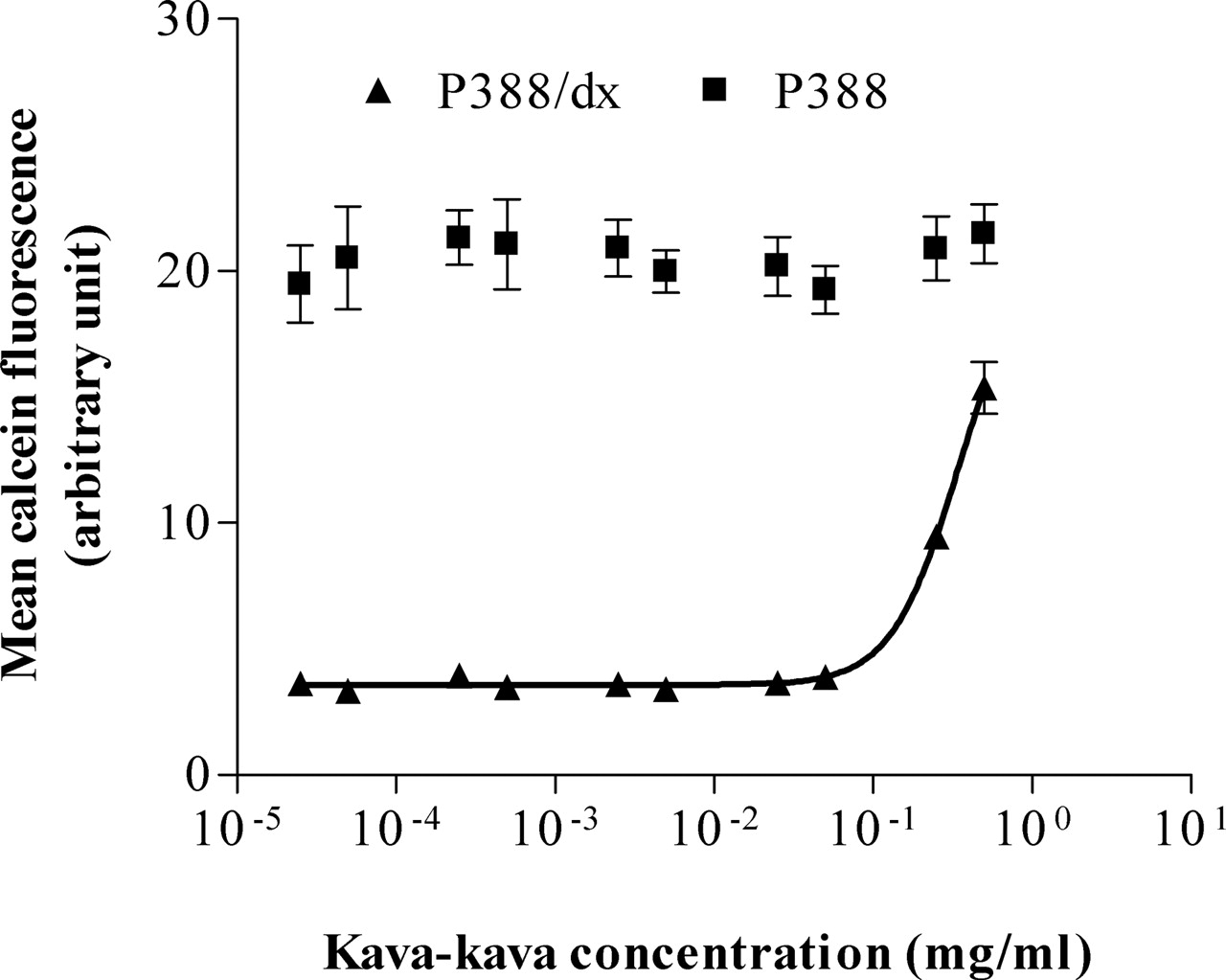
Calcein assay. Concentration-dependent effect of kava-kava on the calcein accumulation in P388 and P388/dx cells. Each curve depicts one representative experiment of a series of three. Data are expressed as mean ± S.D. for n = 8 wells.
Stock Solutions. The kava-kava root powder (1 g) was extracted with 10 ml of aqueous methanol (80% v/v) in an ultrasonic bath for 30 min. After centrifugation at 9000g for 10 min, the resulting yellow supernatant was stored in the dark at room temperature. All test compounds were dissolved in DMSO (50 mM stock solutions) and tested up to the highest soluble concentration. The DMSO concentration in the assays never exceeded 1% (v/v), a concentration that was found not to influence the results of the assay in pilot experiments.
Calcein Uptake Assay and Cytotoxicity Assay. To determine the P-gp inhibitor potency of the test compounds, their effects on the uptake of calcein-AM in P388/dx and P388 cells were investigated as published previously (Fröhlich et al., 2004). In this assay, inhibition of P-pg leads to an accumulation of intracellular calcein fluorescence. Cells were used at a concentration of 5 × 105 cells/well. Each experiment was performed at least in triplicate on different days.
The concentration range used in the calcein assays was also screened for possible cytotoxic effects of the test compounds in each cell line with the Cytotoxicity Detection Kit (Roche Applied Science, Mannheim, Germany), because cytotoxic effects may diminish the increase in fluorescence in the calcein assay, leading to underestimation of P-gp-inhibitory potency. A detailed description of the methods and the validation of the calcein assay have been published previously (Weiss et al., 2003).
Statistical Analysis. Due to the limited solubility of the compounds investigated, no plateau effects could be reached in the calcein assay, eliminating the possibility of the calculation of IC50 values (concentration leading to 50% of the maximal effect). Therefore a previously validated alternative method for calculation of the inhibitory activity, the f2 value (inhibitor concentration needed to double baseline fluorescence), was determined (Weiss et al., 2003).
Data are presented as means ± standard deviation (S.D.) and were analyzed using GraphPad Prism Version 4.0 (GraphPad Software Inc., San Diego, CA). Statistical significance was assessed using analysis of variance with Dunnett's multiple comparison test for post hoc pairwise comparison of the results with the control (verapamil). A p value <0.05 was considered significant.
Results
All compounds tested increased the intracellular calcein fluorescence in the P-gp-overexpressing cell line P388/dx (Figs. 1 and 2), but not in the parental cell line P388 (only shown for kava-kava; Fig. 1), indicating P-gp-inhibitory activity, which was concentration-dependent. None of the test compounds revealed cytotoxic effects in P388 and P388/dx cells or quenched the calcein fluorescence.
Due to their low solubility, none of the test compounds reached plateau effects. Thus, for comparison of the inhibitory potencies, f2 values instead of IC50 values were calculated (Table 1). For yangonin, the kavalactone with the lowest solubility in HEPES-supplemented Hanks' balanced salt solution, the f2 value could not be determined. Nevertheless, a clear effect of yangonin on the intracellular calcein concentration can be seen in Fig. 2C.
Inhibition of P-gp by an extract and kavalactones of Piper methysticum in the P-gp-overexpressing cell line P388/dx
Values represent mean ± S.D. of at least three independent assays. p values were determined by analysis of variance with Dunnett's multiple comparison test for post hoc pairwise comparison of the results with the verapamil control.

Calcein assay. Concentration-dependent effect of kavain and dihydrokavain (A), desmethoxyyangonin and dihydromethysticin (B), and yangonin and methysticin (C) on the calcein accumulation in P388/dx cells expressing murine mdr1a/mdr1b. Each curve depicts one representative experiment of a series of three to four. Data are expressed as mean ± S.D. for n = 8 wells.
Discussion
Patients often conceal the use of herbal remedies because they are generally accepted to be safe and harmless (Martin-Facklam et al., 2004). However, if taken together with synthetic drugs (or other herbal remedies), the possibility of adverse effects increases due to pharmacokinetic interactions (Ioannides, 2002), as has been scrutinized for St. John's wort (Moore et al., 2000; Dresser et al., 2003; Rengelshausen et al., 2005). For kava-kava there are several reports of liver toxicity (Teschke et al., 2003; Anke and Ramzan, 2004a,b; Clouatre, 2004), which might be due to interactions with simultaneously applied drugs. Several studies have demonstrated the inhibition of P450s by kava-kava and its components in vitro (Unger et al., 2002; Côté et al., 2004; Unger and Frank, 2004; Zou et al., 2004) and in vivo (Gurley et al., 2005; Russmann et al., 2005) but, hitherto, no data were available on the interaction of kava-kava with P-gp.
This study quantified the P-gp-inhibitory potency of kava-kava and the kavalactones in the previously established and validated calcein assay (Weiss et al., 2003; Fröhlich et al., 2004).
All compounds revealed concentration-dependent P-gp-inhibitory activity. The crude kava-kava root extract made with aqueous MeOH (80% v/v) inhibited the P-gp-mediated efflux of calcein-AM with a potency comparable to the inhibitory activity on CYP3A4 determined by Unger et al. (2002) (Fig. 1; Table 1).
Also, the kavalactones showed a moderate to potent effect on the intracellular accumulation of calcein-AM with f2 values between 10 and 90 μM (Table 1; Fig. 2). Concerning their inhibitory potency, the kavalactones can be divided into three distinct groups with f2 values of 88.1 ± 25.2 and 88.6 ± 13.9 μM (kavain/dihydrokavain), 47.5 ± 17.1 and 54.6 ± 10.1 μM (methysticin/dihydromethysticin), and 17.1 ± 0.6 μM for desmethoxyyangonin (Table 1; Fig. 2). Compared with desmethoxyyangonin, the inhibition curve for yangonin is shifted to the left with an exponential increase in the intracellular calcein concentration at about 1 μM. Thus, it can be speculated that the inhibitory potency of yangonin is comparable to or even greater than that of desmethoxyyangonin.
This study for the first time demonstrates a concentration-dependent, moderate to potent inhibition of P-gp by kava-kava root extracts and its major components, the kavalactones. These in vitro results enable the hypothesis that the intake of kava-kava extracts might lead to possible pharmacokinetic interactions with concomitantly used P-gp substrates. However, almost nothing is known about concentrations of kavalactones in the intestine, other organs, or plasma. So far, only kavain was tested for its ability to reach the systemic circulation in humans (Tarbah et al., 2003). A single oral dose of 800 mg of kavain resulted in a serum concentration of 40 ng/ml (0.1 μM) after 1 h. This plasma level is unlikely to cause a significant inhibition of P-gp in vivo. However, following oral application, the concentration of kavalactones might be considerably higher in the gut wall. Assuming that a single dose of kava-kava extract that contains 7 to 17 mg of the individual kavalactones (Gurley et al., 2005) is taken together with a 200-ml volume of beverage, a rough estimation of the concentration of each kavalactone yields concentrations between 35 and 85 μg/ml (136–370 μM) in the gut. In comparison to the plasma levels, these concentrations are higher than the obtained f2 values and are therefore sufficient for inhibition of P-gp in endothelial cells of the intestine.
Interestingly, the results obtained with in vitro and in vivo studies investigating the inhibitory effect of kava-kava extracts and kavalactones on P450s are contradictory. In vitro inhibition of CYP1A2, CYP2C8, CYP2C9, CYP2C19, CYP2D6, CYP2E1, and CYP3A4 by kava-kava extracts could be demonstrated by several groups (Unger et al., 2002; Côté et al., 2004; Unger and Frank, 2004; Zou et al., 2004). In contrast, one in vivo study demonstrated inhibition of CYP1A2 (Russmann et al., 2005), whereas another one detected inhibition only for CYP2E1 but not for CYP1A2, CYP2D6, and CYP3A4 (Gurley et al., 2005). These contradictory results substantiate the fact that the extrapolation of in vitro results on the in vivo situation always remains speculative. Hence, there is a clear need for in vivo studies investigating whether the P-gp inhibition found in vitro by kava-kava and its constituents also occurs in vivo and whether this is of relevance for drug interactions in vivo.
Acknowledgments
We thank Dr. Dario Ballinari (Pharmacia and Upjohn, Milan, Italy) for the cell lines P388 and P388/dx.
Footnotes
-
This work was supported by Grant 01EC9902 from the German Ministry for Education and Research (Bundesministerium für Bildung und Forschung).
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.105.005892.
-
ABBREVIATIONS: P450, cytochrome P450; P-gp, P-glycoprotein; calcein-AM, calcein-acetoxymethylester; DMSO, dimethyl sulfoxide; f2, concentration needed to double baseline fluorescence.
- Received June 1, 2005.
- Accepted July 27, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}