


Abstract
Cytochromes P450 3A (CYP3A) and P-glycoprotein (P-gp) are mainly located in enterocytes and hepatocytes. The CYP3A/P-gp system contributes to the first-pass metabolism of many drugs, resulting in a limited bioavailability. During the neonatal period, a shift between CYP3A7, the fetal form, and CYP3A4 occurs in the liver, but data on the expression of the CYP3A/P-gp complex in the intestine are very limited. A total of 59 normal duodenal biopsies from white children aged 1 month to 17 years were studied. Localization of the proteins by immunohistochemistry analysis was performed using a polyclonal antibody, Nuage anti-CYP3A, and a monoclonal antibody, C494 anti-P-gp. The mRNA quantification was performed using highly specific real-time reverse transcription-polymerase chain reaction. Villin mRNA quantification was used for normalization. CYP3A protein was detected in all enterocytes in the samples from patients over 6 months of age, whereas it was not in younger samples. P-gp protein was expressed at the apical and upper lateral surfaces of the enterocytes. CYP3A isoforms and P-gp mRNA levels were highly variable. CYP3A4 and CYP3A5 mRNA levels were high during the first year of life and decreased with age, whereas CYP3A7 was detected at a low level in 64% of samples, whatever the age. P-gp mRNA expression level was also highly variable. Our results showed that neonates and infants had a significant expression of CYP3A and P-gp mRNA in the intestine, suggesting a different maturation profile of CYP3A and P-gp with age in the liver and the intestine.
Cytochrome P450 isoenzymes are part of the phase I metabolizing enzymes. The CYP3A subfamily is predominant, accounting for almost 30% of the cytochromes in the liver and 70% of the cytochromes in the small intestine, and is involved in the metabolism of more than 70% of currently administered drugs (Paine et al., 1997; Wacher et al., 1998). The CYP3A subfamily consists of three major isoforms: CYP3A4, CYP3A5, and CYP3A7, and a recently identified isoform, CYP3A43. They are closely related since they share at least 83% amino acid sequence identity. In adults, CYP3A4 is the predominant form in the liver and the intestinal tract (Koch et al., 2002). It is variably expressed, but none of the identified genetic polymorphisms showed relevant clinical effect (Lown et al., 1994; Wacher et al., 1998; Wandel et al., 2000). CYP3A5 is present in the liver (Wrighton et al., 1990; Schuetz et al., 1994) and intestine (Koch et al., 2002). The variability of CYP3A5 expression is under genetic control. The contribution of CYP3A5 to drug metabolism is important since CYP3A5 can contribute to over 50% of total CYP3A content (Hustert et al., 2001; Kuehl et al., 2001). CYP3A7 is the major cytochrome P450 isoform detected in embryonic, fetal, and newborn liver (Schuetz et al., 1994; Lacroix et al., 1997). It is also expressed, at a low level, in the adult liver and intestine (Koch et al., 2002). P-glycoprotein (MDR1) is a plasma membrane glycoprotein belonging to the ATP-binding cassette transporters superfamily (Tanigawara, 2000) acting as an efflux system contributing to the protection of the organism. It is located in many tissues and specifically within the brush border at the apical surface of mature enterocytes in the small intestine (Thiebaut et al., 1987). Its expression is genetically controlled (Brinkmann et al., 2001; Cascorbi et al., 2001). CYP3A and P-gp share common substrates, and their coordinated action in the intestine results in a reduced bioavailability of drugs administered orally (Wacher et al., 1998; Paine et al., 1996). Although many of these drugs are used in children, studies concerning the intestinal ontogeny of CYP3A and P-gp are scarce. In the present study, we report new data regarding the localization and the mRNA expression of the three isoforms of CYP3A and P-gp in the duodenum of infants and children aged 1 month to 17 years.
Materials and Methods
Patient Samples. Fifty-nine duodenal biopsies (second or third part of duodenum) from white patients aged 1 month to 17 years, included in the tissue bank of the department of Anatomopathology, were selected retrospectively. These biopsies were performed for medical investigations (growth retardation, gastrointestinal reflux or pain, hypotrophy, or diarrhea) and were found normal after histological examination. Biopsies from patients receiving known inducers or inhibitors of CYP3A and P-gp were excluded. The study was conducted according to the French legislation. Snap-frozen tissues were used for mRNA quantification and formalin-embedded tissues were used for immunohistochemistry analysis.
Immunohistochemistry Analysis. Staining of duodenal biopsies was performed using a three-step immunoperoxidase (sandwich) technique. For CYP3A, the primary antibody was a rabbit anti-human polyclonal CYP3A antibody called “Nuage;” the secondary antibody was a goat anti-rabbit antibody purchased from MP Biomedicals (Irvine, CA) and the tertiary antibody was a rabbit peroxidase anti-peroxidase complex obtained from DakoCytomation California Inc. (Carpinteria, CA). For P-gp, a mouse anti-human monoclonal P-gp antibody called “C494” was purchased from DakoCytomation, the biotinylated sheep anti-mouse antibody (Silenus) was obtained from Chemicon (Temecula, CA), and the streptavidine peroxidase complex was obtained from Boehringer Ingelheim GmbH (Ingelheim, Germany). The detection for both reactions was conducted with diaminobenzidine (DAB Kit from Menarini, Florence, Italy). Negative controls (without adding the primary antibody) showed absence of staining, and adult liver and intestine sections were used for the validation of the CYP3A- and P-gp-specific staining (data not shown). Sections were viewed using a microscope (Leitz Laborlux).
RT-PCR.Extraction of Total RNA and Reverse Transcription. Total RNA was extracted from human duodenal biopsies according to the method described by the manufacturer with the RNA Plus solution (Qbiogene, Illkirch, France). The reverse transcription (RT) reaction was performed on 1 μg of the extracted total RNA.
Quantitative Real-Time PCR with TaqMan Detection. The quantitative real-time PCR analysis system allows an accurate, specific, and reproducible quantification of mRNA. Forward and reverse primers and the fluorogenic TaqMan probe for CYP3A isoforms and villin were designed to span exon junctions in the fully processed mRNA using Primer Express software (Applied Biosystems, Foster City, CA). P-gp primers and probe sequences were the courtesy of Dr. V. Barbu (Pharmacy, Hôpital Robert Debré, Paris, France). 5-Carboxyfluorescein was used as the 5′-fluorescent reporter and 5-carboxytetramethylrhodamine was added to the 3′-end as a quencher for CYP3A5, CYP3A7, P-gp, and villin probes; an MGB probe was used for CYP3A4. Primers and probe sequences are listed in Table 1. Primers were purchased from Genset/Proligo (Boulder, CO) and probes from Applied Biosystems.
Primers and probes sequences for real-time quantitative PCR (TaqMan)
PCR Conditions. cDNA (50 ng) resulting from RT was subjected to PCR amplification in the ABI 7700 Sequence Detection System (Applied Biosystems). For CYP3A5, CYP3A7, P-gp, and villin, the final reaction conditions were 200 nM forward and reverse primers, 100 nM probe, and 1× Master Mix (Applied Biosystems). For CYP3A4 we used the Core Reagent Mix (Applied Biosystems) with 400 nM primers, 100 nM MGB probe, and 7 mM MgCl2. For 18S, we used the Applied Biosystems kit (ref. 4319413E). PCR conditions for the six genes were 2 min at 50°C, 10 min at 95°C, followed by 50 cycles each of 15 s at 95°C and 1 min at 60°C. Each sample was analyzed in duplicate. Negative and positive controls were present in all the assays. Data were analyzed using the Sequence Detector V1.6 program (Applied Biosystems).
PCR Specificity Control and Normalization. The specificity of the assays was determined using plasmids specifically expressing the gene of interest. The two plasmids expressing, respectively, CYP3A4 and CYP3A5 were the courtesy of Pr. P. Beaune (Biochemistry, Hôpital Europeen Georges Pompidou, Paris, France) the one expressing CYP3A7 the courtesy of Dr. T. Cresteil (Institut de Chimie des Substances Naturelles, Centre National de la Recherche Scientifique, Gif-sur-Yvette, France) and the one expressing P-gp the courtesy of Dr. V. Barbu. The villin plasmid was constructed using the TOPO TA cloning kit with pCR2.1-TOPO as the plasmid vector (Invitrogen, Carlsbad, CA). The assay specificity was verified for each CYP3A isoform by testing the amplification of primers and probes of the gene of interest versus the plasmids of the two other isoforms. In all cases, the amplification with the target gene was at least 105-fold higher than with the other isoforms. The CYP3A5 probe recognizes the corresponding pseudogene. Standard curves were constructed with the use of serial 10-fold dilutions ranging from 102 to 107 copies of the plasmid containing the target gene. Villin gene expression was chosen to normalize the different mRNA quantifications with respect to the depth of the biopsy and proteolysis. The ubiquitously expressed gene, 18S, was used to control the extraction and the RT step efficacy. Data used for statistical tests were expressed as the ratio of the number of copies of the target gene mRNA over the number of villin mRNA copies (ratio of the target gene over V; e.g., CYP3A4/V).
Statistics. Statistical analyses were conducted with the use of Stata Software version 7 (Stata Corporation, College Station, TX). Quantitative variables by age subgroups were expressed as percentiles because of skewed distribution. The relationships between age and the different genes were assessed using median regression. A p value less than 0.05 was considered statistically significant.
Results
Immunohistochemical Localization of CYP3A and P-gp Proteins. CYP3A protein was detected in all biopsies and located within enterocytes. In the samples obtained from patients younger than 6 months of age, CYP3A protein was detected in half of the enterocytes (staining in cells with no staining in adjacent ones) (Fig. 1A), whereas it was expressed in all the cells in older patients (Fig. 1B). P-gp protein was detected in all the enterocytes and located on the apical surface (Fig. 1, C and D). In children aged less than 3 years, additional staining was located on a limited upper part of the lateral surface. The expression variability of both CYP3A and P-gp proteins was observed between enterocytes in the same villous tip and between adjacent villi.
CYP3A and P-gp mRNA Quantification. A significant correlation was evidenced between 18S and villin mRNA (n = 59, r = 0.92; p < 0.001). The mRNAs of the three different CYP3A isoenzymes were quantified separately and their expressions were highly variable between samples. CYP3A4 and CYP3A5 mRNA levels were in the same order of magnitude, whereas CYP3A7 levels were much lower. P-gp mRNA levels were similar to those of CYP3A4, although P-gp was overexpressed in some samples. For CYP3A4, the mRNA quantification varied more than 100-fold with a 3A4/V mRNA ratio ranging from 4 × 10–4 to 0.5. There was a significant relation between CYP3A4 and age (p = 0.0001). Expression was high in the first year of life (CYP3A4/V ratio from 0.04 to 0.5) and decreased with age to reach lower values in older children (CYP3A4/V ratio from 0.01 to 0.2). CYP3A5 mRNA was expressed in all but one sample. The CYP3A5/V ratio ranged from 10–3 to 0.46 and decreased with age without reaching statistical significance (p = 0.061). CYP3A7 mRNA was evidenced in 37 of 58 (64%) patients but at a much lower level than CYP3A4 and CYP3A5. The CYP3A7/V ratio ranged from 16 × 10–6 to 5 × 10–4, with two patients showing relatively high ratios of 0.001 and 0.002 but with no statistical relation with age (p = 0.97). Pooled CYP3A mRNA, expressed as the CYP3A/V ratio was higher in the first year of life (CYP3A/V ratio ranging from 0.029 to 0.9) than in older children. CYP3A4 mRNA accounted for 58.6 ± 20% of total CYP3A mRNA expression (range, 17.5–100%). CYPA5 contributed to more than 50% of total CYP3A mRNA when CYP3A5/V ratio was higher than 0.08. P-gp mRNA was detected in all the samples. Here again, expression was highly variable between samples, the P-gp/V ratio ranging from 0.005 to 4, with nine patients having a ratio higher than 1.3. No significant relation was found between P-gp and age (p = 0.195).
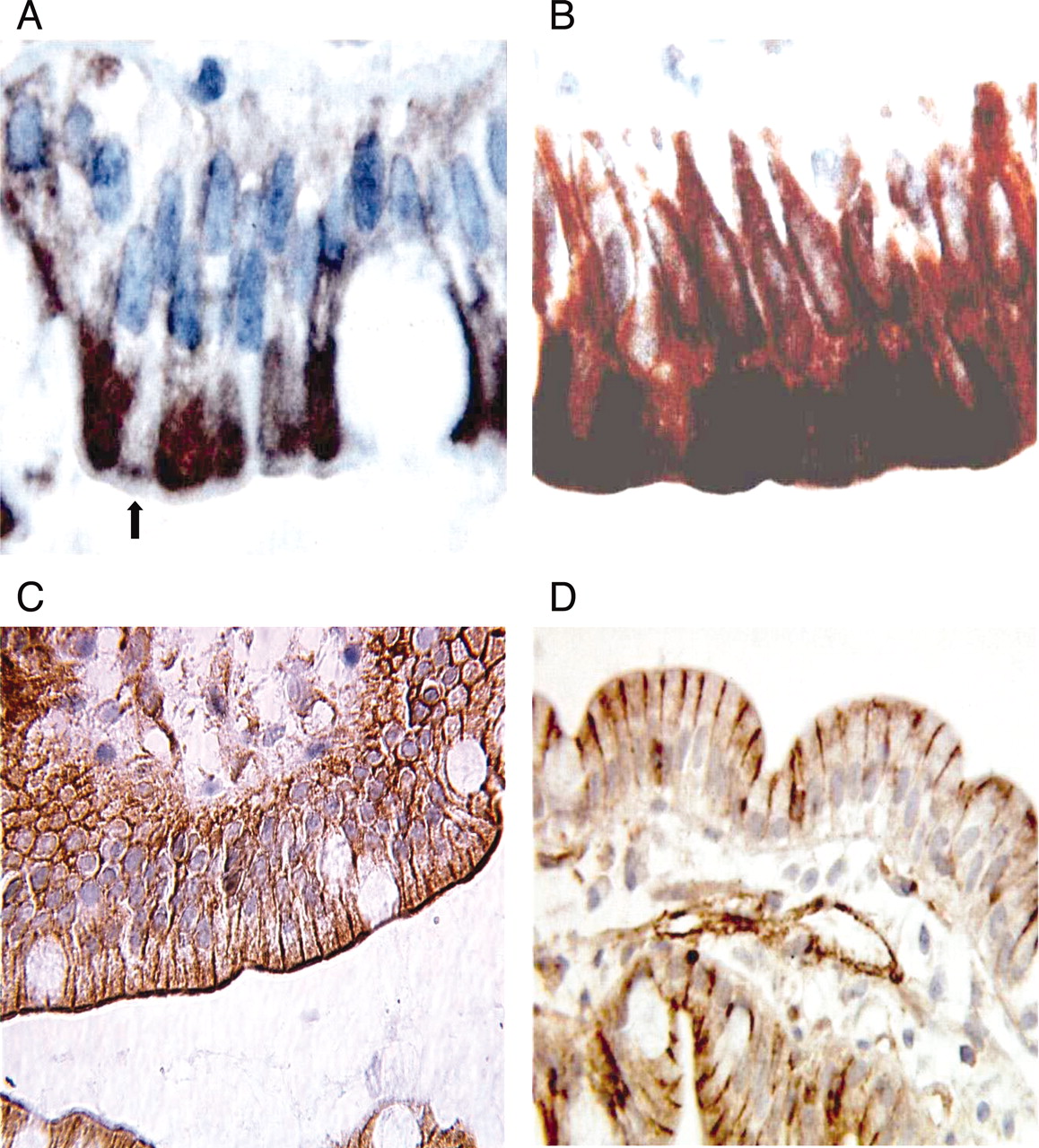
Immunohistochemistry on formalin-embedded duodenal tissue sections with anti-CYP3A polyclonal antibody Nuage and anti-P-gp monoclonal antibody C494. Some enterocytes do not express CYP3A in the samples from patients aged less than 6 months, whereas all enterocytes express CYP3A protein in biopsies from older patients. A, the arrow shows the enterocyte not expressing CYP3A (biopsy from a 48-day-old patient). B, all enterocytes express CYP3A (biopsy from a 10-year-old patient). P-gp is located on the apical and lateral surfaces of the enterocytes. C, apical staining of P-gp (biopsy from a 7-year-old patient). D, lateral staining of P-gp (biopsy from a 3-month-old patient).
To get insight into the impact of age on the CYP3A and P-gp mRNA expression, our population was arbitrarily divided into three groups: group 1, patients aged 1 month to 1 year (n = 19); group 2, 1 to 6 years (n = 16); and group 3, patients over 6 years of age (n = 24). 18S and villin mRNA levels were not different among the three groups (data not shown). The high interindividual variability in the expression found for the CYP3A isoforms as well as for P-gp was highlighted by using percentiles as shown in Fig. 2. The dispersion was higher in group 1 (1 month to 1 year) when compared with groups 2 and 3 for CYP3A4 and CYP3A5.
Discussion
CYP3A and P-gp are present in many tissues, but primarily in the liver and gut. However, very few studies investigated the postnatal development of the CYP3A/P-gp system in humans, since tissues from neonates, infants, and children are difficult to collect. In the present study, the duodenal biopsies were taken for medical investigations. The very small amount of tissue did not allow us to determine specific protein content or activity, but immunohistochemical localization and mRNA quantification were performed.
For CYP3A, we observed a staining heterogeneity between adjacent enterocytes of the same villus in patients aged less than 6 months, whereas the protein was uniformly expressed in older biopsies. A heterogeneous expression of CYP3A protein had been previously reported in hepatic biopsies from children older than 6 months and adults (Ratanasavanh et al., 1991). These results suggest that the developmental profile of CYP3A might be cell dependent. The P-gp protein localization was reported to be apical but not lateral in the rare available data obtained in adults using “MRK16” antibody (Thiebaut et al., 1987). Since an experimental artifact was ruled out by testing C494 antibody on adult liver and intestine biopsies, this new undescribed lateral P-gp localization reported in our study might be the result of a developmental process.

Ratio box-plots and percentile tables of mRNA copies/villin mRNA copies for A, CYP3A4; B, CYP3A5; C, CYP3A7; and D, P-gp.
To quantify the mRNA, we developed a very specific and sensitive TaqMan method that showed no cross-amplification among the three CYP3A isoforms. Our mRNA quantification results, normalized to villin, were in the same order of magnitude as those reported by Nishimura et al. (2003) on a pooled small intestinal total RNA.
The CYP3A protein content in the intestine was reported in the literature to gradually increase in the first months of life (Johnson et al., 2001), which is not consistent with the mRNA level decrease reported in our study. This dissociation between mRNA and protein contents during the maturation process was already reported for CYP2D6 (Treluyer et al., 1991). This might reflect a post-transcriptional regulatory mechanism, with the possible implication of a local intestinal trigger factor, in the activation of translation. We were able to detect CYP3A7 mRNA in 64% of our duodenal samples. Its level of expression was much lower than that of CYP3A4 and did not vary with age. According to our data, the contribution of CYP3A7 in the overall CYP3A-dependent enterocytic metabolism is very limited in children. The interindividual variability in CYP3A5 mRNA expression was important, with high expression in a limited number of samples. The pharmacogenetically dependent expression of CYP3A5 was not investigated in the present study, although the high levels of CYP3A5 protein expression were previously associated with a genetic polymorphism in intron 3 of CYP3A5 (Hustert et al., 2001; Kuehl et al., 2001). Many studies described the hepatic ontogeny of CYP3A isoforms. CYP3A7, detected in fetal liver, was replaced by increasing protein and activity levels of CYP3A4 after birth (Lacroix et al., 1997; Kearns et al., 2003). In contrast, in our duodenal samples, after birth, CYP3A4 mRNA expression decreased significantly after the first month of life to reach adult levels. This discrepancy in the expression profile of the three isoforms between the liver and the intestine was already observed in rats (Johnson et al., 2000) and suggest that an organ-dependent factor might control the expression of the different isoenzymes. Additional studies are required to confirm these findings and to obtain additional data from other duodenal/jejunal segments of the intestine.
In contrast to CYP3A isoform expression, the ontogenic profile of P-gp mRNA and protein content are in agreement. In the literature, in humans, P-gp mRNA and protein were detected as early as 11 to 14 weeks' gestation in the liver and the kidney but were only detected in the intestine in older samples (van Kalken et al., 1992). Similarly, P-gp mRNA was present in all our samples, from 1 month until adulthood, with a large interindividual variability. The dissociation between the CYP3A mRNA level reported in the present study and protein content and activity found in the literature highlights the possibility of a post-transcriptional regulation which should be supported by additional research. In addition, in the intestine, food and orally administered xenobiotics might trigger CYP3A4 and CYP3A5 protein synthesis in the first months of life, playing a protective role against xenobiotic exposure. Additional studies would allow us to determine whether the local ontogeny of drug-metabolizing enzymes and transporters in the intestine might depend on the type of diet and the time of diet diversification.
Acknowledgments
We thank Professor Philippe Beaune, Dr. Veronique Barbu, and Dr. Thierry Cresteil for providing antibodies and/or plasmids, and Regine Paris and Agnes Florentin for technical assistance in the immunohistochemistry studies.
Footnotes
-
This work was supported by the Center for Biological Investigations (CIB), Hôpital Robert Debré; Assistance Publique, Hôpitaux de Paris, France; and the Conseil de la Recherche, Saint Joseph University, Beirut, Lebanon. This work was presented as a poster at the Annual Meeting of the American Society of Clinical Pharmacology and Therapeutics and published. (Clin Pharmacol Ther75:P50, 2004).
-
Present address (C.L.): Sainte Justine Hospital, Montreal, Canada.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.105.005611.
-
ABBREVIATIONS: P-gp, P-glycoprotein; RT, reverse transcription; PCR, polymerase chain reaction.
- Received May 20, 2005.
- Accepted July 26, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}