


Abstract
In addition to the many articles reporting on the marked differences in species and large differences in rat strains in response to aldehyde oxidase (AO), individual differences in some rat strains have also been reported. However, little has been clarified about any related molecular biological mechanisms. We previously revealed that nucleotide substitutions of 377G>A and 2604C>T in the AO gene might be responsible for individual differences in AO activity in Donryu strain rats. By using native polyacrylamide gel electrophoresis/Western blotting in this study, the lack of formation of the AO dimer protein, which is essential for catalytic activity, was shown in poor metabolizer Donryu rats, and this could be a major reason for the individual differences. Rat strain differences were also verified from the same perspectives of nucleotide substitutions and expression levels of a dimer protein. Rat strains with high AO activity showed nucleotide sequences of (377G, 2604C) and a dimer protein. In the case of those with low AO activity, the nucleotide at position 2604 was fixed at T, but varied at position 377, such as G, G/A, and A. An AO dimer was detected in the liver cytosols of rat strains with (377G, 2604T), whereas a monomer was observed in those with (377A, 2604T). These results suggest that the lack of formation of a dimer protein leading to loss of catalytic activity might be due to 377G>A nucleotide substitution. Individual and strain differences in AO activity in rats could be explained by this 377G>A substitution, at least in the rat strains used in this study.
Both aldehyde oxidase (AO) and xanthine oxidase are major members of the molybdenum hydroxylase family. They consist of a homodimer with a subunit molecular mass of about 150 kDa. Each subunit of AO contains a molybdopterin cofactor, a FAD, and two different 2Fe-2S redox centers that are essential for catalytic activity. AO catalyzes the oxidation of a wide range of endogenous and exogenous aldehydes and N-heterocyclic aromatic compounds. N-heterocyclic drugs such as methotrexate, 6-mercaptopurine, cinchona alkaloids, and famciclovir are oxidized by this enzyme (Beedham 1985, 1987, 1997, 2002; Kitamura et al., 2006). In addition, aldehyde oxidase can catalyze in vitro reduction of a variety of functional groups including sulfoxides, N-oxides, azo dyes, and N-hydroxycarbamoyl substituents in the presence of an appropriate donor (Kitamura et al., 2006). In fact, the atypical antipsychotic drug ziprasidone is chiefly metabolized to its reductive ring-cleaved S-methyldihydroziprasidone by AO after administration in humans (Prakash et al., 1997; Beedham et al., 2003).
Marked species differences have been well documented for the AO-catalyzed metabolism of drugs including methotrexate (Jordan et al., 1999; Kitamura et al., 1999b) and famciclovir (Rashidi et al., 1997). A large variation in rat strains has also been demonstrated in the oxidation activity of benzaldehyde (Sugihara et al., 1995) and methotrexate (Kitamura et al., 1999a). An interesting individual difference has been reported in Wistar rats (Gluecksohn-Waelsch et al., 1967) and in Sprague-Dawley (SD) rats (Beedham et al., 1998). Similar to those reports, we observed remarkable species differences (Takasaki et al., 2005; Itoh et al., 2006), large differences in rat strains (Sasaki et al., 2006), and striking individual differences in Donryu rats (Itoh et al., 2007b) in the AO-catalyzed oxidation pathway of the (S)-enantiomer of RS-8359 [(±)-4-(4-cyanoanilino)-5,6-dihydro-7-hydroxy-7H-cyclopenta[d]-pyrimidine] (Itoh et al., 2007b). The compound is a reversible and selective MAO-A inhibitor (Yokoyama et al., 1989; Miura et al., 1993) and has been developed as an antide-pressant (Plenker et al., 1997; Puchler et al., 1997). One of the major metabolic pathways of RS-8359 is the AO-catalyzed 2-oxidation on the pyrimidine ring to give the 2-keto metabolite (Itoh et al., 2005).
The AO-catalyzed 2-oxidation activity of (S)-RS-8359 in Donryu strain rats was shown to be divided into three groups: ultrarapid metabolizer (UM), extensive metabolizer (EM), and poor metabolizer (PM). Nucleotide substitutions at positions 377 and 2604 were suggested to be primarily responsible for differences in aldehyde oxidase activity observed in that strain. In particular, 377G>A substitution was supposed to be more central for the deficiency of the catalytic activity of AO than 2604C>T substitution, because the former substitution accompanies an amino acid change from a symmetric Gly to an asymmetric Ser very close to the second Fe-S redox center (Itoh et al., 2007b). In another report, we demonstrated that rat strains with high AO activity exhibited a dimeric protein on native PAGE/Western blot in contrast to rat strains with low AO activity, which produced only a monomer protein (Itoh et al., 2007a). The results clearly showed that discussions on rat strain differences in AO activity should be based on the expression levels of the dimeric protein itself. From a combined consideration of the results obtained from the two separately conducted studies on individual differences in Donryu rats and differences in rat strains, we assumed that individual differences in Donryu rats might be due to a deficiency in formation of the AO dimeric protein. In this study, we investigated the possibility that 377G>A nucleotide substitution in the AO gene might result in the differences in rat strains.
Materials and Methods
Chemicals. The (S)-enantiomer and 2-keto metabolite of RS-8359 were supplied by Ube Kosan Co. Ltd. (Yamaguchi, Japan). Hydrocortisone, an internal standard of HPLC analysis, was purchased from Sigma Chemical Co. (St. Louis, MO). All other reagents were of reagent grade.
Preparation of Liver Cytosolic Fractions. Eight-week-old male rats of 10 strains were used. Crj:Donryu strain rats were purchased from Charles River Japan (Yokohama, Japan). Wistar strain rats were obtained from Charles River Japan (Crlj:Wistar and LEW/CrlCrlj), Japan Slc (Shizuoka, Japan; Slc:Wistar, WKY/Izm, and WKAH/Hkm), Clea Japan (Tokyo, Japan; Jcl:Wistar), and Imamichi Institute for Animal Reproductions (Saitama, Japan; Iar:Wistar or Wistar-Imamichi). Sprague-Dawley strain rats were obtained from Charles River Japan (Crl:SD) and Japan Slc (Slc:SD). Fisher strain rats (F344/DuCrl-Crlj) were purchased from Charles River Japan. F344/DuCrlCrlj, LEW/Crl-Crlj, WKY/Izm, and WKAH/Hkm are inbred; the others are a closed colony. The animals were housed according to the Guidelines for Animal Experimentation (Tohoku Pharmaceutical University) in cages in rooms with a unidirectional airflow at a controlled temperature (22 ± 2°C), relative humidity (50 ± 10%), and 12-h light/dark cycles (7:00 AM—7:00 PM). Tap water was available ad libitum and CE2 food (Clea Japan) was available ad libitum except for overnight fasting before use. The animals were sacrificed by bleeding from the carotid artery under anesthesia and their livers were immediately extracted. The livers were homogenized in 3 volumes of 10 mM phosphate buffer (pH 7.4) containing 1.15% KCl and 100 μM phenylmeth-anesulfonyl fluoride by a Potter-Elvehjem Teflon homogenizer. The cytosolic fractions were prepared by successive centrifugation at 9000g for 20 min and then at 105,000g for 60 min. Protein concentration was determined using the Protein Quantification Kit-Rapid (Dojindo Laboratories, Kumamoto, Japan) with bovine serum albumin as the standard.
Enzyme Activity Assay. The (S)-enantiomer of RS-8359 (100 μM) was incubated at 37°C for 30 min in a reaction mixture (0.25 ml) consisting of 100 mM phosphate buffer (pH 6.0), 1.0 mM K3Fe(CN)6, 0.13 mM EDTA, and prepared cytosol (20 mg/ml, 0.10 ml). The reaction was stopped by the addition of acetonitrile (0.50 ml) containing 0.2 mg/ml hydrocortisone as an internal standard, after which the mixture was centrifuged at 10,000g for 3 min. Aliquots (25 μl) of the supernatant were analyzed for quantification of the oxidation product by reversed-phase HPLC on a YMC ODS A-312 column (6.0 mm i.d. × 150 mm; YMC Co. Ltd., Kyoto, Japan). A mobile phase was composed of acetonitrile/0.5% ammonium acetate (25:75); the flow rate was 1.0 ml/min. The HPLC instrument was a Shimadzu model 6A High Performance Liquid Chromatograph System (Shimadzu Seisakusho Co. Ltd., Kyoto, Japan). The peaks were monitored for absorbance at 315 nm, and the peak area was calculated on a Chromatopac C-R4A (Shimadzu Seisakusho).
Preparation of Anti-Monkey Liver AO Antisera. The rabbit anti-monkey AO antisera used in this study were prepared as in the previous report (Hoshino et al., 2007).
Native PAGE/Western Blot Analysis. Cytosolic proteins were separated by native PAGE, which was performed on PhastGel gradient 8-25 in PhastGel SDS buffer strips (GE Healthcare Ltd. UK, Buckinghamshire, UK). The proteins were transferred electrophoretically to a polyvinylidene difluoride (PVDF) membrane (Daiichi Pure Chemicals Co. Ltd., Tokyo, Japan) in transfer buffer (15% methanol containing 25 mM Tris and 192 mM glycine, pH 8.3). AO was detected by using an ECF Western Blotting Kit (GE Healthcare). The membrane was blocked with 5% nonfat dry milk in phosphate-buffered saline and then successively incubated with a primary rabbit anti-monkey AO antibody at 0.5 μg/ml, a secondary antibody (anti-rabbit fluorescein-linked whole antibody) at a dilution of 1:600, and a tertiary antibody (anti-fluorescein alkaline phosphatase conjugate) at a dilution of 1:2500. The blocking and incubation at each immunoreaction step were performed at room temperature for 1 h, and the membrane was washed two or three times with phosphate-buffered saline containing 0.1% Tween 20. The target proteins on the membrane were detected by the enhanced chemifluorescence detection system (GE Healthcare). An HMW Calibration Kit (GE Healthcare) was used for the molecular weight standards.
Gel Permeation Chromatography of UM and PM Donryu Rat AO. The rats were divided into the UM and PM groups by measuring the AO activity of liver cytosol. AO was purified from each group basically according to the method described previously (Itoh et al., 2005), except for the heat treatment at 60°C for 10 min and the use of gel permeation chromatography instead of hydroxyapatite chromatography. In brief, ammonium sulfate was added to cytosol to 50% saturation. The protein precipitates were dissolved in 10 mM phosphate buffer (pH 7.4) and applied to a Benzamidine Sepharose 6B column (3 × 22 cm; GE Healthcare). Then, AO was purified by a HiLoad Superdex 200 pg column (2.6 × 60 cm; GE Healthcare) chromatography, which was eluted with 50 mM phosphate buffer (pH 7.2) containing 0.15 M NaCl. The elutes were concentrated using Viva Spin 6 (Vivascience, New York, NY) for measurement of AO activity. A Gel Filtration Calibration Kit (GE Healthcare) was used to estimate the molecular weight. Standard proteins are ferritin (mol. wt. 440,000), aldolase (mol. wt. 158,000), conalbumin (mol. wt. 75,000), and ovalbumin (mol. wt. 43,000).
Direct Sequence Analysis of Genomic DNA. Genomic DNA was isolated with a DNeasy Blood and Tissue Kit (QIAGEN, Hilden, Germany) according to the manufacturer's instructions. Primer sets were designed in reference to the nucleotide sequences of rat (GenBank accession number NW 047816) on introns as follows: 377 forward: 5′-gaggtcttctctccaaattcagtg-3′, 377 reverse: 5′-ggtaggtgtccaattattactgtca-3′, 2604 forward: 5′-cttcccaatatcactaaaccagcta-3′, 2604 reverse: 5′-gcagtgagccctttattagatgtta-3′.
The PCR was conducted in a solution of 50 μl containing 1× Ex-Taq buffer, 200 μM dNTP mix solution, 2.5 U Ex-TaqDNA polymerase (TaKaRa Bio Inc., Shiga, Japan), 2 mM MgCl2, 0.2 μM concentration of forward and reverse primer, and 2 μl of the template. PCR amplification was conducted using a PCR thermal cycler MP (TaKaRa) as follows: initial denaturation at 94°C for 5 min, and 30 cycles of 94°C for 30 s, 60°C for 30 s, 72°C for 30 s, and a final extension at 72°C for 10 min. The PCR products obtained were purified using the Wizard SV Gel and PCR Clean-Up System (Promega, Madison, WI). The DNA sequences were determined using a CEQ 8000 Genetic Analysis System (Beckman Coulter Inc., Fullerton, CA) and a DTCS Quick Start Kit, for which the above-mentioned forward or reverse primer was used as the direct sequence primer.
Statistical Analysis. The results are expressed as the mean ± S.E. for the number of experiments. Statistical significance was compared between low and high activity groups by Student's t test. Values with p < 0.05 were considered statistically significant.
Results
Native PAGE/Western Blot Analysis of AO in Donryu Strain Rats. Native PAGE/Western blot analysis was conducted on the liver cytosol of Donryu rats. That strain demonstrated polymorphism in AO such as UM in those with nucleotide sequences of (377G, 2604C), EM in those with (377G/A, 2604C/T), and PM in those with (377A, 2604T) (Itoh et al., 2007b). Figure 1 clearly shows that UM, EM, and PM contained 300 kDa of dimer protein, a mixture of both dimer and monomer protein, and only 150 kDa of monomer protein, respectively. Gel permeation chromatography of the partially purified AO revealed that the UM rat AO eluted at 300 kDa as an active enzyme (shown by an arrow in Fig. 2a), whereas the PM rat AO eluted at 150 kDa as a nonactive enzyme (shown by an arrow in Fig. 2b).
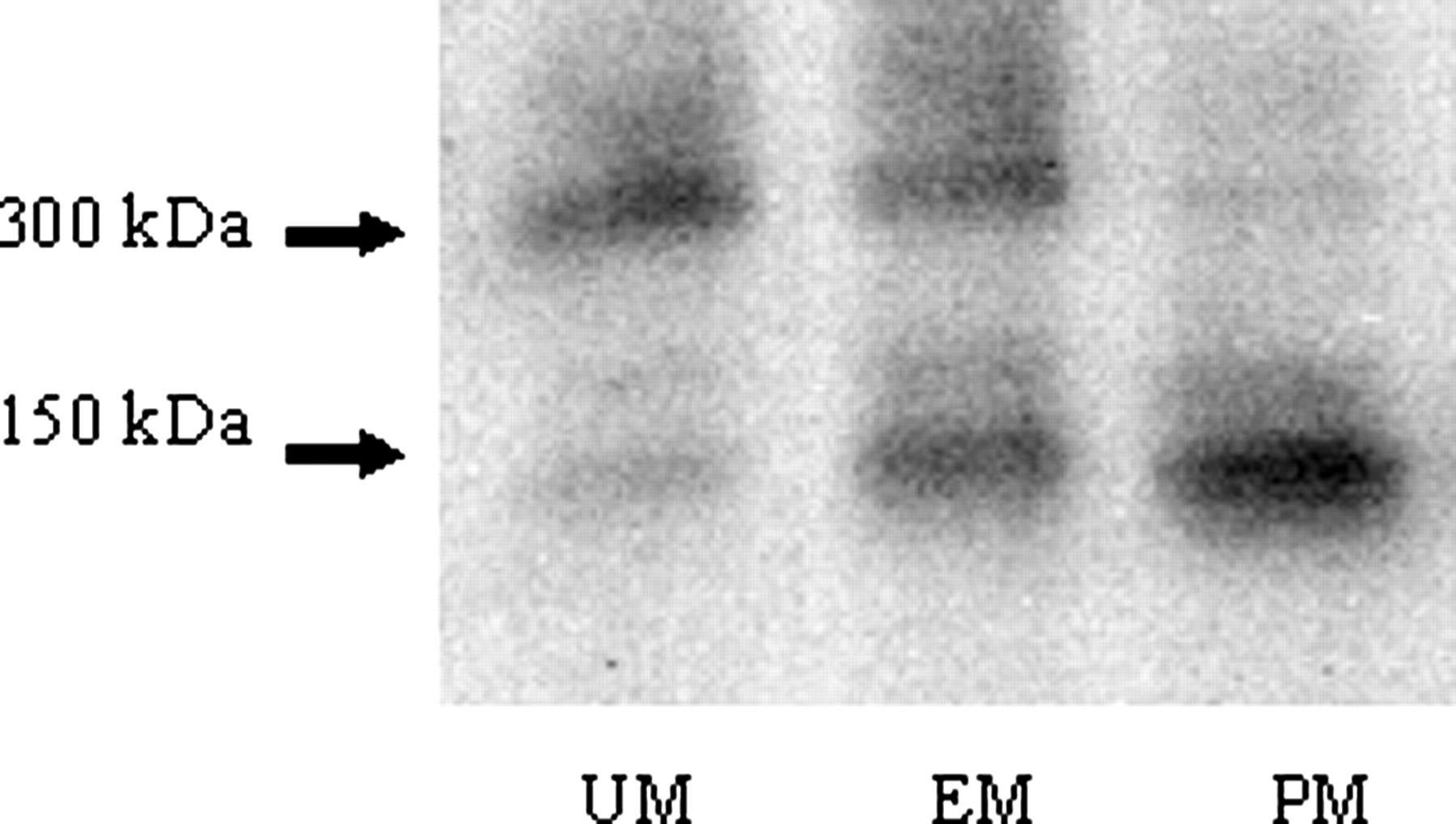
Native PAGE/Western blot analysis of AO protein in liver cytosols of Donryu strain rats. Cytosolic proteins were separated by native PAGE, transferred electrophoretically to a PVDF membrane, and immunoreacted against rabbit antimonkey AO antibody. The target proteins on the membrane were detected by the enhanced chemifluorescence detection system.
Genomic DNA Analysis. As the relationship between nucleotide substitutions and dimer formation was revealed for AO activity in the Donryu rat strains, the genomic DNA of the livers from the 10 rat strains was analyzed by the direct sequence method. The nucleotide sequences were (377G, 2604C) in the high AO activity group of the WKAH/Hkm, WKY/Izm, LEW/Crlj, Jcl:Wistar, and Wistar-Imamichi strains. In contrast, they were (377A, 2604T) in the low AO activity group of the Slc:Wistar and F344/DuCrlCrlj strains (Table 1). The three individual rats in these seven strains showed completely identical nucleotide sequences. However, Crlj:Wistar rats showed three different types of nucleotide sequences, (377G, 2604C), (377G/A, 2604C/T), and (377A, 2604T), which were the same as those of the Donryu strain (Itoh et al., 2007b). In addition, the two SD strains of Crl:SD and Slc:SD had other sequence patterns, (377G, 2604T), (377A, 2604T), and (377G/A, 2604T), thus being heterozygous at position 377, but homozygous at position 2604.
Nucleotide substitutions of AO genomic DNA in seven rat strains

Gel permeation chromatography showing the dimeric nature of UM rat AO and the monomeric nature of PM rat AO. The chromatography of partially purified AO was performed on a HiLoad Superdex 200-pg column. The molecular mass of AO was calculated by comparing its elution volume with those of standard proteins. The UM rat AO eluted at 300 kDa as an active enzyme (shown by an arrow in Fig. 2a), whereas the PM rat AO eluted at 150 kDa as a nonactive enzyme (shown by an arrow in Fig. 2b). Molecular mass markers in kilodaltons are 440 for ferritin, 158 for aldolase, 75 for conalbumin, and 43 for ovalbumin, respectively.
Characteristics of AO in SD Strain Rats. Because new combinations of nucleotide substitutions were found in the AO of SD strain rats, detailed analyses of nucleotide sequences, enzyme activities, and immunoreactive 300-kDa expression levels were performed using 14 Crlj:SD strain rats. The nucleotide sequences (Table 2) were divided into three groups: group 1 (n = 3; 377G, 2604T), group 2 (n = 5; 377G/A, 2604T), and group 3 (n = 6; 377A, 2604T). Group 1 exhibited nearly the same activity as that of group 2. In contrast, group 3 showed significantly lower activity (Fig. 3). The presence of AO dimer protein was confirmed in group 1 but not in group 3, in which only a monomer protein was detected; both the dimer and monomer proteins were present in group 2 according to the respective native PAGE/Western blot analyses (Fig. 4). Almost the same results were obtained in the experiment using 13 Slc:SD strain rats (Table 2).
Nucleotide substitutions of AO genomic DNA in Crl:SD and Slc:SD rat strains
Both Crl:SD and Slc:SD strains belong to the low AO activity group. The number in each group is indicated in parentheses.
Discussion
In addition to marked species differences and large variations in rat strains (Beedham et al., 2002; Kitamura et al., 2006), individual differences have been reported in AO activity in Wistar rats (Gluecksohn-Waelsch et al., 1967), SD rats (Beedham et al., 1998), and Donryu rats (Itoh et al., 2007b). However, little has been known about the mechanisms contributing to the differences. In the previous report (Itoh et al., 2007b), we demonstrated polymorphism in the AO of the Donryu rat strain such that UM, EM, and PM possess nucleotide substitutions of (377G, 2604C), (377G/A, 2604C/T), and (377A, 2604T), respectively. The current study revealed that this polymorphism was caused by a lack of dimer formation necessary for onset of catalytic activity. Only two nucleotides among 4032 base pairs were different between UM and PM in Donryu strain rats, suggesting that the two nucleotide substitutions are very critical for dimer formation. The nucleotide substitutions of 377G>A and 2604C>T result in the amino acid changes of Gly110Ser and Ala852Val, respectively (Itoh et al., 2007b). The results suggest that changing from a symmetric Gly to an asymmetric Ser in a position very near the second Fe-S redox center (Wright et al., 1999) might cause a more serious conformational change in AO protein structure and loss of enzyme activity than would the amino acid change of Ala852Val.
The above-mentioned mechanism of individual differences in AO activity in Donryu strain rats, i.e., a genetic polymorphism, was also postulated to be the case for the difference in AO activity among rat strains. The postulate was verified with direct sequence analysis of genomic DNA and native PAGE/Western blot analysis of AO dimer protein in 10 rat strains. However, the situations were considerably more complex for the differences in rat strains than for the individual differences in Donryu rats. Rat strains with high AO activity showed simple nucleotide sequences of (377G, 2604C) and a dimer protein, whereas among rat strains with low activity, Slc:Wistar and F344/DuCrlCrlj rats had different types of nucleotide substitutions of (377A, 2604T) and a monomer protein. In contrast, Crl:SD and Slc:SD rats possessed more complex sequences of (377G, 2604T), (377G/A, 2604T), and (377A, 2604T) and a mixture of a dimer and a monomer protein. Furthermore, high activity Crlj:Wistar rats exhibited the same three nucleotide substitutions and protein profiles as those of Donryu rats, suggesting that Crlj:Wistar rats will exhibit polymorphism in AO activity. As shown previously (Itoh et al., 2007a), the larger variation seen in the activities and expression levels of AO for Crlj:Wistar than those for other high activity rat strains might be ascribed to polymorphic metabolism. Thus, the nucleotide substitutions at positions 377 and 2604 were obviously responsible not only for the individual differences in the Donryu strain rats, but also for the rat strain differences in AO activity.

AO-catalyzed 2-oxidation activity of (S)-RS-8359 in liver cytosols of Crl:SD strain rats. The (S)-enantiomer of RS-8359 (200 μM) was incubated at 37°C for 10 min with cytosol (20 mg/ml, 0.10 ml) and the oxidation product was quantified by reversed-phase HPLC.
The complex nucleotide substitutions at positions 377 and 2604 were observed in both Crl:SD and Slc:SD rats. In those cases, the nucleotide at 2604 was fixed to T, but was varied at 377; for example, as G, G/A, and A. A 300-kDa dimer protein showing high AO activity was found in rats with the nucleotide substitutions of (377G, 2604T). In contrast, only a 150-kDa monomer protein showing low AO activity was found in rats with substitutions of (377A, 2604T). The mechanism of individual differences in the AO activity of Donryu strain rats was presumed to be the 377G>A substitution among 4032 base pairs, as mentioned above. Judging from the results obtained in the SD strain rats and the possible mechanism of individual differences in the Donryu strain rats, it is suggested that the 377G>A substitution might be the major cause of the lack of dimer formation of AO. It is possible that the nucleotide substitution of 2604C>T is associated with the dimer formation of AO because of the relatively low expression level of the 300-kDa protein in rats with the nucleotide substitutions of (377A, 2604T). This will be an interesting subject for future clarification.

Native PAGE/Western blot analysis of AO protein in liver cytosols of Crl:SD strain rats. Cytosolic proteins were separated by native PAGE, transferred electrophoretically to a PVDF membrane, and immunoreacted against rabbit anti-monkey AO antibody. The target proteins on the membrane were detected by the enhanced chemifluorescence detection system.
In conclusion, the 377G>A nucleotide substitution in the AO gene accompanying amino acid change from a symmetric Gly to an asymmetric Ser at the 110 position was suggested as the reason for the lack of formation of AO dimer protein and the loss of its catalytic activity. The substitution might be primarily responsible not only for individual differences in AO of the Donryu and Crlj:Wistar strains of rats, but also for differences in AO activity in rat strains, at least in the rat strains used in this study.
Acknowledgments
We thank Dr. T. Ikeda, Director of the Drug Metabolism and Pharmacokinetics Research Laboratories, Sankyo Co. Ltd., and Drs. K. Nishimura and Y. Kawahara, Ex-directors of the Research Laboratories, for kind encouragement.
Footnotes
-
doi:10.1124/dmd.107.015503.
-
ABBREVIATIONS: AO, aldehyde oxidase; SD, Sprague-Dawley; RS-8359, (±)-4-(4-cyanoanilino)-5,6-dihydro-7-hydroxy-7H-cyclopenta[d]-pyrimidine; MAO-A, monoamine oxidase A; PM, poor metabolizer; EM, extensive metabolizer; UM, ultrarapid metabolizer; PCR, polymerase chain reaction; native PAGE, native polyacrylamide gel electrophoresis; HPLC, high-performance liquid chromatography; PVDF, polyvinylidene difluoride.
- Received March 1, 2007.
- Accepted July 12, 2007.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}