


Abstract
Alterations in transporter expression may represent a compensatory mechanism of damaged hepatocytes to reduce accumulation of potentially toxic compounds. The present study was conducted to investigate the expression of hepatobiliary efflux transporters in livers from patients after toxic acetaminophen (APAP) ingestion, with livers from patients with primary biliary cirrhosis (PBC) serving as positive controls. mRNA and protein expression of multidrug resistance-associated protein (MRP) 1-6, multidrug resistance protein (MDR) 1-3/P-glycoprotein (P-gp), and breast cancer resistance protein (BCRP) in normal (n = 6), APAP overdose (n = 5), and PBC (n = 6) human liver samples were determined by branched DNA and Western blot analysis, respectively. Double immunohistochemical staining of P-gp and proliferating cell nuclear antigen (PCNA), a marker of proliferation, was performed on paraffin-embedded tissue sections. Compared with normal liver specimens, MRP1 and MRP4 mRNA levels were elevated after APAP overdose and in PBC. Up-regulation of MRP5, MDR1, and BCRP mRNA occurred in PBC livers. Protein levels of MRP4, MRP5, BCRP, and P-gp were increased in both disease states, with MRP1 and MRP3 protein also being induced in PBC. Increased P-gp protein was confirmed immunohistochemically and was found to localize to areas of PCNA-positive hepatocytes, which were detected in APAP overdose and PBC livers. The findings from this study demonstrate that hepatic efflux transporter expression is up-regulated in cases of APAP-induced liver failure and PBC. This adaptation may aid in reducing retention of byproducts of cellular injury and bile constituents within hepatocytes. The close proximity of P-gp and PCNA-positive hepatocytes during liver injury suggests that along with cell regeneration, increased efflux transporter expression is a critical response to hepatic damage to protect the liver from additional insult.
The incidence of acetaminophen (APAP) overdose, which frequently leads to severe liver injury at supratherapeutic doses, has been steadily increasing in the United States. The number of APAP-related cases of liver failure has increased from 39% (1998-2001) to nearly 50% (2003) of all cases of acute liver failure, with well over 100,000 cases of APAP overdose reported annually in the United States (Lee, 2004). Although APAP poisoning is often intentional, an increased incidence of unintentional overdose cases is gaining attention. A recent study reported that 50% of APAP overdose cases were attributable to unintentional exposure, most commonly because of APAP overutilization in multiple over-the-counter and prescription preparations (Larson et al., 2005).
Mouse models of APAP-induced hepatotoxicity are widely used to recapitulate human exposures because of the similarities in APAP bioactivation and patterns of liver injury between mice and humans. Similar doses of APAP cause hepatotoxicity in both species, and the liver injury is dose-dependant (Mitchell et al., 1973; Davis et al., 1974). Another striking similarity between the two species is the development of resistance to toxic doses of APAP through prior exposure to subtoxic APAP doses. Several studies in rodents demonstrated that repeated exposure to subtoxic doses of APAP protects against a subsequent lethal dose, a phenomenon known as autoprotection (Strubelt et al., 1979; Shayiq et al., 1999). In humans, acquired resistance to APAP has been described in individual case reports, such as that described by Shayiq et al. (1999). Similar findings in both rodents and humans indicate that many adaptive responses to hepatotoxicity occur.
Whereas the development of resistance to APAP probably involves multiple compensatory changes, altered expression of hepatobiliary efflux transporters may be critical to achieve this resistance. We have previously reported increased mRNA and protein expression of multidrug resistance-associated proteins 2-4 (Mrp2-4) in mouse liver after exposure to a hepatotoxic dose of APAP (Aleksunes et al., 2005, 2006b). This increased expression of efflux transporters may help to prevent the accumulation of potentially toxic compounds within hepatocytes, thus contributing to acquired resistance. To determine whether similar transporter changes occur in humans after toxic APAP exposure, we investigated the gene and protein expression of nine major hepatic efflux transporters in the livers of APAP overdose patients compared with normal human livers. Transporter expression in the livers of patients with primary biliary cirrhosis (PBC) was also examined as a positive control because of the well characterized changes in expression of some transporters in this disease state (Kojima et al., 2003; Zollner et al., 2003). Furthermore, we investigated whether changes in efflux transporter expression were localized to areas of hepatocyte replication, as compensatory proliferation has been implicated as an important part of autoprotection (Mehendale et al., 1994; Shayiq et al., 1999).
Materials and Methods
Human Liver Specimens. Archival samples of frozen and formalin-fixed, paraffin-embedded adult explant livers (normal: n = 6; PBC: n = 6; APAP overdose: n = 5) were obtained from the Liver Tissue Procurement and Distribution System at the University of Minnesota. Diagnosis was first established by the University of Minnesota and confirmed by histological examination at the University of Connecticut in a blinded fashion. Sections were photographed on an Olympus BX50 microscope equipped with a QImaging MicroPublisher 3.3 RTV camera. Images were acquired with QCapture Pro software. Medical history available for the patients was limited; however, their basic characteristics are described in Table 1.
Characteristics of patients
RNA Extraction. RNAzol B reagent (Tel-Test Inc., Friendswood, TX) was used to extract total liver RNA according to the manufacturer's protocol. Integrity of the RNA samples was confirmed by agarose gel electrophoresis and ethidium bromide staining.
Branched DNA Signal Amplification Assay. Human MRP1, 2, 3, 4, 5, 6 (ABCC1-6), multidrug resistance protein (MDR) 1 and 3 (ABCB1 and ABCB4, respectively), breast cancer resistance protein (BCRP) (ABCG2), and bile salt export pump (BSEP) (ABCB11) mRNA were measured using the bDNA signal amplification assay (Quantigene High Volume bDNA Signal Amplification Kit; Panomics, Inc., Fremont, CA) as described previously (Aleksunes et al., 2005). Probe sets for MRP1-5, MDR1, MDR3, BCRP, and BSEP were obtained from Panomics. The human MRP6 gene sequence was acquired from GenBank. Oligonucleotide probe sets against MRP6 were designed using Probe Designer software (version 1.0; Bayer Corp., Emeryville, CA) (Table 2). Ten microliters of total RNA (1 μg/μl) were added to each well of a 96-well plate containing capture hybridization buffer and 100 μl of each diluted probe set for all transporters. The RNA was allowed to hybridize to probe sets overnight at 53°C. The subsequent hybridization steps were performed according to the manufacturer's protocol. Luminescence was measured with a Quantiplex 320 bDNA Luminometer interfaced with Quantiplex Data Management Software version 5.02. The luminescence for each well is reported as relative light units per 10 μg of total RNA.
Oligonucleotide probes generated for analysis of human MRP6 gene expression by bDNA signal amplification assay
Preparation of Crude Membrane Fractions. Liver membrane fractions were prepared as described previously (Aleksunes et al., 2006b). The resulting pellet of the crude membrane fraction was resuspended in sucrose-Tris buffer. Protein concentration was determined using Bio-Rad protein assay reagents (Bio-Rad, Hercules, CA).
Western Blot Analysis. Membrane proteins were electrophoretically resolved using polyacrylamide gels and transblotted overnight at 4°C onto PVDF-Plus membrane (GE Osmonics, Minnetonka, MN). Immunochemical detection of MRP1 (∼190 kDa), MRP2 (∼190-200 kDa), MRP3 (∼180-190 kDa), MRP4 (∼160-170 kDa), MRP5 (∼160-180 kDa), MRP6 (∼165 kDa), BCRP (∼75 kDa), and P-glycoprotein (P-gp) (∼170 kDa) proteins was performed as described by Aleksunes et al. (2006b) using MRPr1, M2III-5, M3II-21, M4I-10, M5I-10, M6II-7, BXP-53, and C219 antibodies, respectively. Anti-MRP and BCRP antibodies were provided by G.L.S., and the P-gp antibody was obtained from Abcam Inc. (Cambridge, MA). Equal protein loading was confirmed using β-actin (ab8227; Abcam Inc.) as a loading control. Protein detection and semiquantitation of band intensities was performed as described previously (Aleksunes et al., 2006b).
Immunohistochemistry. Sections of formalin-fixed, paraffin-embedded livers were deparaffinized in xylene and then rehydrated through a graded alcohol series. Endogenous peroxidase activity was blocked with 3% H2O2 for 10 min, and antigen retrieval was performed by incubating slides in citrate buffer (10 mM) at 95°C for 10 min. For double-immunohistochemical staining of P-gp and PCNA, deparaffinized sections were incubated overnight with mouse anti-P-gp antibody diluted 1:100. Protein-antibody complexes were visualized using the mouse Vectastain Elite ABC kit and developed with 3,3′-diaminobenzidine according to the manufacturer's instructions (Vector Laboratories, Burlingame, CA). Sections were then counterstained for PCNA using the Zymed PCNA kit (Zymed Laboratories, South San Francisco, CA). PCNA immunostaining was visualized by developing with VIP peroxidase substrate (Vector Laboratories). Tissues were counterstained with methyl green, followed by dehydration in ethanol and clearing with xylene. Negative control staining was performed by incubating the sections without primary antibody. Antibodies used for Western blot immunostaining of MRP1-6 and BCRP were not suitable for immunohistochemical detection on paraffin sections in this study because of inconsistent staining patterns and/or lack of detection of the human proteins.
Statistical Analysis. mRNA data are presented as mean relative light units per 10 μg of total RNA ± S.E. For protein data, semiquantitative results are expressed as mean optical density ± S.E., and the data are normalized to control values (normal liver specimens). Data were analyzed using Student's t test with GraphPad Prism statistics software, comparing normal livers to either APAP or PBC livers. p < 0.05 was considered significant.
Results
Histopathology of Livers from Normal, APAP Overdose, and PBC Patients. Although limited data were provided regarding biochemical markers of liver injury in these patients (e.g., plasma alanine aminotransferase levels), hematoxylin and eosin-stained liver sections provided a histological indication of the degree of liver damage. Liver samples used as controls for transporter mRNA and protein analysis had no apparent lesions (Fig. 1A). Histological examination of APAP livers showed severe centrilobular necrosis and cell loss with lymphocytic accumulations and minimal biliary hyperplasia (Fig. 1B). Necrosis and fibrosis were present throughout the livers of PBC patients (Fig. 1C). Nodular regeneration and lymphocytic-histiocytic infiltration were also seen in these liver sections.

Histopathology of normal, APAP, and PBC livers. Formalin-fixed, paraffin-embedded liver sections from normal (A), APAP overdose (B), and PBC (C) patients were prepared and stained with hematoxylin and eosin. Images were taken at 10× magnification.
Efflux Transporter Gene Expression. Elevated MRP1 and MRP4 mRNA levels were seen in APAP (4.5- and 7.3-fold, respectively) and PBC (4.3- and 7.2-fold, respectively) livers (Fig. 2). Induction of MRP4 mRNA, along with increased MRP5 (5.7-fold) and BCRP (2.3-fold) mRNA levels in PBC has not been previously reported. mRNA expression of MDR1 was elevated 1.9-fold in PBC livers similar to published reports (Ros et al., 2003; Zollner et al., 2003). A trend toward increased MDR3 mRNA was noted in both APAP and PBC livers. Increased MDR1 and BCRP mRNA levels in APAP livers were also observed, but these increases were not significant, which may be due to individual variations. No changes in MRP2, 3, and 6 or BSEP (data not shown) mRNA were observed in APAP or PBC livers relative to normal expression.
Western Blot Analysis of Sinusoidal Efflux Transport Proteins. Immunoblot analysis was performed to determine whether increases in efflux transporter mRNA levels after APAP-induced hepatotoxicity lead to corresponding changes in protein expression. Elevated protein levels of sinusoidal efflux transporters were detected in livers of APAP overdose patients. Specifically, MRP4 and MRP5 proteins were up-regulated compared with those in normal liver (Fig. 3). Similarly, elevated levels of efflux transport proteins were detected in PBC livers. Increased expression of MRP1 and MRP3 protein was detected in PBC, as has been previously demonstrated (Ros et al., 2003; Zollner et al., 2003). An increase of MRP4 and MRP5 proteins was detected in livers of PBC patients. No significant changes in MRP6 protein expression were observed in APAP or PBC livers.
Western Blot Analysis of Canalicular Efflux Transport Proteins. APAP overdose resulted in enhanced protein expression of both BCRP and P-gp compared with that in normal livers (Fig. 4). Similarly, higher levels of these two canalicular efflux transporters were observed in PBC livers (BCRP and P-gp). The C219 P-gp antibody recognizes all members of the MDR family (Vos et al., 1998). No change in MRP2 protein expression was observed in either disease state.
Immunochemical Localization of P-gp and PCNA. Immunohistochemical staining was performed to assess the zonal distribution of P-gp expression in relation to regions of hepatocyte proliferation. Normal livers demonstrated a uniform expression pattern of P-gp protein, with no apparent zonal distribution throughout the liver lobule (Fig. 5A). There was minimal proliferation of hepatocytes in normal livers, as determined by PCNA staining. PCNA is a nuclear protein involved in DNA synthesis/cell cycle regulation, which increases in expression in early G1 and S phases of the cell cycle (Kurki et al., 1986). In APAP overdose, increased P-gp staining intensity was detected in midzonal and periportal hepatocytes adjacent to regions of centrilobular cell loss (Fig. 5B). The greater intensity of P-gp staining in APAP overdose compared with normal liver correlated with the increased protein expression detected by Western blot analysis. Double immunohistochemical staining for P-gp and PCNA revealed that the number of proliferating hepatocytes increased after APAP-induced acute liver failure and that induction of P-gp was localized to regions of hepatocyte proliferation (Fig. 5B). Similarly, in PBC liver sections both P-gp and PCNA immunostaining was increased compared with that in normal liver, primarily on the periphery of cirrhotic nodules, and enhanced P-gp staining was localized to areas also containing proliferating hepatocytes (Fig. 5C). Although elevated numbers of PCNA-positive hepatocytes were seen in both disease states, this was more dramatically increased in APAP livers.
Discussion
The present study investigates the expression of hepatobiliary efflux transporters during human liver injury caused by toxic drug exposure (APAP) or chronic disease (PBC). Up-regulation of several efflux transporters during PBC has been described in previous studies. However, this is the first time that elevated efflux transporter expression has been documented in cases of acute liver failure after ingestion of toxic doses of APAP. The results of this study demonstrate that mRNA and protein levels of hepatocellular efflux transporters increase during APAP-induced acute liver failure. In particular, hepatic MRP4, MRP5, P-gp, and BCRP proteins are increased after APAP overdose. Altered expression of hepatobiliary transporters in drug-induced acute liver failure and PBC indicates that these changes may be a general response of a damaged liver rather than a disease- or toxicant-specific effect. Induction of efflux transporters during PBC may limit accumulation of toxic biliary constituents within hepatocytes. These transporter changes may also prevent hepatocyte retention of drug metabolites and endogenous mediators of injury and inflammation generated during drug-induced hepatotoxicity. In addition, increased efflux transporter expression may facilitate the export of signaling molecules to adjacent hepatocytes and other cell types within the liver.
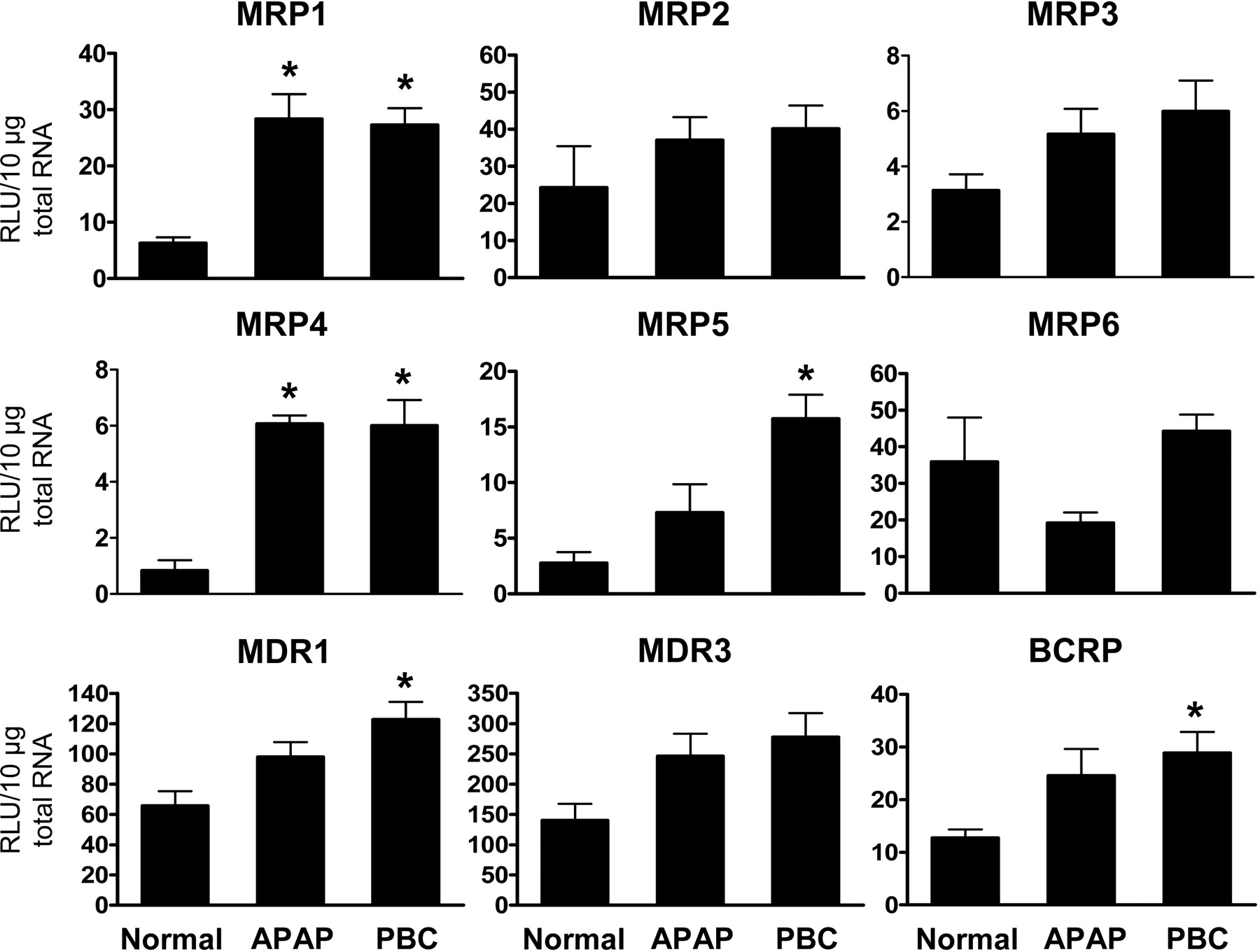
Hepatic expression of efflux transport genes. Total RNA was isolated from normal, APAP, and PBC human liver specimens. RNA was analyzed by bDNA assay for expression of MRP1, MRP2, MRP3, MRP4, MRP5, MRP6, MDR1, MDR3, and BCRP. The data are presented as mean relative light units (RLU) ± S.E. *, p < 0.05 versus normal liver.

Western blot analysis of sinusoidal efflux transporters. Western blots were performed using liver membrane fractions from normal, APAP, and PBC human liver specimens. The data are presented as representative blots (A), with each band representing an individual liver sample, and as mean relative MRP1, MRP3, MRP4, MRP5, or MRP6 protein expression ± S.E. (B). *, p < 0.05 versus normal liver.
Patterns of transporter expression in mouse liver after APAP administration have been well studied by our laboratory (Aleksunes et al., 2005, 2006b). Toxic APAP treatment enhanced liver levels of mouse Mrp2, 3, and 4 efflux transporters, which are responsible for export across the canalicular (Mrp2) and sinusoidal (Mrp3 and Mrp4) membranes (Aleksunes et al., 2005, 2006b). The most dramatically up-regulated efflux transporter in mouse liver, Mrp4 (16-fold induction), was also increased in human liver after APAP exposure (Fig. 3). Additional efflux transport proteins that were changed in human liver include MRP5, BCRP, and P-gp. Similar to human liver, APAP exposure induced P-gp protein levels in mice (unpublished observations from our laboratory) and rats (Ghanem et al., 2004).
In the mouse model there were minimal changes in transporter mRNA expression at low, subtoxic doses of APAP, suggesting that there is a threshold or dependence on the level of liver injury for changes in transporter expression to occur (Aleksunes et al., 2005). Differences in severity and progression of hepatic injury between APAP overdose patients and mice exposed to APAP may also account for some of the species differences in transporter isoform up-regulation. Liver sections from APAP overdose patients exhibited severe centrilobular necrosis and inflammation, compared with more modest centrilobular necrosis in the mouse. Although limited clinical information is available on the APAP dose ingested, time after exposure, and biochemical markers of hepatic damage for the APAP overdose patients, we know that the liver injury was severe enough to warrant liver transplantation. Although there are no standard selection criteria for liver transplantation after APAP poisoning, the King's College Hospital criteria are widely accepted (Dargan and Jones, 2002). In a previous prospective study patients who either died or received a liver transplant had a median alanine aminotransferase level of 4429 IU/liter with a range of 136 to 19,826 IU/liter (Larson et al., 2005). Considering that the severity of liver injury in the transplant recipient livers used in this study is probably similar to these documented cases and also taking into account the extent of histopathological damage, it must be considered that the changes in transporter expression may not be a result of the initial hepatic insult but may be due to complete liver failure. It is also possible that the increased efflux transporter levels may reflect a variation in the acinar distribution of transporters in response to the marked loss of pericentral hepatocytes in APAP overdose livers.

Western blot analysis of canalicular efflux transporters. Western blots were performed using liver membrane fractions from normal, APAP, and PBC human liver specimens. The data are presented as representative blots (A), with each band representing an individual liver sample, and as mean relative MRP2, BCRP, or P-gp protein expression ± S.E. (B). *, p < 0.05 versus normal liver.

Immunohistochemical localization of P-gp and PCNA. Double immunochemical staining of canalicular P-gp (brown) and nuclear PCNA (purple) was performed on paraffin sections from normal (A), APAP (B), and PBC (C) liver specimens. Tissues were counterstained with methyl green. Images of all groups were taken at 20× and 40× magnification.
Along with differences in the extent of hepatic injury, other factors contributing to the differences in transport protein expression between humans and mice may include activation or presence of divergent transcriptional pathways, gender, age, and concomitant disease states. Possible exposure to other medications, either before or during the overdose and also after admission to the hospital should also be considered. The medical profile of these patients was not available to us. Profiles of transporter expression during PBC have been well characterized, justifying use of these livers as positive controls. Similar alterations in transporters during cholestasis in rodents and humans indicate that transporter changes are common in multiple forms of liver diseases (Lee and Boyer, 2000). PBC is a chronic, cholestatic autoimmune disease characterized by inflammation and progressive destruction of bile ducts. Changes in transporter expression during PBC have been proposed to reduce the retention of toxic biliary constituents in hepatocytes as part of an adaptive response (Zollner et al., 2003). Zollner et al. (2003) reported an increase in MRP3 and P-gp expression, and Ros et al. (2003) also reported an increase in MRP3 and MDR1 protein, along with MRP1 protein (Ros et al., 2003). In the present study MRP1, MRP3, and P-gp protein expression was increased in PBC livers, as previously reported. In addition, novel changes in other efflux transport proteins were observed, with PBC livers exhibiting a substantial increase in protein expression levels of MRP4, MRP5, and BCRP.
The functional significance of altered transporter expression during these two different types of severe human liver disease may be better understood by considering the types of substrates effluxed by these transporters. Increases in transporters responsible for alternate routes of bile acid extrusion from hepatocytes may aid in the adaptive response to cholestatic liver injury through prevention of bile acid retention within hepatocytes (Zollner et al., 2003). MRP4 has recently been shown to transport bile acids and bile acid conjugates, which may be an additional means of retrograde efflux during cholestasis (Mennone et al., 2006; Rius et al., 2006). Other substrates of MRP4 include cyclic nucleotides (cAMP and cGMP), prostaglandins (E1 and E2), and glutathione (Reid et al., 2003; Bai et al., 2004; Rius et al., 2005). Similar to MRP4, MRP5 also transports cyclic nucleotides and is thought to be localized to the sinusoidal hepatocyte membrane (Jedlitschky et al., 2000; Wielinga et al., 2003). MRP6 has been shown to transport glutathione-conjugated leukotriene C4 and other glutathione conjugates (Belinsky et al., 2002; Iliás et al., 2002). In addition, MRP4 and MRP5 transport antiviral nucleotide analogs, whereas several anticancer agents such as etoposide and doxorubicin are potential substrates of MRP6 (Schuetz et al., 1999; Wijnholds et al., 2000; Belinsky et al., 2002). Sulfate conjugates of several steroids and xenobiotics are preferentially transported by BCRP (Suzuki et al., 2003). P-gp, or more specifically MDR1, transports a broad range of substrates including steroids and members of several classes of pharmaceuticals such as chemotherapeutic agents, cardiac glycosides, and narcotic analgesics (Matheny et al., 2001).
Enhanced export of multiple substrates across the sinusoidal and canalicular hepatocyte membrane may aid in minimizing their intracellular levels, thus limiting further injury and influencing the outcome of toxicant exposure. Alternatively, the exported compounds may function as paracrine signaling molecules to adjacent hepatocytes and nonparenchymal cells to facilitate the recovery process. As mentioned previously, MRP4 is capable of exporting prostaglandins, which have been documented to play an important role in hepatocyte proliferation after partial hepatectomy and to be hepatoprotective during liver ischemia-reperfusion injury (Rudnick et al., 2001; Reid et al., 2003; Hossain et al., 2006). In addition, cyclic nucleotides are substrates of both MRP4 and MRP5, which have also been shown to be important signaling molecules during hepatocyte proliferation and have been suggested to be a potential paracrine signaling pathway in the liver (Schlosser et al., 1996; Della Fazia et al., 1997). Importantly, elevated efflux transporter expression may also result in altered pharmacokinetics for a large number of therapeutic agents (Ayrton and Morgan, 2001). This finding may be relevant because a number of medications are prescribed for APAP overdose patients during their recovery in critical care units.
Prevention of further hepatocyte damage through increased efflux of potentially toxic compounds may serve as a compensatory response, but it is only one of several protective mechanisms that are set in motion after hepatic injury. Besides increased efflux transport proteins, decreased uptake transporter expression, induction of detoxification enzymes, and proliferation of hepatocytes are examples of protective compensatory responses to hepatic injury. Up-regulated protein and activity levels of NAD(P)H quinone oxidoreductase 1, an enzyme capable of reducing quinones and scavenging superoxide, were detected in these same liver specimens (Aleksunes et al., 2006a). Compensatory proliferation of hepatocytes is a critical component of recovery from hepatic injury and has been demonstrated to be an important part of the development of resistance to toxicants (Mehendale et al., 1994). Increased levels of hepatocyte proliferation are documented in both PBC and after APAP exposure (Rudi et al., 1995; Shankar et al., 2003). To determine the relationship between regions of hepatocyte proliferation and regions of enhanced transporter expression, immunochemical analysis of PCNA and P-gp was performed. Canalicular P-gp protein was localized to regions of PCNA-positive hepatocytes. This suggests that along with regeneration, increased efflux transporter expression is a critical response to hepatic damage to protect the liver from additional insult. Previous studies have shown that MDR1 gene expression is elevated in regenerating rat liver and in primary cultures of hepatocytes (Thorgeirsson et al., 1987; Lee et al., 1993), and it may be that in response to APAP-induced hepatotoxicity the dividing hepatocytes up-regulate efflux transporters. Alternatively, more rapid efflux of potentially harmful substances may enable the hepatocyte to more efficiently and successfully proliferate by protecting the genomic DNA during cell division. The close proximity and in some cases colocalization of P-gp-expressing hepatocytes to PCNA-positive hepatocytes also support the hypothesis that transporter-mediated efflux of substrates may be a way of signaling adjacent hepatocytes to divide.
The results of this study document the induction of efflux transport proteins in human livers after APAP overdose. Further studies in rodents are warranted to assess the functional contribution of altered transporter expression and to identify key substrates whose efflux may aid the recovery process. Future potential therapeutic approaches to the treatment of liver disease of varying etiologies may be designed to promote and exploit these adaptive defense mechanisms.
Footnotes
-
This work was supported in part by Pfizer Global Research and Development and by National Institutes of Health Grants DK069557 (J.E.M.) and DK068039 (N.J.C.). L.M.A. is a Howard Hughes Medical Institute Predoctoral Fellow and S.N.B. is a PhRMA Foundation Predoctoral Fellow. Human liver specimens were obtained from the Liver Tissue Procurement and Distribution System at the University of Minnesota, which is funded by National Institutes of Health Contract N01-DK-9-2310.
-
S.N.B. and L.M.A. contributed equally to this work.
-
Portions of this work were presented at the 2006 Annual Meeting of the Society of Toxicology (Barnes et al., 2006a), and at the 14th North American ISSX Meeting (Barnes et al., 2006b).
-
doi:10.1124/dmd.107.016170.
-
ABBREVIATIONS: APAP, acetaminophen; MRP (Mrp), multidrug resistance-associated protein; PBC, primary biliary cirrhosis; bDNA, branched DNA; MDR, multidrug resistance protein; BCRP, breast cancer resistance protein; BSEP, bile salt export pump; P-gp, P-glycoprotein; PCNA, proliferating cell nuclear antigen.
- Received April 5, 2007.
- Accepted July 9, 2007.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}