


Abstract
Rat lines congenic for the rat N-acetyltransferase 2 [(RAT)Nat2] gene were constructed and characterized. F344 (homozygous Nat2 rapid) males were mated to Wistar Kyoto (homozygous Nat2 slow) females to produce heterozygous F1. F1 females were then backcrossed to F344 males. Heterozygous acetylator female progeny from this and each successive backcross were identified by rat Nat2 genotyping and mated with F344 rapid acetylator males. After 10 generations of backcross mating, heterozygous acetylator brother/sister progeny were mated to produce the homozygous rapid and slow acetylator Nat2 congenic rat lines. p-Aminobenzoic acid (selective for rat NAT2) and 4-aminobiphenyl N-acetyltransferase activities were expressed in all tissues examined (liver, lung, esophagus, stomach, small intestine, colon, pancreas, kidney, skin, leukocytes, and urinary bladder in male and female rats and in breast of female and prostate of male rats). NAT2 expression in rat extrahepatic tissues was much higher than that in liver. In each tissue, activities were Nat2-genotype-dependent, with the highest levels in homozygous rapid acetylators, intermediate levels in heterozygous acetylators, and lowest in homozygous slow acetylators. Sulfamethazine (selective for rat NAT1) N-acetyltransferase activities were observed in all tissues examined in both male and female rats except for breast (females), bladder, and leukocytes. In each tissue, the activity was Nat2 genotype-independent, with similar levels in homozygous rapid, heterozygous, and homozygous slow acetylators. These congenic rat lines are useful for investigating the role of NAT2 genetic polymorphisms in susceptibility to cancers related to arylamine carcinogen exposures.
N-Acetyltransferase 1 (NAT1) and 2 (NAT2) catalyze the N-acetylation of aromatic amines (Hein et al., 1993). Genetic polymorphism in NAT2 segregates humans and other mammals such as rats into rapid and slow acetylators (Hein, 2002; Boukouvala and Fakis, 2005). Homozygous rapid (F344) and slow [Wistar Kyoto (WKY)] acetylator inbred rats have been characterized as an animal model for investigations of the N-acetylation polymorphism (Hein et al., 1991a,b; Juberg et al., 1991). (RAT)Nat1 and (RAT)Nat2 genes from rapid and slow acetylator rats each contain an intronless 870-base pair (bp) open reading frame (Doll and Hein, 1995). Rats also possess a third N-acetyltransferase locus (RAT)Nat3 (Walraven et al., 2006) that does not differ between F344 and WKY inbred strains (Walraven et al., 2007). Slow acetylator WKY inbred rats are homozygous for a rat Nat2 allele with four single nucleotide polymorphisms (SNPs): G361A (Val121→ Ile), G399A (synonymous), G522A (synonymous), and G796A (Val266→ Ile), compared with the Nat2 allele in the F344 rapid acetylator inbred rat (Doll and Hein 1995). NAT2 from F344 rat has been reported to exhibit significantly higher N-acetyltransferase activities than WKY in liver, kidney, colon, prostate, and urinary bladder (Hein et al., 1991a,b) and after recombinant expression in bacteria (Doll and Hein, 1995; Zhang et al., 2006). In contrast, Nat1 coding regions from rapid and slow acetylator rats are identical to each other, and their recombinant proteins exhibit equivalent N-acetyltransferase activities (Doll and Hein, 1995).
The Nat2 genetic polymorphism in the rat model has previously been shown to modify metabolism and toxicity of 3,2′-dimethyl-4-aminobiphenyl (Feng et al., 1997; Jiang et al., 1999), 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (Purewal et al., 2000a,b), 6-hydroxydopamine (Grundmann et al., 2004), and 4,4′-methylene dianiline (Zhang et al., 2006). However, F344 and WKY inbred rat strains differ in many other characteristics, including other metabolic enzymes and DNA repair. Thus, we constructed and characterized rat lines congenic for the rat (RAT)Nat2 gene and compared rat NAT1 and NAT2 expression with previously reported mouse and Syrian hamster congenic lines.
Materials and Methods
Animals. F344 (rapid Nat2 acetylator) and WKY (slow Nat2 acetylator) rats were purchased from Charles River Laboratories, Inc. (Wilmington, MA). All procedures were approved by the University of Louisville Institutional Animal Care and Use Committee.
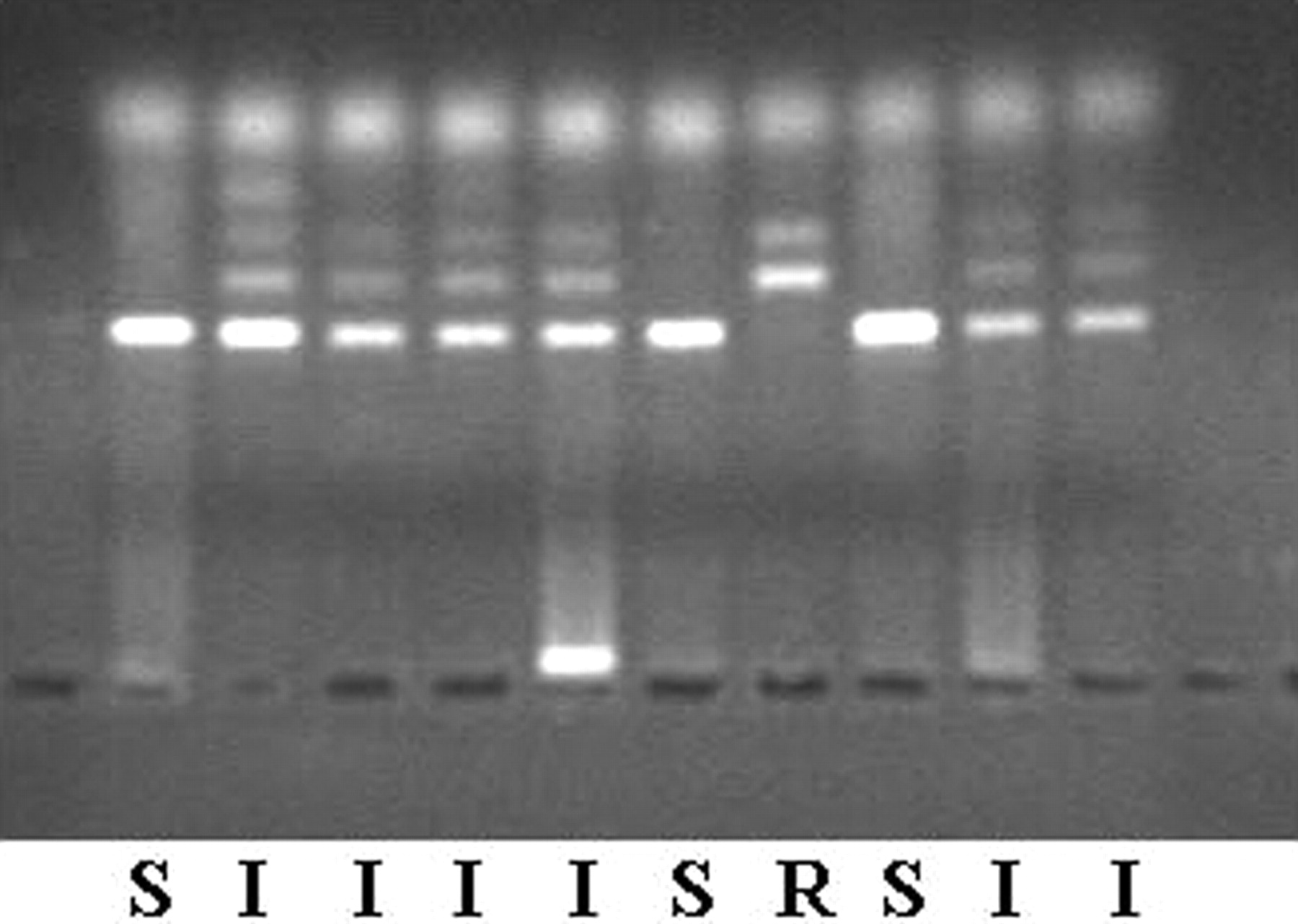
Rat Nat2 Genotyping. Rat genomic DNA was isolated from blood or tail clips using proteinase K followed by phenol-chloroform extraction. Approximately 0.25 to 0.5 inch of tail was removed, minced with a scalpel, and homogenized in a buffer containing 1% sodium dodecyl sulfate, 1 mM EDTA, 25 mM Tris-HCl, pH 7.4, and 0.5 mg of proteinase K. Samples were incubated at 37°C for 1 h and then extracted with equal volumes of phenol, phenolchloroform, and chloroform. The DNA in the aqueous layer was precipitated by the addition of one-tenth volume of 5 M NaCl and 3 volumes of 95% ethanol. The DNA was washed twice with 70% ethanol, dried in air, and resuspended in 10 mM Tris-HCl-1 mM EDTA, pH 8.0. Rat Nat2 was amplified by polymerase chain reaction (PCR) and digested with the restriction enzyme RsaI. The PCR reaction contained 10 mM Tris-HCl, 50 mM KCl, 3.0 mM MgCl2, 0.2 mM concentrations of each deoxynucleotide triphosphate, 0.2 μg of each primer (5′-AGGACACCAAAACTGCATAT-3′ and 5′-CAGCATCCTTGTTTACAAGT-3′), 0.6 U of Taq DNA polymerase (Applied Biosystems, Foster City, CA), and 10 to 100 ng of rat genomic DNA. Samples were incubated at 94°C for 5 min, followed by 30 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 1 min. The reaction was concluded with incubation at 72°C for 5 min. The 257-bp PCR product was digested with the restriction enzyme RsaI following the manufacturer's instruction. Digested PCR products were separated on a 2% agarose gel and visualized under UV light. Samples homozygous for the rapid Nat2 allele possessed bands of 160 and 97 bp. Samples homozygous for the slow Nat2 allele had a single band at 257 bp. Samples heterozygous for Nat2 alleles had bands of 257, 160, and 97 bp (Fig. 1).

Restriction fragment length polymorphism Nat2 genotyping. The 257-bp PCR product was digested with the restriction enzyme RsaI following the manufacturer's instruction. Digested PCR products were separated on a 2% agarose gel and visualized under UV light. Samples homozygous for the rapid Nat2 allele (R) possessed bands of 160 and 97 bp. Samples homozygous for the slow Nat2 allele (S) had a single band at 257 bp. Samples heterozygous for Nat2 alleles (I) had bands of 257, 160, and 97 bp.
Construction of Rapid and Slow Acetylator Rat Lines Congenic at the Nat2 Locus. Rapid and slow acetylator congenic rat lines were constructed with methods previously used by our laboratory to construct rapid and slow acetylator congenic Syrian hamster lines (Hein et al., 1994). F344 (homozygous Nat2 rapid) males were mated to WKY (homozygous Nat2 slow) females to produce the obligate heterozygous F1 generation. F1 females were then backcrossed to F344 males. Heterozygous F1 acetylator female progeny from this and each successive backcross were identified by rat Nat2 genotyping (as described above) and mated with F344 rapid acetylator males. After 10 generations of backcross mating, heterozygous acetylator brother/sister progeny were mated to produce the homozygous rapid and slow acetylator congenic rat lines. The genome from the WKY inbred rat was the source for the slow acetylator Nat2 allele, which was reduced by double dilution of each backcross generation (Fig. 2).
Preparation of Tissue Cytosols. Adult rats were sacrificed by carbon dioxide asphyxiation and the liver, lung, colon stomach, breast (females), prostate (males), small intestine, pancreas, urinary bladder, kidney, esophagus, leukocytes, and skin were collected. Tissues were homogenized in 20 mM sodium phosphate (pH 7.4), 1 mM EDTA, 1 mM dithiothreitol, 100 μM phenylmethanesulfonyl fluoride, 10 μg/ml aprotinin, and 1 μM pepstatin. Homogenates were centrifuged at 100,000g for 1 h, and supernatants were aliquoted and stored at -80°C until used. Protein concentrations were determined using the Bio-Rad protein assay kit (Bio-Rad, Hercules, CA).
N-Acetyltransferase Assays. Tissue cytosols were assayed for the level of NAT2 activity using the NAT2-selective substrate PABA and for NAT1 activity using the NAT1-selective substrate SMZ as described previously (Leff et al., 1999). No selective substrates for rat NAT3 have been identified, and it does not catalyze the N-acetylation of PABA or SMZ (Walraven et al., 2006). In brief, reactions containing tissue cytosol (<2 mg of protein/ml), 300 μM PABA or SMZ, and 1 mM acetyl-coenzyme A were incubated at 37°C for 10 min. Reactions were terminated by the addition of one-tenth volume of 1 M acetic acid. The reaction tubes were centrifuged to precipitate protein, and supernatant was injected onto a LiChrospher 100 RP-18 (125 mm × 4 mm; 5 μm) reverse-phase column. Reactants and products were separated by high-performance liquid chromatography (Beckman Coulter, Fullerton, CA). N-Acetyl-PABA and N-acetyl-SMZ were quantitated by their absorbance at 280 and 260 nm, respectively. ABP N-acetyltransferase assays were carried out as described previously (Hein et al., 2006). In brief, reactions containing suitably diluted cytosol, ABP (1 mM), and acetyl coenzyme A (1 mM) were incubated for 10 min at 37°C and terminated by the addition of one-tenth volume of 1 M acetic acid. The reaction tubes were centrifuged to precipitate protein, and supernatant was injected onto a LiChrospher 100 RP-18 (125 mm × 4 mm; 5 μm) reverse-phase column. Reactants and products were separated by high-performance liquid chromatography. N-Acetyl-ABP was measured by its absorbance at 260 nm.

Construction of F344 Nat2 congenic lines. The percentage of F344 genome (○) and WKY genome (▪) is illustrated for the F1 and each congenic (N) generation. Heterozygous Nat2 acetylator males and females from the N10 generation were bred to produce homozygous rapid and homozygous slow Nat2 acetylator F344 rats.
Results
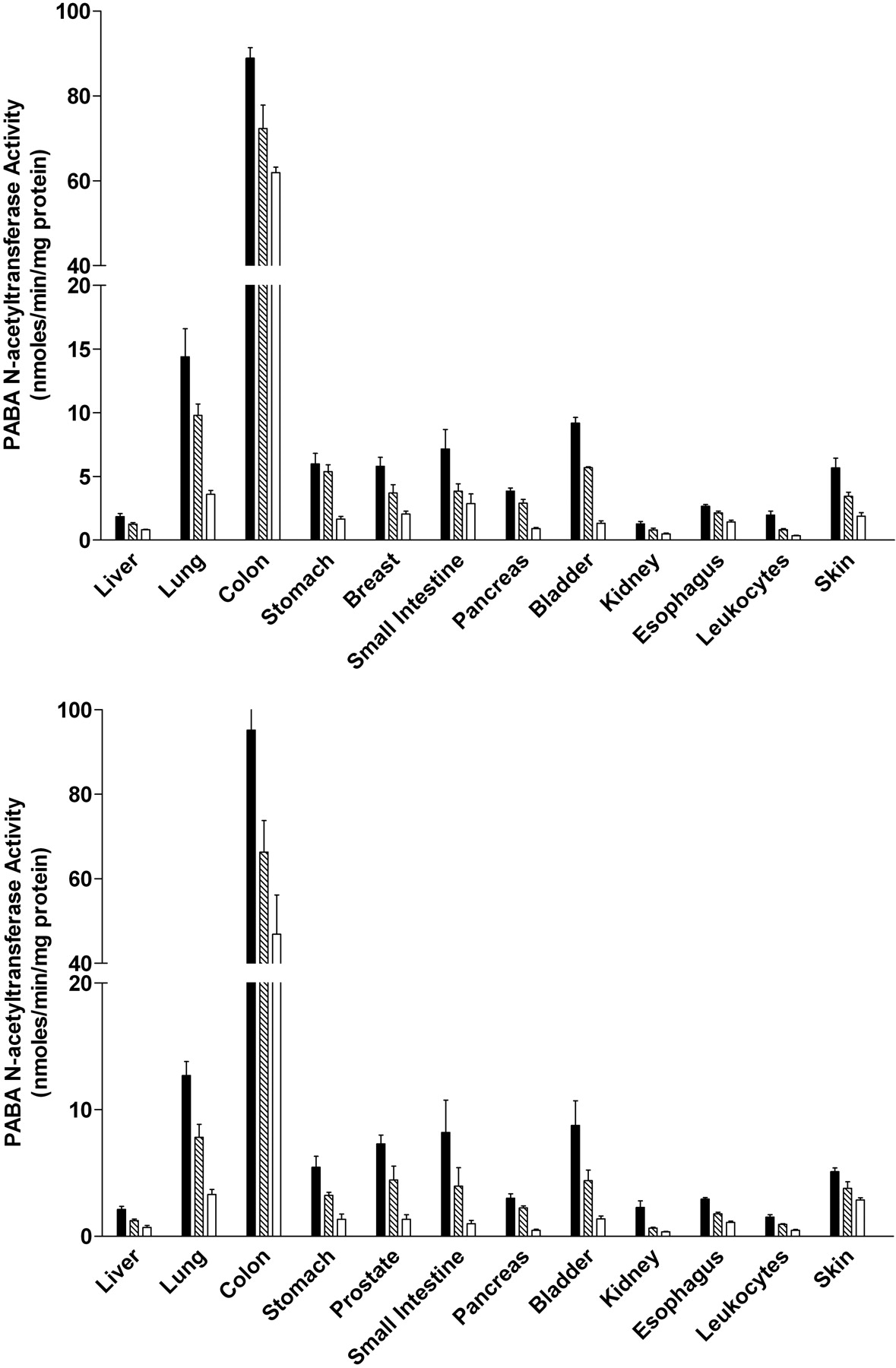
NAT2 Activities. PABA (selective for rat NAT2) N-acetyltransferase activities were readily detected in all tissues examined in both male and female rats (Fig. 3). In each tissue, the activity was Nat2 genotype-dependent, with the highest levels in homozygous rapid acetylators, intermediate levels in heterozygous acetylators, and lowest levels in homozygous slow acetylators. The magnitude of activity differences between rapid and slow acetylators ranged from 1.4- to 8.3-fold. NAT2 activities differed with tissue in the following relative order: colon  lung > small intestine, urinary bladder > stomach, breast or prostate, pancreas, skin > liver, kidney, leukocytes. NAT2 activity levels did not differ significantly between males and females in any tissue examined.
lung > small intestine, urinary bladder > stomach, breast or prostate, pancreas, skin > liver, kidney, leukocytes. NAT2 activity levels did not differ significantly between males and females in any tissue examined.
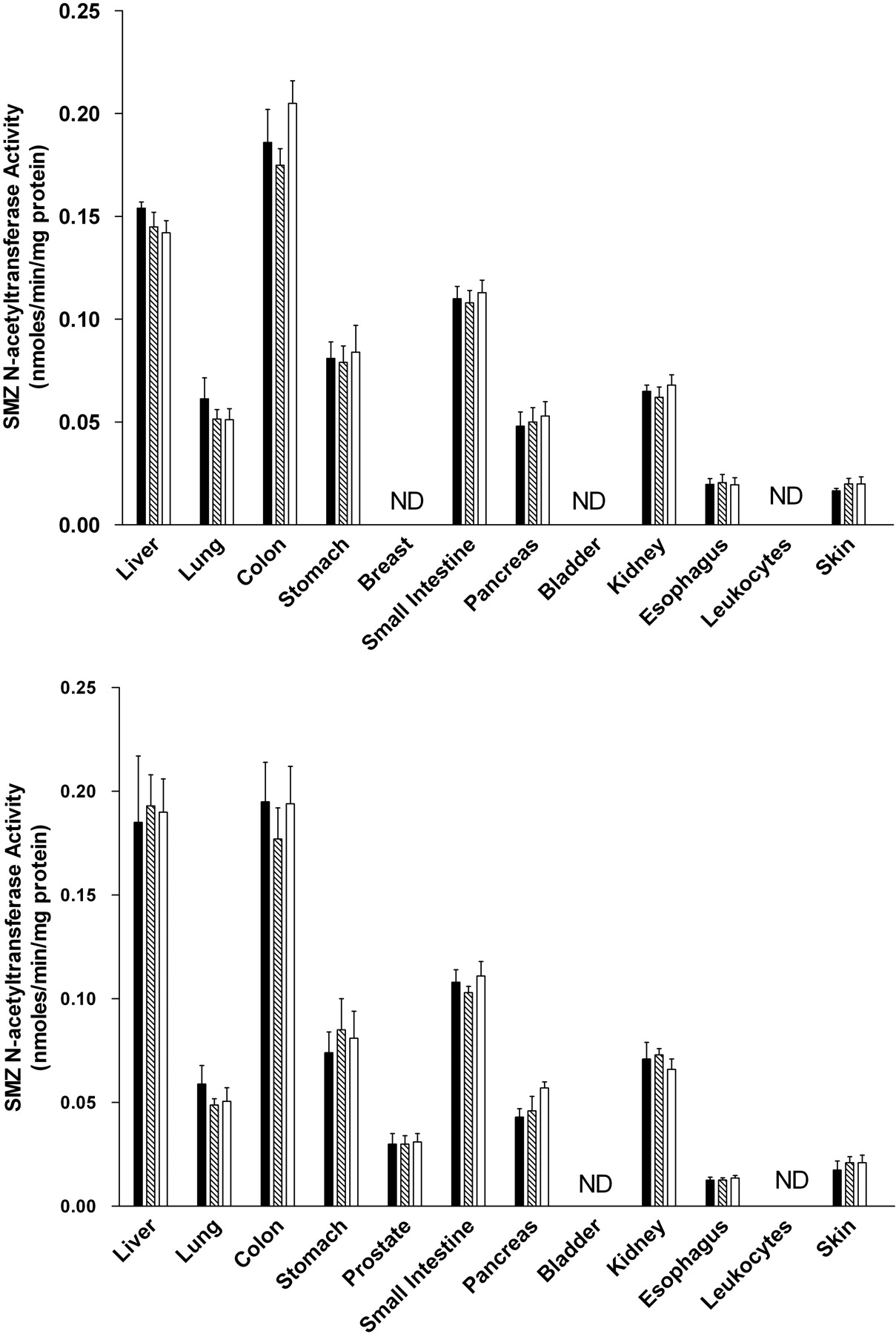
NAT1 Activities. SMZ (selective for rat NAT1) N-acetyltransferase activities were observed in all tissues examined in both male and female rats except for breast (females), bladder, and leukocytes (Fig. 4). In each tissue, the activity was Nat2-genotype independent, with similar levels in homozygous rapid, heterozygous, and homozygous slow acetylators. NAT1 activities differed with tissue in the following relative order: colon, liver >small intestine > stomach, kidney, lung, pancreas >esophagus, skin, prostate (males). NAT1 activity levels did not differ significantly between males and females in any tissue examined.

PABA N-acetyltransferase activities in female (top) and male (bottom) tissues as listed on the abscissa. PABA N-acetyltransferase activities differed significantly (p < 0.05) with respect to Nat2 genotype in every tissue. ▪, homozygous Nat2 rapid acetylator genotype;  , heterozygous intermediate acetylator Nat2 genotype; □, homozygous Nat2 slow acetylator genotype. Each bar is the mean ± S.E.M. for five individual rats.
, heterozygous intermediate acetylator Nat2 genotype; □, homozygous Nat2 slow acetylator genotype. Each bar is the mean ± S.E.M. for five individual rats.
ABP N-Acetyltransferase Activities. ABP N-acetyltransferase activities were readily detected in all tissues examined in both male and female rats (Fig. 5). In each tissue, the activity was Nat2 genotype-dependent, with the highest levels in homozygous rapid acetylators, intermediate levels in heterozygous acetylators, and lowest levels in homozygous slow acetylators. The magnitudes of activity differences between rapid and slow acetylators were more modest than those observed with PABA, reflecting the fact that ABP is not a NAT2-selective substrate in the rat. ABP N-acetyltransferase activities differed with tissue in the following relative order: colon  lung, small intestine, urinary bladder, stomach, breast (female), prostate (male), skin, leukocytes > liver, kidney, pancreas. ABP N-acetyltransferase activity levels did not differ significantly between males and females in any tissue examined.
lung, small intestine, urinary bladder, stomach, breast (female), prostate (male), skin, leukocytes > liver, kidney, pancreas. ABP N-acetyltransferase activity levels did not differ significantly between males and females in any tissue examined.
Discussion
Nat1 and Nat2 in animal models such as rabbit, mouse, Syrian hamster, and rat are highly homologous to both human NAT1 and NAT2 (Hein, 2002; Boukouvala and Fakis, 2005). Several different mechanisms are responsible for Nat2 polymorphisms in nonhuman species. The molecular basis for the slow acetylator phenotype is Nat2 gene deletion in the rabbit (Blum et al., 1989), a nonsense SNP yielding a truncated NAT2 enzyme in the Syrian hamster (Ferguson et al., 1994; Nagata et al., 1994), and nonsynonomous SNP(s) in the mouse (Martell et al., 1991) and rat (Doll and Hein, 1995). The molecular basis for the slow acetylator phenotype in humans likewise is nonsynonomous SNPs in the NAT2 coding region (Hein, 2002).

SMZ N-acetyltransferase activities in female (top) and male (bottom) tissues as listed on the abscissa. SMZ N-acetyltransferase activities did not differ significantly (p > 0.05) with respect to Nat2 genotype in any tissue. ▪, homozygous Nat2 rapid acetylator genotype;  , heterozygous intermediate acetylator Nat2 genotype; □, homozygous Nat2 slow acetylator genotype. Each bar is the mean ± S.E.M. for five individual rats. ND, nondetectable activity.
, heterozygous intermediate acetylator Nat2 genotype; □, homozygous Nat2 slow acetylator genotype. Each bar is the mean ± S.E.M. for five individual rats. ND, nondetectable activity.
ABP is a widespread environmental carcinogen present in cigarette smoke (Stabbert et al., 2003) and cooking oil fumes (Chiang et al., 1999) that induces breast tumors in the rat (Tanaka et al., 1985). ABP DNA adducts have been detected in human breast (Gorlewska-Roberts et al., 2002; Faraglia et al., 2003; Ambrosone et al., 2007), and the NAT2 genotype has been shown to modify breast cancer risk in smokers (van der Hel et al., 2003; Ambrosone et al., 2008). Thus, ABP bioassays in rapid and slow acetylator rats have been proposed. PABA (selective for rat NAT2) and ABP N-acetyltransferase activities were expressed in all tissues examined (liver, lung, esophagus, stomach, small intestine, colon, pancreas, kidney, skin, leukocytes, and urinary bladder in male and female rats and in breast of female and prostate of male rats). In each tissue, activities were clearly Nat2 genotype-dependent, with the highest levels in homozygous rapid acetylators, intermediate levels in heterozygous acetylators, and lowest levels in homozygous slow acetylators. Higher levels of N-acetyltransferase activity in F344 versus WKY inbred rat strains have been reported previously in several rat tissues (Hein et al., 1991a,b; Juberg et al., 1991; Martell and Weber, 1993; Ware et al., 1995; Ware and Svensson, 1996) In contrast, SMZ N-acetyltransferase activities were Nat2-genotype independent, with similar levels in homozygous rapid, heterozygous, and homozygous slow acetylators. These results reflect the substrate specificity of PABA for rat NAT2 and SMZ for rat NAT1 (Walraven et al., 2006). Although a rat NAT3-selective substrate has yet to be identified, rat Nat3 transcripts are at least 100- to 1000-fold lower than rat Nat1 and Nat2 transcripts in rat hepatic and extrahepatic tissues (Walraven et al., 2007; Barker et al., 2008).
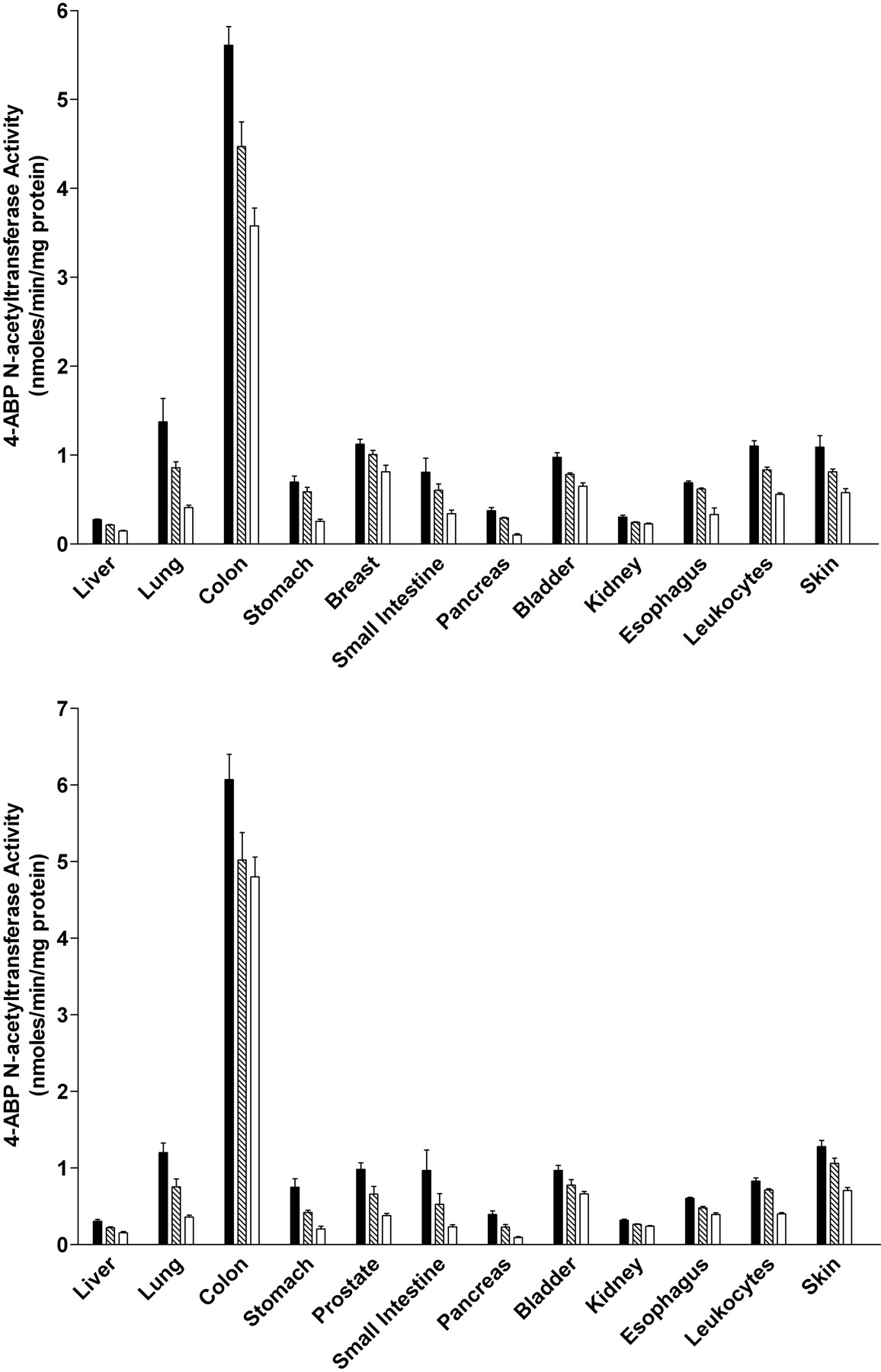
ABP N-acetyltransferase activities in female (top) and male (bottom) tissues as listed on the abscissa. ABP N-acetyltransferase activities differed significantly (p < 0.05) with respect to Nat2 genotype in every tissue. ▪, homozygous Nat2 rapid acetylator genotype;  , heterozygous intermediate acetylator Nat2 genotype; □, homozygous Nat2 slow acetylator genotype. Each bar is the mean ± S.E.M. for five individual rats.
, heterozygous intermediate acetylator Nat2 genotype; □, homozygous Nat2 slow acetylator genotype. Each bar is the mean ± S.E.M. for five individual rats.
In a congenic rat model in which all slow acetylators are homozygous for a single rat slow Nat2 allele and obligate heterozygotes all possess the same combination of rapid and slow rat Nat2 alleles, the Nat2 acetylation polymorphism clearly segregates into three phenotypes in hepatic and extrahepatic tissues. This trimodal distribution of rapid, intermediate, and slow acetylator phenotypes also is clearly evident in the congenic Nat2 Syrian hamster model (Hein et al., 1991c, 1992, 1994). Although many human studies often exhibit bimodal distributions of rapid and slow acetylator NAT2 phenotypes, they also often yield trimodal distributions of rapid, intermediate, and slow acetylator phenotypes (Hein, 2006).

A widely held hypothesis is that human NAT2 is expressed primarily in liver and gastrointestinal tract, whereas human NAT1 has widespread tissue distribution. This hypothesis derives from studies in the rabbit model in which N-acetyltransferase activities reflected the Nat2 genetic polymorphism in liver and gut but not in other tissue cytosols, suggesting either absence or a much smaller contribution of rabbit NAT2 in these other tissues (Hearse and Weber, 1973). Studies have shown widespread tissue distribution of human NAT1 and NAT2 mRNA (Boukouvala and Fakis, 2005; Barker et al., 2006; Husain et al., 2007a,b). Although extrahepatic expression of N-acetyltransferase activities have been reported in rat (Hein et al., 1991a), mouse (Chung et al., 1993; Stanley et al., 1997; Sugamori et al., 2003; Wakefield et al., 2008), and Syrian hamster (Hein et al., 1991c, 1992, 1994), substrates were not selective for NAT1 or NAT2.
Recent studies with substrates selective for NAT1 versus NAT2 reported widespread distribution of both NAT1 and NAT2 catalytic activities in the rapid and slow acetylator congenic hamster (Hein et al., 2006). NAT2 catalytic activity in the Syrian hamster followed the relative order: liver > small intestine, cecum > colon, rectum, lung, pancreas, stomach, bladder > prostate, esophagus, heart. Thus, the rat differs markedly from the Syrian hamster with respect to hepatic versus extrahepatic expression of NAT2 activity. NAT1 catalytic activity in the Syrian hamster followed the relative order: liver  small intestine, cecum, colon, rectum, pancreas, stomach, esophagus, lung > prostate and heart (Hein et al., 2006). Thus, the relative hepatic versus extrahepatic NAT1 expression seems to be fairly similar in rat and Syrian hamster. No differences in NAT1 expression were observed between rapid and slow acetylators, consistent with our findings in the rat. ABP N-acetyltransferase activity in the Syrian hamster followed the relative order: liver, small intestine > cecum, colon, rectum, pancreas, stomach, lung, prostate, esophagus, bladder, heart (Hein et al., 2006). Thus, the contribution of hepatic versus extrahepatic NAT2 to 4-aminobiphenyl metabolism is much less in the rat than in the Syrian hamster. Lower and Bryan (1973) reported previously that ABP N-acetyltransferase activity was much higher in hamster than in rat liver. Previous reports clearly showed that PABA N-acetyltransferase activity was much higher in rat colon (Hein et al., 1991a) than in liver (Hein et al., 1991b). Subsequent reports confirmed that rat PABA N-acetyltransferase activity was much lower in rat liver than in the intestine (Ware et al., 1995) or colon (Purewal et al., 2000a).
small intestine, cecum, colon, rectum, pancreas, stomach, esophagus, lung > prostate and heart (Hein et al., 2006). Thus, the relative hepatic versus extrahepatic NAT1 expression seems to be fairly similar in rat and Syrian hamster. No differences in NAT1 expression were observed between rapid and slow acetylators, consistent with our findings in the rat. ABP N-acetyltransferase activity in the Syrian hamster followed the relative order: liver, small intestine > cecum, colon, rectum, pancreas, stomach, lung, prostate, esophagus, bladder, heart (Hein et al., 2006). Thus, the contribution of hepatic versus extrahepatic NAT2 to 4-aminobiphenyl metabolism is much less in the rat than in the Syrian hamster. Lower and Bryan (1973) reported previously that ABP N-acetyltransferase activity was much higher in hamster than in rat liver. Previous reports clearly showed that PABA N-acetyltransferase activity was much higher in rat colon (Hein et al., 1991a) than in liver (Hein et al., 1991b). Subsequent reports confirmed that rat PABA N-acetyltransferase activity was much lower in rat liver than in the intestine (Ware et al., 1995) or colon (Purewal et al., 2000a).
Mouse N-acetyltransferase catalytic activity was found in all tissues examined except blood plasma and seminal vesicles (Chung et al., 1993). Activity in lymphoid tissue, in general, was high as was that in skin and much of the digestive system. The NAT2 polymorphism with PABA and 2-aminofluorene was apparent in most tissues (except brain, muscle, parotid gland, submaxillary gland, and testis). Subsequently, Loehle et al. (2006) reported that mouse Nat1 and Nat2 mRNA and catalytic activities were present in all mouse tissues examined (i.e., liver, gut, pancreas, bladder, and prostate). Similar results were reported by Sugamori et al. (2006, 2007) with activities in the following relative order: liver > kidney, colon, spleen > lung, bladder > cortex. Thus, NAT2 expression in the rat differs from that in both Syrian hamster and mouse. Mouse N-acetyltransferase activities were relatively constant across tissues and gender, with the exception of kidney in which activities were approximately 2-fold higher in males than in females (Sugamori et al., 2007). Previous rat studies on gender have been inconsistent with one study reporting that female N-acetyltransferase activities were lower than those in males in several rat tissues (Juberg et al., 1991), whereas another study reported that female N-acetyltransferase activity was higher in female than in male kidney (Ware et al., 1995). In contrast, we did not observe gender-related differences in any tissue for PABA, SMZ, or ABP N-acetyltransferase activity in the rat.
NAT2 activities were higher than NAT1 activities in all tissues. The NAT2/NAT1 activity ratio varied considerably from tissue to tissue but did not vary much between males and females (Fig. 6). Differences between NAT2 and NAT1 activity were highest in colon (500-fold) and lowest in liver (2-fold). Thus, the tissue with the highest (colon) and lowest (liver) N-acetyltransferase activity also showed the highest and lowest difference between NAT2 and NAT1 activity.
In conclusion, rat lines congenic for the (RAT)Nat2 gene were constructed and characterized. NAT2 expression across various tissues differed markedly from that previously reported in mouse and Syrian hamster congenic lines. Because many carcinogenic bioassay studies are conducted in the rat, particularly for arylamine carcinogens that share target organ specificity with humans, these congenic rat lines are useful for investigating the role of Nat2 genetic polymorphism in susceptibility to cancers related to arylamine carcinogen exposures. Bioassays with ABP are in progress.
Footnotes
-
This study was supported by U.S. Public Health Service Grants R01-CA034627 and P30-ES014443.
-
Parts of this work were previously presented as follows: Hein D, Neale JR, Bendaly J, and Doll MA (2007) Construction and initial characterization of Nat2 congenic rat lines. Fourth International Workshop on the Arylamine N-Acetyltransferases; 2007 Sept 14–16; Alexandroupolis, Greece.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.108.023960.
-
ABBREVIATIONS: Nat1/NAT1, N-acetyltransferase 1; Nat2/NAT2, N-acetyltransferase 2; bp, base pair; SNP, single nucleotide polymorphism; PCR, polymerase chain reaction; PABA, p-aminobenzoic acid; SMZ, sulfamethazine; ABP, 4-aminobiphenyl.
- Received August 15, 2008.
- Accepted September 16, 2008.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}