


Abstract
Buprenorphine (BN) is a thebaine derivative with analgesic properties. To identify and characterize the cytochrome P450 (CYP) enzyme(s) involved in BN N-dealkylation, in vitro studies using human liver microsomes and recombinant human CYP enzymes were performed. Norbuprenorphine formation from BN was measured by a simple HPLC-UV assay method, without extraction. The BNN-dealkylation activities in 10 human liver microsomal preparations were strongly correlated with microsomal CYP3A-specific metabolic reactions, i.e. triazolam 1′-hydroxylation (r = 0.954), midazolam 1′-hydroxylation (r = 0.928), and testosterone 6β-hydroxylation (r = 0.897). Among the eight recombinant CYP enzymes studied (CYP1A2, CYP2A6, CYP2B6, CYP2C9, CYP2C19, CYP2D6, CYP2E1, and CYP3A4), only CYP3A4 could catalyze BN N-dealkylation. The apparent KM value for recombinant CYP3A4 was similar to that for human liver microsomes (23.7 vs.39.3 ± 9.2 μM). The demonstration of BNN-dealkylation by recombinant CYP3A4 and the agreement in the affinities (apparent KM values) of human liver microsomes and recombinant CYP3A4 provide the most supportive evidence for BN N-dealkylation being catalyzed by CYP3A4.
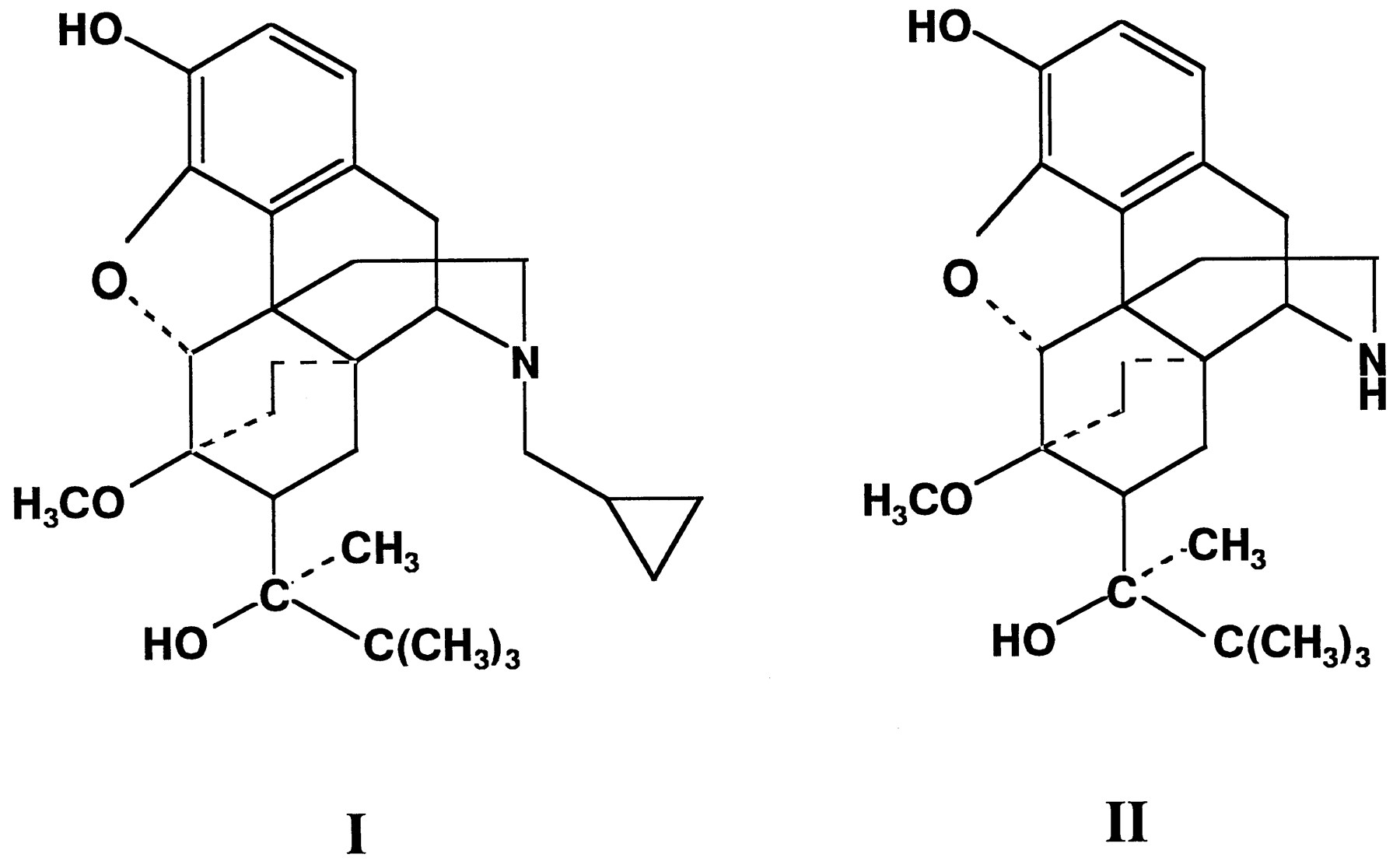
BN1(fig. 1) is a synthetic derivative of the morphine alkaloid thebaine with mixed agonist/antagonist analgesic properties (Cowan et al., 1977b; Lewis, 1974). The analgesic effects of BN are 30 times more potent than those of morphine, and it produces little physical dependence (Cowan et al., 1977a,b; Jasinski et al., 1978). In human subjects, BN is metabolized by phase I and phase II reactions to NBN and conjugates of BN and NBN, respectively (Brewster et al., 1981). After oral BN administration, no free BN was detected in urine and the amounts of NBN metabolites excreted in urine generally exceeded those of conjugated BN (Coneet al., 1984). NBN is produced by N-dealkylation of the N-cyclopropylmethyl group, which is a principal metabolic pathway for BN (Cone et al., 1984).
Most oxidative metabolism of xenobiotics, including drugs, environmental pollutants, and procarcinogens, is catalyzed by CYP enzymes, which comprise a large family of hemoproteins (Guengerich, 1992; Gonzalez, 1989; Wrighton and Stevens, 1992). More than 15 isozymes have been identified in human liver, and several forms play important roles in xenobiotic metabolism in humans (Kerremans, 1996). The structures of codeine and dextromethorphan, which are extensively metabolized by CYP, are similar to that of BN. Both codeine and dextromethorphan are metabolized via O3-demethylation by CYP2D6 andN17-demethylation by CYP3A4 (Ladonaet al., 1991; Yue et al., 1991; Jacqz-Aigrainet al., 1993). Although BN also has anN-cyclopropylmethyl group at the 17-position, the enzyme responsible for the N-dealkylation of BN in human subjects had not been identified. Recently, Iribarne et al. (1997)reported that BN N-dealkylation in human liver microsomes is mainly catalyzed by CYP3A4. To further characterize the enzyme(s) responsible for BN N-dealkylation in human liver microsomes, we performed a kinetic study using human liver microsomes and cDNA-expressed human CYP enzymes.
Materials and Methods
Chemicals.
BN and NBN were supplied by Otsuka Pharmaceuticals (Tokushima, Japan). Triazolam and 1′-hydroxytriazolam were generous gifts from Pharmacia Upjohn Co. (Tokyo, Japan), midazolam and 1′-hydroxymidazolam from Japan Roche Co. (Tokyo, Japan), and ketoconazole from Janssen Research Foundation (Beerse, Belgium). Testosterone and 6β-hydroxytestosterone were purchased from Daiichi Pure Chemical Co. (Tokyo, Japan). Acetonitrile and methanol of HPLC grade ando-ethoxybenzamide, NADP+, glucose-6-phosphate, and glucose-6-phosphate dehydrogenase were purchased from Wako Pure Chemical Industries (Osaka, Japan).
Assays with Human Liver Microsomes.
Ten samples of human liver microsomes were obtained from Japanese patients undergoing partial hepatectomy for treatment of metastatic tumors at the Division of General Surgery, Department of Surgery, International Medical Center of Japan (Tokyo, Japan), and were prepared as reported previously (Chiba et al., 1993). The primary incubation medium contained 0.1 mg/ml human liver microsomes, 0.5 mM NADP+, 2.0 mM glucose-6-phosphate, 1 IU/ml glucose-6-phosphate dehydrogenase, 4 mM MgCl2, 0.1 mM EDTA, 100 mM potassium phosphate buffer (pH 7.4), and 2.5–150 μM BN, in a final volume of 250 μl. The mixture was incubated at 37°C for 15 min, and the reaction was stopped by the addition of 100 μl of cold acetonitrile. After termination of the incubation, 50 μl of o-ethoxybenzamide (1 μg/ml in methanol) was added to the samples as an internal standard. The mixture was centrifuged at 10,000g for 5 min, and 100 μl of supernatant was analyzed by HPLC as described below.
HPLC Conditions.
NBN determinations were carried out using an HPLC-UV assay method. The HPLC system consisted of a model L-6000 pump (Hitachi, Tokyo, Japan), a model L-4000 UV detector (Hitachi), a model AS-2000 autosampler (Hitachi), a model D-2500 integrator (Hitachi), and a 4.6- × 250-mm CAPCELL PAK C18 UG120 column (Shiseido, Tokyo, Japan). The mobile phase consisted of 0.05 M potassium phosphate buffer (pH 4.0)/acetonitrile (72:28, v/v) and was delivered at a flow rate of 0.8 ml/min. The eluate was monitored at a wavelength of 232 nm. The column temperature was maintained at 30°C. Calibration curves were generated from 0.3 to 3 nmol/ml by processing the authentic standard substance through the entire procedure. Under these chromatographic conditions, NBN, o-ethoxybenzamide, and BN were eluted at 7.7, 9.5, and 21.3 min, respectively. The detection limit for NBN was 1.5 ng. NBN was quantified by comparison with the standard curves, using the peak height ratio method. Intraassay (N = 6) coefficients of variation were 8.5%.
Kinetics of BN N-Dealkylation.
The rate of formation of NBN was linear for up to 15 min when 100 μM BN and 0.1 mg/ml human liver microsomal protein were present. A linear relationship was also observed between the rate of NBN formation in 15 min and protein concentrations of up to 0.15 mg/ml. The rate of formation of NBN from 2.5 μM BN was also linear with up to 0.15 mg/ml human liver microsomal protein concentrations in 15-min incubations. Accordingly, the kinetic studies were performed at 37°C with a 15-min incubation time and a protein concentration of 0.1 mg/ml. The kinetic studies were performed using five different human liver microsomes. The kinetic parameters (KM ,Vmax, andVmax/KM ) for BN N-dealkylation were determined from the rate of formation of NBN at BN concentrations ranging from 2.5 to 150 μM and were estimated by linear regression analysis of Eadie-Hofstee plots.
Correlation Study.
The BN N-dealkylation activities were compared with CYP3A-specific metabolic reactions, i.e. triazolam 1′-hydroxylation, midazolam 1′-hydroxylation, and testosterone 6β-hydroxylation, using microsomes obtained from 10 human liver samples. The substrate concentrations used were 25 μM for BN, triazolam, and midazolam and 30 μM for testosterone. Assays were performed in duplicate on the same day with the same set of microsomal preparations. 6β-Hydroxytestosterone, 1′-hydroxymidazolam, and 1′-hydroxytriazolam were determined using the respective HPLC-UV assay methods reported previously (Yoshimoto et al., 1995;Kobayashi et al., 1997). Correlations between BNN-dealkylation and each of the three CYP3A-specific metabolic reactions were examined with the least-squares linear regression method.
Assays with Recombinant CYP Enzymes.
Microsomes from human B lymphoblastoid cells expressing human CYP1A2 (lot 29), CYP2A6 (lot 17), CYP2B6 (lot 20), CYP2C9-Cys (lot 10), CYP2C19 (lot 1), CYP2D6-Met (lot 38), CYP2E1 (lot 18), and CYP3A4 (lot 4) (Gentest, Woburn, MA) were used. Recombinant CYP2C9, CYP2E1, and CYP3A4 were coexpressed with NADPH-CYP reductase in human B lymphoblastoid cells. The reactions were carried out as described for the human liver microsomal study. To examine the roles of individual CYP enzymes involved in BN N-dealkylation, each of the eight recombinant CYP enzymes (0.5 mg/ml protein concentration) described above was first incubated with 100 μM BN for 120 min at 37°C, according to the procedure recommended by the supplier.
Results and Discussion
In this study, BN N-dealkylation in human liver microsomes was assessed with a newly developed HPLC-UV assay method. The intraassay coefficient of variation was <10%, and the detection limit was as low as 1.5 ng for NBN. Therefore, this method was applicable to determinations of N-dealkylation activity at low concentrations of BN (i.e. 2.5 μM). Eadie-Hofstee plots for BN N-dealkylation yielded a straight line for all samples studied, suggesting that BN N-dealkylation by human liver microsomes shows simple Michaelis-Menten kinetics (fig.2). The mean ± SD values for kinetic parameters were as follows: apparentKM = 39.3 ± 9.2 μM,Vmax = 0.712 ± 0.279 nmol/min/mg of protein, andVmax/KM = 17.8 ± 3.1 μl/min/mg of protein (table1).

Chemical structures of BN ( I ) and NBN ( II ).

Representative Eadie-Hofstee plots for BNN-dealkylation by human liver microsomes (•) and recombinant CYP3A4 (○).
The BN N-dealkylation activities were determined from the rates of formation of NBN at substrate concentrations ranging from 2.5 to 150 μM. V, BN N-dealkylation activity; S, concentration of BN.
Apparent kinetic parameters for BN N-dealkylation by recombinant CYP3A4 and human liver microsomes
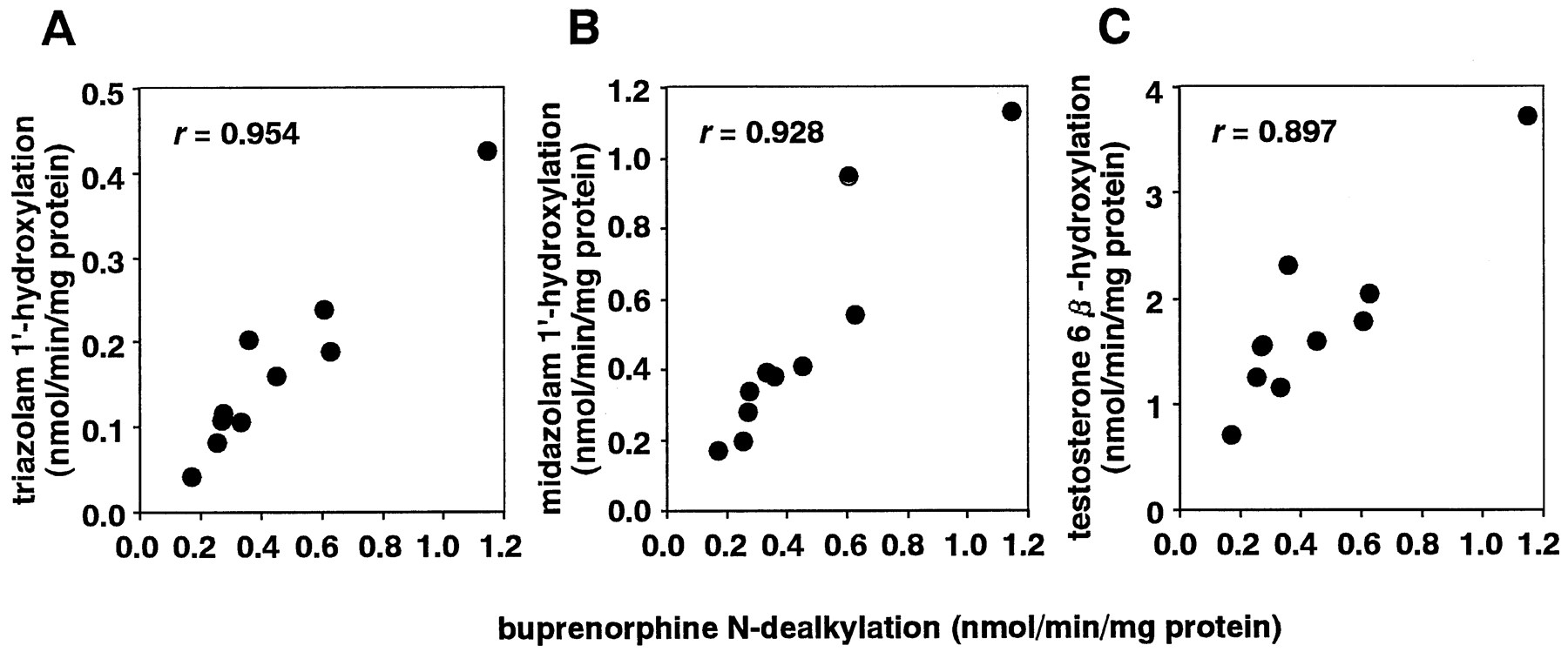
The formation of NBN in human liver microsomes was dependent on NADPH and was completely inhibited by SKF 525-A. These results suggest that BN N-dealkylation is a CYP-dependent metabolic process. Therefore, microsomes from human B lymphoblastoid cells expressing each of eight human CYP enzymes were examined, to clarify the abilities of individual CYP proteins to catalyze BN N-dealkylation. Among the eight recombinant CYP enzymes (CYP1A2, CYP2A6, CYP2B6, CYP2C9, CYP2C19, CYP2D6, CYP2E1, and CYP3A4), only CYP3A4 catalyzed the reaction (2.02 pmol/min/pmol of CYP). The other isoenzymes showed negligible activity for BN N-dealkylation (table2). These data indicated that BNN-dealkylation was catalyzed by CYP3A4. Moreover, the BNN-dealkylation activities showed strong correlations with the CYP3A-specific activities of triazolam 1′-hydroxylation (r = 0.954) (fig.3A), midazolam 1′-hydroxylation (r = 0.928) (fig. 3B), and testosterone 6β-hydroxylation (r = 0.897) (fig.3C). These results suggest that CYP3A4 is the major enzyme involved in BN N-dealkylation by human liver microsomes.
BN N-dealkylation activities in microsomes from human B lymphoblastoid cells expressing CYP enzymes

Correlation of BNN-dealkylation with triazolam 1′-hydroxylation (A), midazolam 1′-hydroxylation (B), and testosterone 6β-hydroxylation (C) in 10 human liver microsomal preparations.
The BN N-dealkylation activities in 10 human liver microsomal preparations were compared with the activities of CYP3A-specific reactions (triazolam 1′-hydroxylation, midazolam 1′-hydroxylation, and testosterone 6β-hydroxylation). The substrate concentrations used were 25 μM for BN, triazolam, and midazolam and 30 μM for testosterone. Assays were performed in duplicate on the same day with the same set of microsomal preparations. Correlation between BN N-dealkylation and each of the three CYP3A-specific metabolic reactions was examined with the least-squares linear regression method.
Recently, Iribarne et al. (1997) reported that BNN-dealkylation in human liver microsomes is mainly catalyzed by CYP3A4; our results are consistent with these findings. We performed additional kinetic studies using recombinant CYP3A4 with the same range of BN concentrations as used in the human liver microsomal studies. The rates of formation of NBN were linear for at least 15 min when 100 μM BN was incubated with recombinant CYP3A4 (0.4 mg/ml protein concentration). Consequently, the kinetic studies with recombinant CYP3A4 were performed at 37°C with a 15-min incubation time and a protein concentration of 0.4 mg/ml. The kinetic parameters estimated for recombinant CYP3A4 for BN N-dealkylation were as follows: apparent KM = 23.7 μM,Vmax = 0.592 nmol/min/mg of protein, andVmax/KM = 25.0 μl/min/mg of protein. The apparent KM value for BN N-dealkylation by recombinant CYP3A4 was close to the mean KM value obtained for human liver microsomes (23.7 vs. 39.3 μM). This finding suggests that the affinity of the human liver microsomes used in the present study for BN N-dealkylation is similar to that of recombinant CYP3A4. This finding supports the concept that BNN-dealkylation by human liver microsomes is mainly catalyzed by CYP3A4.
Among the recombinant enzymes used in this study, B lymphoblastoid cells expressing CYP1A2, CYP2A6, CYP2B6, CYP2C19, or CYP2D6-Met did not coexpress NADPH-CYP reductase. In addition, there were considerable differences in the expression levels for each CYP enzyme, as listed in table 2. Because the activities of the recombinant CYP enzymes are reflected by the expression levels of the CYP enzymes and the NADPH-CYP reductase activities, not only CYP3A4 but also the other CYP enzymes might catalyze BN metabolism. However, correlations between BNN-dealkylation and the reactions metabolized by enzymes except for CYP3A in microsomes from human liver samples (N = 9) were not significant (p> 0.05), as follows: phenacetin O-deethylation (CYP1A2),r = 0.602; coumarin 7-hydroxylation (CYP2A6),r = 0.000; diclofenac 4-hydroxylation (CYP2C9),r = 0.084; (S)-mephenytoin 4-hydroxylation (CYP2C19), r = 0.110; desipramine 2-hydroxylation (CYP2D6), r = 0.617; chlorzoxazone 6-hydroxylation (CYP2E1), r = 0.090. Therefore, we considered that the involvement of enzymes except for CYP3A in BN N-dealkylation is minor.
In conclusion, we have shown that BN is metabolized in human liver microsomes, by a CYP-dependent process, to its N-dealkylated product NBN. The present kinetic study using human liver microsomes and recombinant human CYP enzymes reconfirmed that BNN-dealkylation is primarily catalyzed by CYP3A4 (Iribarneet al., 1997).
Acknowledgment
We thank Katsuhiro Suzuki and Masami Fujii for technical assistance.
Footnotes
-
Send reprint requests to: Prof. Toshinori Yamamoto, Department of Clinical Pharmacy, School of Pharmaceutical Sciences, Showa University, Hatanodai 1–5-8, Shinagawa-ku, Tokyo 142, Japan.
-
This study was supported by a Grant-in-aid for Encouragement of Young Scientists from the Ministry of Education and Science (Grant 06772215), by the Japan Health Science Foundation (Grant 1–7-1-C), and by the Drug Innovation Science Project (Grant 1–2-10) (Tokyo, Japan).
- Abbreviations used are::
- BN
- buprenorphine
- NBN
- norbuprenorphine
- CYP
- cytochrome P450
- Received August 15, 1997.
- Accepted March 14, 1998.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}