


Abstract
Models of inflammation and infection, such as bacterial lipopolysaccharide (LPS), cause suppression of cytochrome P450 expression in various species, although the mechanisms involved are poorly understood. The effects of LPS on expression of phenobarbital (PB)-induced CYP2B1/2 in rats have been well characterized, but less is known about the effects of LPS on PB-induced CYP2B in mice. Since genetically manipulated mice represent an attractive model to study the mechanisms involved in the down-regulation of CYP2B expression by LPS, we investigated the effects of LPS on PB-induced CYP2B expression in mouse liver. Female C57BL/6 mice were injected with 100 mg/kg PB once daily for 4 days to induce CYP2B10 expression, and 1 mg/kg LPS was injected i.p. with the last PB dose. LPS inhibited the mRNA expression of CYP2B10 and CYP2B9 at 6 and 12 h of treatment, with the inhibitory effect more profound at 12 h. LPS also suppressed the CYP2B9 mRNA level at 24 h. However, CYP2B10 mRNA levels in mice treated with PB alone had declined markedly by 24 h after the last PB injection; therefore, no effect of LPS could be discerned. Further experiments showed that injections of 33 mg/kg PB every 8 h produced more stable CYP2B10 mRNA and enzymatic activity. Suppression of CYP2B protein level was found in LPS-treated animals at 24 h of treatment, although no significant effects were noticed at 6 and 12 h of treatment. This study suggests that LPS suppresses the expression of phenobarbital-induced CYP2B expression in mice, which resembles its effects in rats.
Cytochrome P450 (CYP1) superfamily monooxygenases are important for biotransformation of drugs and toxins. Numerous studies indicate that inflammation and infection cause suppression of CYP levels (Morgan, 1997) in various species including human, rat, and mouse. Bacterial endotoxin, or lipopolysaccharide (LPS), has been used extensively as a model of sepsis to study the inhibitory effects of inflammation and infection on CYP activity and expression (Gorodischer et al., 1976; Stanley et al., 1988; Morgan, 1989; Shedlofsky et al., 1994). The effects of LPS on the expression of PB-inducible CYP2B1 and CYP2B2 in rat liver have been well characterized. For example,Khatsenko and Kikkawa (1997) reported that LPS inhibits the phenobarbital-induced expression of CYP2B1/2 in rat liver at both mRNA and protein levels. Moreover, they found that these effects were attenuated in rats treated with inhibitors of nitric-oxide synthase.
Since genetically manipulated mice, including an inducible nitric-oxide synthase knockout mouse (Laubach et al., 1995), are commercially available, the mouse is an attractive model to study the mechanism of CYP regulation during inflammation. To use the genetically manipulated mice to study the mechanisms of regulations, it is necessary to first characterize the effects of LPS on PB-induced mouse CYP2B mRNA and protein expression and on their catalytic activities. Stanley et al. (1988) reported that in male mice, three daily injections of low doses of LPS (80 or 300 ug/kg) slightly potentiated the induction of CYP2B protein and enzymatic activity by PB, while a single higher dose of LPS (1 mg/kg) reduced the CYP2B protein expression and enzymatic activity. Surprisingly, they failed to detect any effects of LPS on the expression of murine CYP2B mRNA using a rat CYP2B1 cDNA probe. Similar experiments have not been done using probes that specifically detect murine CYP2B mRNAs. Moreover, the effects of LPS have not been studied in female mice, which express both CYP2B9 and CYP2B10. For these reasons, in the present study, we investigated the effects of LPS on the mRNA, protein, and catalytic activity of PB-induced CYP2B isoforms in female mouse livers.
Materials and Methods
Animals and Treatments.
C57BL/6 mice (Harlan Sprague-Dawley, Indianapolis, IN) were used in this study. The animals were allowed free access to food and water at all times and were allowed to acclimate to the new environment for 1 week before use.
Experiment 1.
Escherichia coli LPS, serotype 0127:B8, and PB (Sigma Chemical Co., St. Louis, MO) were dissolved in sterile 0.9% saline for use. Groups of six female mice (9–11 weeks old) were injected daily with PB (100 mg/kg, i.p.) for 3 days before the experiment. On the day of the experiment, LPS mice were injected i.p. at 8:00 AM with a mixture of LPS (1 mg/kg) and PB. Control mice were injected with PB, and pair-fed at 6-h intervals with the amount of food that the LPS-treated mice ate. Mice were sacrificed via CO2 asphyxiation at 0, 6, 12, and 24 h after treatments. Mouse livers were harvested and perfused with ice-cold 1.15% KCl before they were used for total liver RNA and microsome protein preparation. These procedures were approved by the Institutional Animal Care and Use Committee of Emory University.
Experiment 2.
Female mice (9–11 weeks old) were injected i.p. with PB by two different dosage regimens: either 33 mg/kg every 8 h or 100 mg/kg every 24 h. Three animals per group were sacrificed every 8 h, and their livers were harvested as described above.
Isolation of Total RNA.
Total liver RNA was prepared by the acid-phenol extraction method according to Chomczynski and Sacchi (1987) and stored at −80°C. Total RNA concentration was determined spectrophotometrically from the absorbances of the samples at 260 nm.
Northern Blotting.
Northern blotting of total mouse liver RNA was performed as described by Sewer et al. (1996). The relative abundances of CYP2B9 and CYP2B10 mRNAs were measured by probing blots with oligonucleotide fragments labeled with 32P using T4 polynucleotide kinase. The oligonucleotide probes were designed to specifically detect CYP2B9 and CYP2B10 in mice. Their sequences were as follows (5′ to 3′): TTC CTT GAA GGT TGG CTC AAC GAC AGC AAC T (CYP2B10) and GTC CTG CAT AAC TAG CTG AGC TGC AGC AAT G (CYP2B9). Blots were hybridized overnight at 50°C using 32P-labeled CYP2B9 or CYP2B10 oligonucleotide probes in a buffer containing 10% formamide, 5× standard saline/phosphate/EDTA, 5× Denhardt's solution, 1% SDS, 0.1 mg/ml yeast tRNA, and 1 mM EDTA. Labeled blots were then washed twice for 30 min at room temperature in a buffer containing standard saline citrate (SSC, 15 mM sodium citrate, pH7.0, and 0.15 M NaCl) and 0.5% SDS, followed by two washes of 30 min at room temperature in a buffer containing 0.2× SSC and 0.1% SDS, and a final wash of 30 min at 45°C (0.1× SSC, 0.5% SDS). Bound32P-labeled probes were detected and quantified by phosphoimaging.
Quantified CYP2B9 and CYP2B10 mRNA levels were normalized to the glyceraldehyde-3-phosphate dehydrogenase (GAP) mRNA content in the same samples, which was measured by probing blots with a cDNA probe of 780 base pairs (American Type Culture Collection, Rockville, MD) (Morgan et al., 1994). A 28S rRNA oligonucleotide probe (AAC GAT GAG AGT AGT GGT ATT TCA CC) was also used to control for loading and transfer artifacts on Northern blots. Each of the oligonucleotide probes and the GAP cDNA probe recognized a single band on Northern blots.
Preparation of Liver Microsomes.
Pyrophosphate-washed microsomes were prepared by differential ultracentrifugation according to Haugen and Coon (1976) and stored at −80°C. Protein concentrations of microsome samples were measured according to the method of Lowry et al. (1951), using bovine serum albumin as a standard.
SDS-PAGE and Western Blotting.
The relative levels of CYP2B proteins were measured via SDS-polyacrylamide gel electrophoresis (PAGE) (Laemmli, 1970) and Western blotting (Towbin et al., 1979). Microsomal proteins were separated via SDS-PAGE (7.5% polyacrylamide) and transferred electrophoretically onto nitrocellulose membranes (Schleicher & Schuell, Keene, NH). Blots were immunolabeled with anti-rat CYP2B1/2 (1:100,000), kindly donated by Dr. James Halpert (University of Texas, Medical Branch at Galveston, TX). Bound antibodies were detected using horseradish peroxidase-coupled goat anti-rabbit IgG and the electrochemiluminescence detection system (Amersham Pharmacia Biotech, Arlington Heights, IL) according to the manufacturer's instructions. The intensities of the bands were quantified via photodensitometry and were proportional to the amount of antigen loaded on the blots.
CYP2B Catalytic Activity.
Pentoxyresorufin O-dealkylase (PROD) activity of the microsomes was used as an indicator of CYP2B catalytic activity of the microsomes (Honkakoski et al., 1992). Reactions were performed using a modified protocol by Burke et al. (1985) in amber plastic microcentrifuge tubes containing 20 to 40 μg of microsomes, 5 mM HEPES buffer, pH 7.6, 20 μM pentoxyresorufin (Sigma Chemical Co.), and 1 mM NADPH in a total volume of 500 μl. After 5 min of incubation at 37°C, reactions were terminated by the addition of 500 μl of acetonitrile. The tubes were centrifuged at 13,000g for 2 min, and 200 μl of supernatant was transferred to a light-shielded 96-well microplate. The amount of resorufin formed was determined by measuring the fluorescence of the samples in a Spectra Max Gemini fluorescence microplate reader (Molecular Devices Corporation, Sunnyvale, CA) at an excitation wavelength of 535 nm and an emission wavelength of 590 nm and by comparison to a resorufin standard curve (Sigma Chemical Co.).
Statistical Analysis.
Quantified data from Northern blots, Western blots, and PROD assays were expressed as a percentage ± S.E.M. of the control group in each experiment. One-way analysis of variance and the Newmann-Keuls post hoc test were used to detect statistically significant differences among the treatment groups. Groups were determined to be significantly different when they had a p value of 0.05 or less.
Results
Specificity of Oligonucleotide Probes and Antibody.
Previous work has shown that CYP2B10 is expressed in the livers of both male and female mice. Although its basal level is low, CYP2B10 is greatly induced by PB treatment. On the other hand, CYP2B9 is female-specific; no CYP2B9 was detected in male mouse liver (Honkakoski et al., 1992). To verify the specificities of the designed CYP2B9 and CYP2B10 oligonucleotide probes, we analyzed mouse liver mRNAs from control and PB-treated animals of both sexes by Northern blotting. The CYP2B10 oligonucleotide probe detected a single band in both male and female PB-treated mice (Fig. 1A). The level of this mRNA in PB-treated mice was much higher than that in control animals, which was barely detectable. The CYP2B9 oligonucleotide probe also detected a single band in control female mouse livers (Fig. 1A), whereas no signal was detected in either control or PB-treated male mice. These results indicated that the designed oligonucleotide probes were specific for CYP2B10 and CYP2B9 mRNA.

Induction of CYP2B9 and CYP2B10 mRNA and CYP2B protein by PB in male and female mice livers.
Male or female C57BL/6 mice, 9 to 11 weeks old, were injected with PB (33 mg/kg, i.p.) every 8 h for 24 h before they were killed for analysis. Control mice were injected with saline. A, CYP2B9 and CYP2B10 mRNA were analyzed via Northern blotting. Blots of 20 μg of mouse liver total RNA were probed with 32P-labeled oligonucleotides designed specifically for CYP2B9 and CYP2B10 mRNA. Ribosomal RNA content in each sample was used as internal control. B, 0.6 μg of mouse liver microsomes were resolved by SDS-PAGE, and CYP2B protein levels were analyzed by Western blotting. Blots were immunolabeled with anti-rat CYP2B1/2 as described in the text. C, control mice; PB, PB-treated mice.
We also tested the specificity of the anti-rat CYP2B1/2 antibody, which we used for mouse CYP2B protein detection on Western blots. Results are shown in Fig. 1B. A major band with apparent molecular weight of 57,000 was detected in microsomes from both male and female mouse livers. Its intensity was higher in untreated females than in males, and it was induced by PB treatment. Therefore, we believe that this band contains both CYP2B9 and -2B10 in PB-treated females. A PB-inducible minor band with an apparent molecular weight of 53,000, the level of which was similar in male and female mice, was also detected. Honkakoski et al. (1992) also detected two protein bands in murine liver microsomes, with apparent molecular masses of 56 and 53 kDa. They concluded that the 53-kDa band was CYPB9, on the basis of its sex dependence and hormonal regulation in DBA/2 mice. However, the 53-kDa band observed on our blots is highly PB-inducible and not female-specific (in contrast to CYP2B9 mRNA, Fig. 1A), making it highly unlikely that it is CYP2B9. These differences between our observations and those of Honkakoski et al. (1992) may be attributable to the different relative mobilities of P450 enzymes under slightly different electrophoresis conditions (Okita et al., 1997) and/or the different strains of mice used. From the gene sequences, CYP2B9 and CYP2B10 proteins are predicted to have 491 and 500 amino acid residues, respectively (Lakso et al., 1991), with corresponding predicted molecular weights of 55,740 and 56,742.
Effects of LPS on CYP2B mRNA Expression.
The above data verified that the constitutive expression of CYP2B10 mRNA is at a low level, although the CYP2B9 mRNA as well as CYP2B protein are easily detected in untreated female mouse livers. Therefore, to study the regulation of CYP2B10 as well as -2B9, we sought an appropriate PB induction protocol. Two routes are commonly used to administer PB to rodents for induction of CYP isoforms: in the drinking water or via i.p. injection. LPS treatment causes reduced eating and drinking activities in mice (Kozak et al., 1994), and thus to ensure that every mouse received the appropriate amount of PB during the course of the experiment, we were compelled to administer it by injection. We used three daily i.p. injections of PB (100 mg/kg each) in our initial study, which is the most widely used injection protocol (Jarukamjorn et al., 1999).
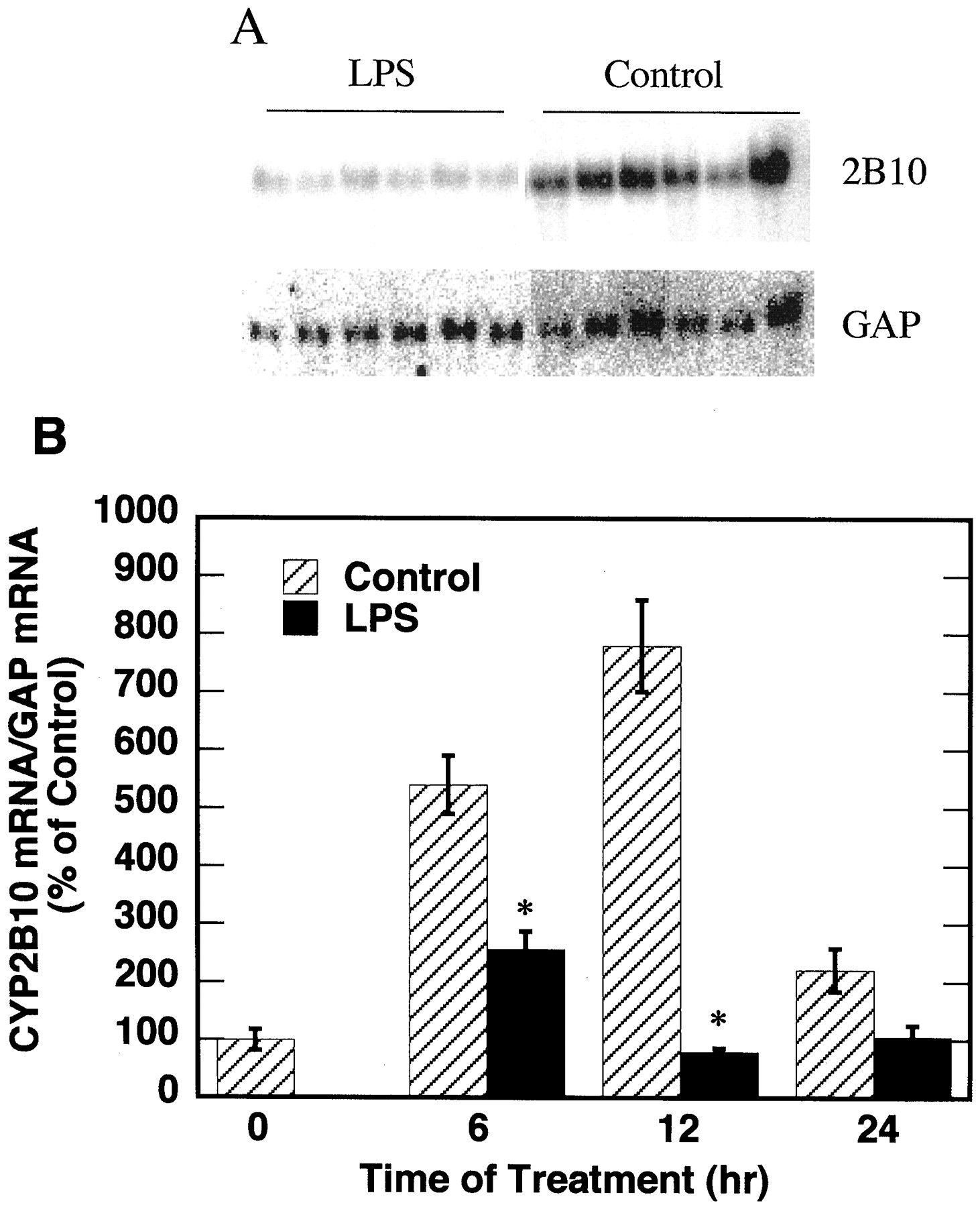
Mice were injected with PB daily for 3 days before the experiment. On the day of experiment, mice were injected with a mixture of LPS (1 mg/kg) and PB, or PB alone. Control mice were pair-fed with the LPS-treated animals. The effects of LPS on the expression of CYP2B10 mRNA are shown in Fig. 2. In animals treated with PB alone, CYP2B10 mRNA level was induced at 6 and 12 h after the last dose of PB and significantly decreased 24 h after PB injection. CYP2B10 mRNA in LPS-treated mice remained at relatively low levels at all three time points tested. Consequently, at 6 and 12 h of treatment, LPS significantly suppressed the expression of CYP2B10 mRNA in comparison with the respective control groups. However, after 24 h of treatment, the CYP2B10 mRNA levels in control mice significantly decreased, and no significant effect of LPS was detected (Fig. 2).

Effect of LPS on the expression of CYP2B10 mRNA.
Female C57BL/6 mice (8–10 weeks old) were injected daily with PB (i.p., 100 mg/kg) for 3 days before the experiment. On the day of experiment, LPS mice were injected with a mixture of LPS (1 mg/kg) and PB in sterile saline. Control mice were injected with PB in sterile saline and pair-fed with the amount of food that the LPS-treated mice ate, divided into 6-h intervals. Mice were sacrificed at 0, 6, 12, and 24 h after treatments and their livers were harvested for RNA and microsomal protein preparation. A, Northern blotting results at 12 h of treatment (20 μg of total RNA per sample). B, quantitative analysis of Northern blot data, normalized to the GAP mRNA level in the same samples. The CYP2B10 mRNA level at 0 h was assigned as 100%. There were six animals in each treatment group. *Significantly different from control, p < 0.05
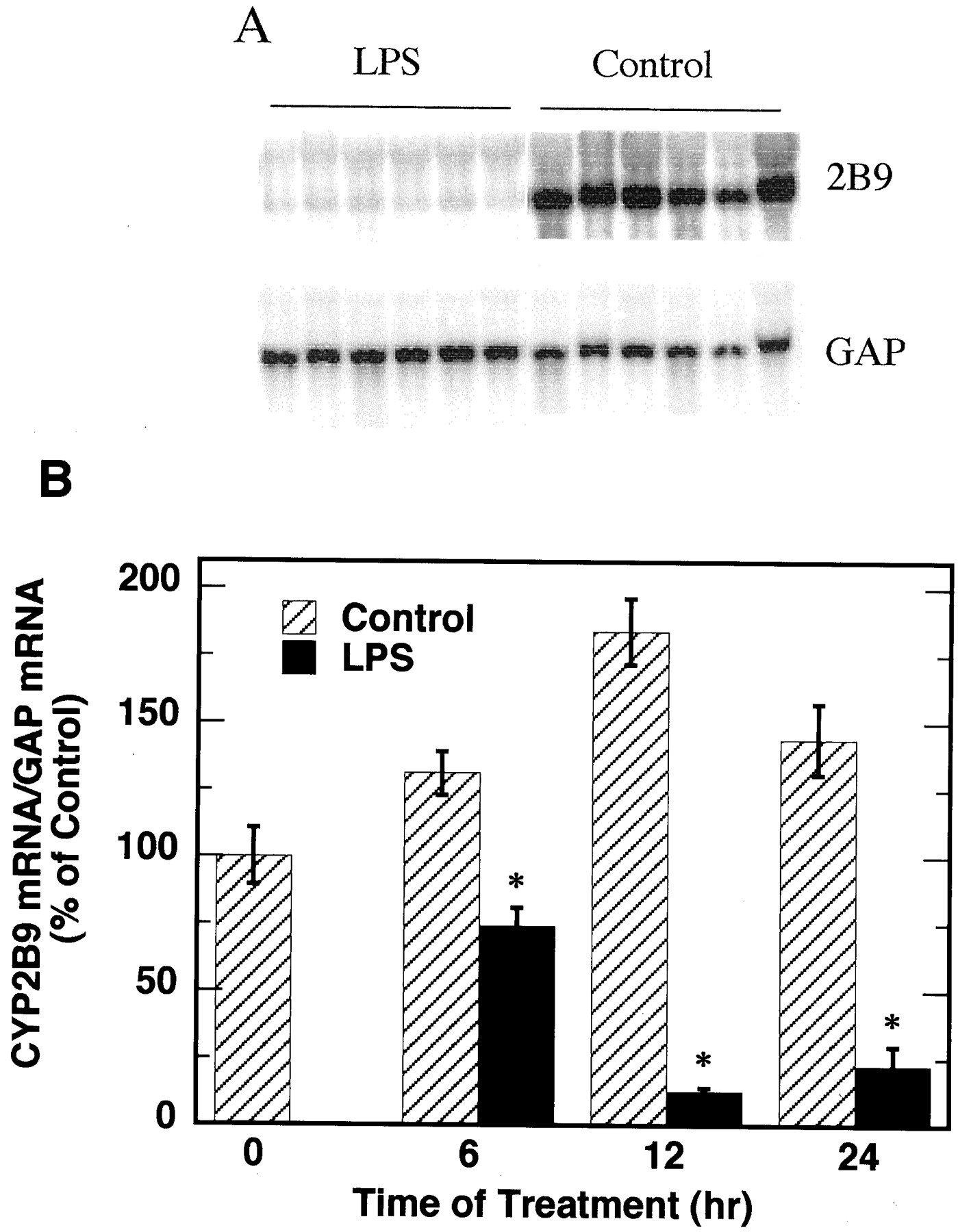
The effects of LPS on the expression of CYP2B9 mRNA were also studied (Fig. 3). CYP2B9 mRNA levels in control mice were significantly induced at 12 and 24 h after last PB injection, although they fluctuated much less than the respective CYP2B10 mRNA levels throughout the 24 h of experiment. LPS significantly suppressed the expression of CYP2B9 at all three time points measured.

Effect of LPS on the expression of CYP2B9 mRNA.
Animals were treated as described in Fig. 2. A, Northern blotting results at 12 h of treatment (20 μg of total RNA per sample). B, quantitative analysis of Northern blot data, normalized to the GAP mRNA level in the same samples. The CYP2B9 mRNA level at 0 h was assigned as 100%. There were six animals in each treatment group. *Significantly different from control, p < 0.05
Effects of LPS on Microsomal CYP2B Protein Levels.
We also studied the effects of LPS on mouse microsomal CYP2B protein levels by Western blotting. As shown in Fig.4, LPS significantly suppressed the expression of CYP2B protein at 24 h of treatment, although no significant effect was detected at 6 and 12 h.
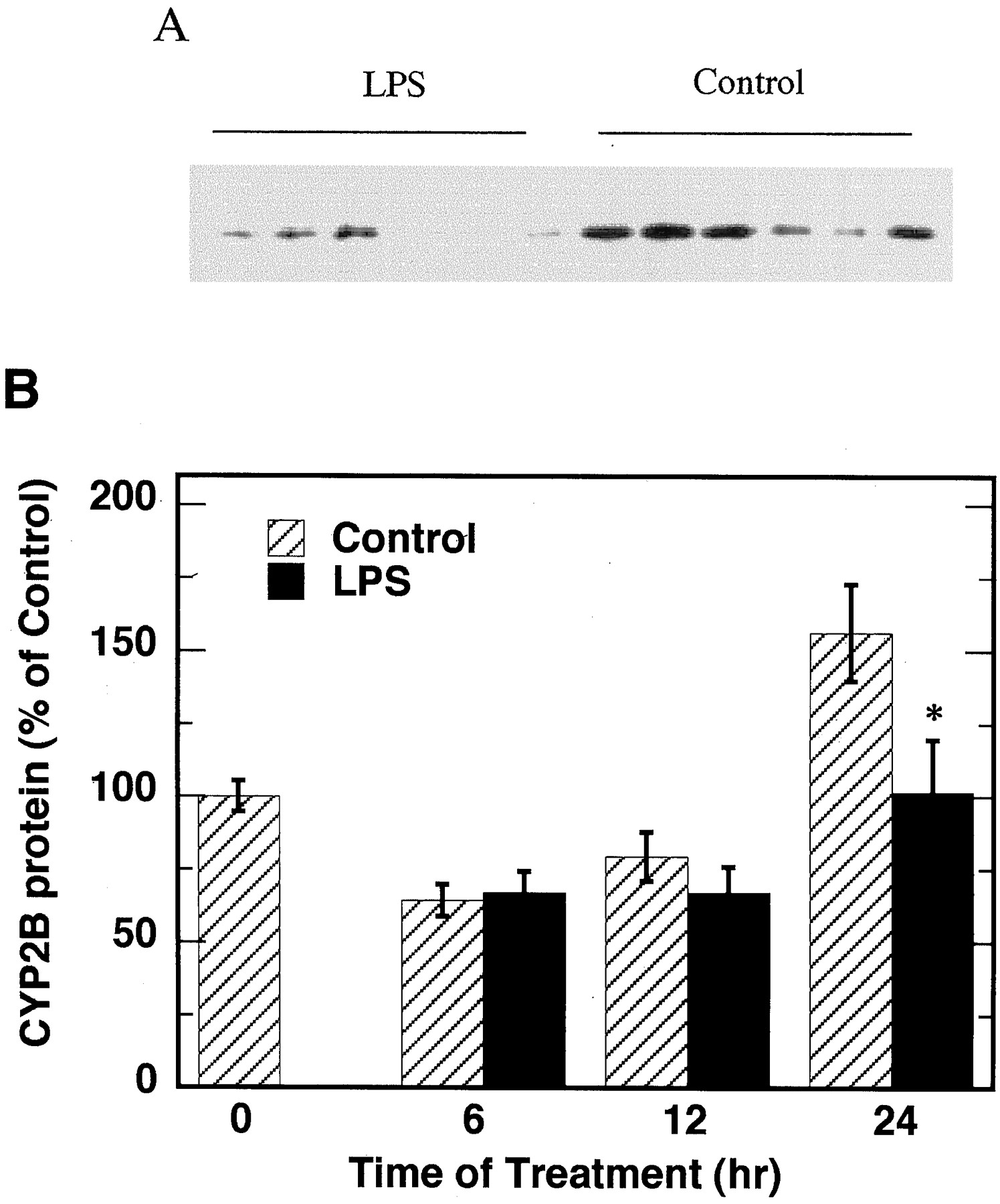
Effect of LPS on microsomal CYP2B protein level.
Animals were treated as described in Fig. 2. A, Western blotting results at 24 h of LPS treatment (0.6 μg of microsomal protein per sample). B, quantitative analysis of Western blot data. The CYP2B protein level at 0 h was assigned as 100%. There were six animals in each treatment group. *Significantly different from control,p < 0.05
Effect of PB Dosage Interval on Induction of CYP2B mRNA.
The above results showed that the CYP2B10 mRNA level in mice treated with only PB decreased dramatically 24 h after the last PB injection, compared with the 6- and 12-h groups. Therefore, we investigated the effect of dosage interval on the expression pattern of PB-induced CYP2B10 mRNA, with the goal of establishing a dose regimen that produces a stable CYP2B10 mRNA expression level in mice. In this study, 8- to 10-week-old C57BL/6 mice were injected with PB either at 33 mg/kg every 8 h or at 100 mg/kg every 24 h.
The effects of PB dosage interval on induction of CYP2B10 mRNA in mouse livers are shown in Fig. 5A. CYP2B10 mRNA levels in both groups of mice were significantly induced by PB treatment. Daily injection of PB produced the highest CYP2B10 mRNA levels at 16 and 8 h after the first and second injection, respectively. However, there existed rapid declines of CYP2B10 mRNA following these peaks. Injections of 33 mg/kg PB every 8 h produced more stable CYP2B10 mRNA levels than daily injections of 100 mg/kg PB. A maximal and relatively stable level of induction was reached after three injections of 33 mg/kg PB (one every 8 h), although the mean steady-state level was lower than the highest levels produced by daily injection of 100 mg/kg PB.

Effect of PB dosage interval on induction of CYP2B10 and -2B9 mRNAs.
Female C57BL/6 mice were injected with PB either at 33 mg/kg every 8 h or at 100 mg/kg every 24 h. Three animals per group were sacrificed every 8 h for analysis. Relative levels of CYP2B10 (A) and CYP2B9 (B) mRNAs in the samples were measured by Northern blotting and normalized to the GAP mRNA levels of the samples. Values were expressed as a percentage of levels in untreated animals (time 0).
We also studied the effects of PB dosage interval on expression of CYP2B9 mRNA. We found that CYP2B9 is constitutively expressed at a relatively high level in female mice but is significantly induced (2- to 3-fold) by PB treatment (Fig. 5B). As for CYP2B10, injections of 33 mg/kg PB every 8 h produced more stable CYP2B9 mRNA levels than daily injections of 100 mg/kg PB.
Effects of PB Dosage Interval on Induction of CYP2B Protein and PROD Activity.
In addition to studying mRNA expression levels, we also studied the effects of PB dosage interval on induction of CYP2B protein via Western blotting, and its catalytic activity using PROD as an indicator (Fig.6). As shown in Fig. 6A, PB treatments significantly induced CYP2B protein levels in both groups of female mice. In comparison with the induction pattern of CYP2B10 mRNA (Fig.5A), the induction of CYP2B protein was much slower and did not reach a maximum and stable level until after 40 h of treatment, probably reflecting a longer half-life of CYP2B protein. Injections of 33 mg/kg PB every 8 h produced a slower induction of CYP2B protein than did daily injections of 100 mg/kg PB, but the maximal induction by each regimen was similar.

Effect of PB dosage interval on induction of CYP2B protein and PROD activity.
Animals were treated as described in Fig. 5. A, relative levels of microsomal CYP2B protein were measured by Western blotting. B, PROD activities of the microsomes were measured as described in the text. Values are expressed as a percentage of levels in untreated animals (time 0).
The patterns of PROD activity induction by PB (Fig. 6B) resembled those of CYP2B protein induction (Fig. 6A). At the 24-h dose interval, PROD activities and CYP2B protein levels fluctuated less than CYP2B10 mRNA (Fig. 5).
Discussion
This study found that LPS suppresses PB-induced CYP2B10 and CYP2B9 mRNA expression in female mouse liver at 6 and 12 h of treatment, similar to its effects on rat CYP2B1/2 (Khatsenko and Kikkawa, 1997). This is important because it allows us to further investigate the mechanisms of this LPS-induced down-regulation of the PB-inducible CYP2B isoforms using genetically manipulated mouse models. LPS treatment caused a significant decrease in CYP2B protein levels at 24 h but not at 6 or 12 h of treatment, indicating that the decrease in CYP2B protein was probably a result of pretranslational suppression due to the earlier decrease in CYP2B mRNAs.
Our results showing a down-regulation of PB-induced CYP2B protein 24 h after LPS injection are in agreement with the earlier work byStanley et al. (1988). However, these investigators reported that 1 mg/kg LPS did not affect the levels of mouse liver mRNAs detected on Northern blots using a rat CYP2B1 mRNA probe (Stanley et al., 1988). In contrast, we detected a clear suppression of CYP2B10 and -2B9 mRNAs by LPS treatment; the discrepancy is probably explained by the interference of P450s other than CYP2B9 or -2B10 in the blots probed with the rat cDNA, and by the fact that they only studied mRNA expression 24 h after the last dose of PB. It is likely based on our findings that levels of CYP2B10 mRNA in their control (PB-treated) mice were already quite low. This can not be ascertained because the results were not shown. Another possible contributing factor could be the different strains and sexes of the mice used. Stanley et al. (1988)used male CBA mice, while we used female C57/BL6 mice because only females express CYP2B9.
Multiple daily injections of PB is a standard protocol used to induce mouse CYP2B expression to a relatively stable level, possibly because early investigations focused on CYP2B protein expressions and catalytic activities, which do not fluctuate as much as CYP2B10 mRNA levels. In this study, we found that this method resulted a rapid decline of CYP2B10 mRNA between 12 and 24 h after the last dose, suggesting a rapid clearance of PB and a short half-life of the CYP2B10 mRNA.
In rats, the maximal induction in CYP2B mRNA levels after a single injection of PB was observed at 16 h, and the induced CYP2B mRNA remained elevated after 24 h of treatment (Hardwick et al., 1983). The sustained rat CYP2B mRNA level after PB treatment was different from the PB-induced mouse CYP2B10 mRNA kinetics in this study, suggesting a species-dependent difference in regard to mRNA half-life. Another factor that could have contributed to this difference is a difference in rates of PB metabolism in mice and rats. It was reported that the PB half-life in rats was approximately 9 h (Pelkonen and Karki, 1973). However, little is known about the PB half-life in mice.
Results from the second experiment indicated that injections of PB at 33 mg/kg every 8 h are better than daily injections at 100 mg/kg in achieving steady levels of CYP2B mRNA expression in mouse liver, and that three injections (one every 8 h) are sufficient to produce a maximum and steady CYP2B mRNA level. These findings will be critical for investigators who wish to investigate the modulation of PB-inducible CYP2B expression by inflammatory stimuli or other factors in mice. However, CYP2B protein and PROD levels in mice receiving daily PB injections fluctuated much less, and reached stable levels much more slowly, than did CYP2B10 mRNA. These observations most likely reflect a longer half-life of CYP2B protein than CYP2B10 mRNA. The half-life of murine CYP2B proteins is not known, but the in vivo half-life of rat CYP2B1 is estimated to be more than 30 h (Shiraki and Guengerich, 1984). Taken together, our observations indicate that the PB dosage regimen is not as critical for studies of CYP2B10 protein and activity as it is for studies on the mRNA
Work by Negishi and coworkers (Honkakoski et al., 1998; Kawamoto et al., 1999) indicated that the constitutively active receptor (CAR), a transcription factor, is involved in the induction of CYP2B mRNA expression. PB treatment rapidly increases the nuclear translocation and binding of CAR to the phenobarbital-responsive enhancer module of the CYP2B10 gene in mouse livers and primary mouse hepatocytes (Kawamoto et al., 1999). The cyclic induction of CYP2B10 mRNA levels in mice treated with PB every 24 h suggests that CAR activation by PB in the mouse is also cyclic, and that the receptor does not undergo long-term desensitization in vivo. Therefore, it would be interesting to investigate the effects of PB dose interval and LPS treatment on the nuclear translocation and binding of CAR to the phenobarbital-responsive enhancer module.
Honkakoski et al. (1992) reported that whereas CYP2B10 was greatly induced by PB treatment of female DBA/2N mice, CYP2B9 mRNA was not affected. In this study, we consistently observed a small induction of CYP2B9 mRNA in animals treated with PB (Figs. 1, 3, and 5). This is unlikely to be due to cross-reactivity of the CYP2B9 probe with the major PB-inducible form, CYP2B10, since the CYP2B9 probe failed to detect the CYP2B10 induced by PB in male mouse liver (Fig. 1A). Our results are in agreement with those of Jarukamjorn et al. (1999), using male or female C57BL/6NCrj mice, who further showed that PB can induce CYP2B9 in murine hepatocyte cultures. The reasons for the contradictory reports from different laboratories are not known, but since both we and Jarukamjorn et al. (1999) used C57BL/6 mouse strains, it is tempting to speculate that the contradictions may be due to a genetically determined strain difference.
Footnotes
-
Send reprint requests to: Edward T. Morgan, Ph.D., Dept. of Pharmacology, Emory University Atlanta, GA 30322. E-mail:etmorga{at}bimcore.emory.edu
- Abbreviations used are::
- CYP or P450
- cytochrome P450
- PB
- phenobarbital
- CAR
- constitutively active receptor
- GAP
- glyceraldehyde-3-phosphate dehydrogenase
- LPS
- bacterial lipopolysaccharide
- PROD
- pentoxyresorufin O-dealkylase
- SSC
- standard saline citrate
- Received October 11, 2000.
- Accepted November 17, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}