


Abstract
Most known interactions between herbal extracts and drugs involve the inhibition of drug-metabolizing enzymes, but little is yet known about the possible role of transporters in these interactions. In this study, we have examined the effects of herbal extracts used in dietary supplements on the function of organic anion-transporting polypeptide B (OATP-B; OATP2B1), which is expressed on human intestinal epithelial cells and is considered to be involved in the intestinal absorption of various drugs. Specifically, the effects of 15 herbal extracts on uptake of estrone-3-sulfate, a typical OATP-B substrate, by human embryonic kidney 293 cells stably expressing OATP-B were evaluated. At concentration levels considered likely to be attainable in the human intestine, extracts of bilberry, echinacea, green tea, banaba, grape seed, ginkgo, and soybean potently inhibited estrone-3-sulfate uptake by 75.5, 55.5, 82.1, 61.1, 64.5, 85.4, and 66.8%, respectively (P < 0.01). The inhibitory effect of ginkgo leaf extract was concentration-dependent (IC50 = 11.2 ± 3.3 μg/ml) and reversible. Moreover, flavonol glycosides and catechins significantly inhibited the function of OATP-B, suggesting that the inhibitory effects of the herbal extracts on OATP-B may be primarily attributable to flavonoids. The extracts of mulberry, black cohosh, and Siberian ginseng moderately (but significantly) inhibited estrone-3-sulfate uptake by 39.1, 47.2, and 49.2%, respectively (P < 0.05). Extracts of barley, Job's tears, rutin, rafuma, and passionflower were ineffective. These results suggest that coadministration of some dietary supplements may decrease the absorption of orally administered substrates of OATP-B.
It has been reported that 42.1% of people questioned in the United States were taking dietary supplements, and 18.4% were taking prescribed drugs together with herbal dietary products (Eisenberg et al., 1998). Likewise, 73.1% of Italian cancer patients were taking the prescribed drugs concomitantly with dietary supplements (Molassiotis et al., 2005). Therefore, detailed information about possible interactions between drugs and dietary supplements is needed.
Besides phase I drug-metabolizing enzymes such as cytochrome P450 (P450) and phase II drug-metabolizing enzymes such as sulfotransferase, ATP-binding cassette transporters such as P-glycoprotein (P-gp) and multidrug resistance-associated protein 2 are expressed on the intestinal epithelial cells, where they serve to limit the uptake of xenobiotics (Thiebaut et al., 1987; Suzuki and Sugiyama, 1998; Mottino et al., 2000). Many other transporters are also expressed on the intestinal epithelial cells and mediate the uptake of various compounds. It has been reported that organic anion-transporting polypeptide B (OATP-B; OATP2B1, gene symbol SLC21A9/SLCO2B1), a member of the organic anion-transporting polypeptide family, mediates the transport of anionic compounds, such as estrone-3-sulfate, sulfobromophthalein, and dehydroepiandrosterone sulfate (DHEAS), and is considered to be involved in the intestinal absorption of anionic drugs in humans (Tamai et al., 2000; Kullak-Ublick et al., 2001; Kobayashi et al., 2003).
Recently, citrus juices, including grapefruit juice (GFJ), have been reported to reduce the bioavailability of orally administered fexofenadine, an antihistamine. This interaction is considered to be caused by the inhibition of intestinal OATPs (Banfield et al., 2002; Dresser et al., 2002). Indeed, we have reported that GFJ and orange juice inhibit the function of OATP-B in vitro (Satoh et al., 2005). Therefore, OATPs expressed in the intestine are quite likely to be involved in citrus juice-drug interactions.
Some herbal products affect the pharmacokinetics of certain drugs, although previous reports have focused mainly on herb-drug interactions involving modulation of the function of drug-metabolizing enzymes. For example, repetitive administration of St. John's wort decreases the blood concentrations of certain drugs, such as immunodepressants, by the induction of P450s (Piscitelli et al., 2000; Bauer et al., 2003). Some herbal extracts, such as ginkgo, echinacea, and soybean extracts, have been reported to inhibit P450s in vitro (He and Liu, 2002; Anderson et al., 2003; He and Edeki, 2004). We have reported that beverages such as green tea, which is processed into dietary supplements, inhibit the function of sulfotransferase (Nishimuta et al., 2005).
On the other hand, the interaction of herbal dietary products with transporters has recently received increasing attention. For example, repetitive administration of St. John's wort induces the expression of not only P450s but also P-gp and decreases the bioavailability of its substrates, such as indinavir, cyclosporin A, and digoxin (Johne et al., 1999; Durr et al., 2000; Perloff et al., 2001). Extracts of green tea, garlic, and milk thistle inhibit the function of P-gp in vitro (Jodoin et al., 2002; Zhang and Morris, 2003; Zhou et al., 2004), and genistein, one of the ingredients of soybean, inhibits the functions of both P-gp and multidrug resistance-associated protein 2 (Jager et al., 1997; Ciolino et al., 1999). However, few studies have been conducted to elucidate the effects of herbal ingredients on the transporters that function to take up drugs into the intestinal epithelial cells. In this study, we examined the effects of 15 herbal extracts on the function of OATP-B, which is expressed in the intestine, to investigate whether novel herb-drug interactions might exist.
Materials and Methods
Materials. [3H]Estrone-3-sulfate ammonium salt (2.12 TBq) and [3H]glibenclamide (1.65 TBq) were purchased from PerkinElmer Life and Analytical Sciences (Boston, MA). Ginkgolide A, ginkgolide B, bilobalide, quercetin-3-rhamnoside, and quercetin-3-rutinoside were purchased from Sigma-Aldrich (St. Louis, MO). Kaempferol-3-glucoside, kaempferol-3-rutinosid, isorhamnetin-3-glucoside, (+)-catechin, (–)-epicatechin (EC), (–)-epigallocatechin (EGC), (–)-epicatechin gallate (ECG), and (–)-epigallocatechin gallate (EGCG) were purchased from Extrasynthese S.A. (Genay, France). Ginkgolide C was purchased from Tokiwa Phytochemical Co. (Chiba, Japan). The herbal extracts (Table 1) used in this study were kind gifts from Tokiwa Phytochemical Co. All other chemicals were commercial products of reagent grade.
Scientific name, parts used, factor, solvent used for extraction, typical ingredient(s), putative clinical applications, and recommended daily dose of test compounds
Uptake Experiments. Human embryonic kidney (HEK) 293 cells stably expressing OATP-B (HEK/OATP-B cells) and HEK293 cells transfected with vector alone (HEK/Mock cells) were previously constructed (Satoh et al., 2005). Cells were cultured in Eagle's minimal essential medium (Nissui Pharmaceutical Co., Ltd., Tokyo, Japan) supplemented with 10% fetal calf serum (Biofluids Inc., Rockville, MD), 2 mM l-glutamine (Sigma-Aldrich), 70 μg/ml penicillin (Wako Pure Chemical Industries, Ltd., Osaka, Japan), 100 μg/ml streptomycin (Nacalai Tesque Inc., Kyoto, Japan), 500 μg/ml geneticin (G418; Sigma-Aldrich), and 2.2 mg/ml NaHCO3 at 37°C under 5% CO2/95% air. For uptake experiments, the cells were seeded at a density of 4 to 8 × 104 cells per well on 96-well microwell plates (Nalgen NUNC International, Rochester, NY). After cultivation for 2 days, the culture medium was replaced with fresh Eagle's minimal essential medium as above, but without G418. On the next day, the uptake study was performed. The culture medium was removed, and the cells were washed three times with 100 μl of uptake buffer (125 mM NaCl, 4.8 mM KCl, 5.6 mM d-glucose, 1.2 mM CaCl2, 1.2 mM KH2PO4, 1.2 mM MgSO4, and 25 mM HEPES, pH 7.4; 37°C) and then preincubated with 200 μl of buffer at 37°C for 10 min. After the preincubation, the buffer was replaced with 100 μl of buffer containing a radiolabeled compound to initiate the uptake reaction. After incubation for the designated times, 200 μl of ice-cold buffer was added to the well to stop the uptake reaction, and immediately, the cells were washed three times with 100 μl of ice-cold buffer. The cells were solubilized in 200 μl of 1 N NaOH over 4 h and then neutralized with 100 μl of 2 N HCl. A liquid scintillation cocktail (Clear-sol I; Nacalai Tesque) was added to 200-μl aliquots of the solubilized cells, and the radioactivity was determined by using a liquid scintillation counter (LSC-3500; Aloka Co., Ltd., Tokyo, Japan). The content of protein in the solubilized cells was measured by Lowry's method (Lowry et al., 1951), with bovine serum albumin as a standard.
Inhibition Experiments. Extracts of barley, bilberry, echinacea, green tea, mulberry, and Job's tears were dissolved in uptake buffer. Extracts of black cohosh, rutin, rafuma, banaba, passionflower, grape seed, ginkgo, Siberian ginseng, and soybean were dissolved in dimethylsulfoxide (DMSO) and diluted with uptake buffer to give a 0.5% final DMSO concentration. In this case, the buffer containing 0.5% DMSO was used as the control. The concentration of herbal extracts for the inhibition study was determined by dividing the recommended daily dose (Table 1) by the estimated volume of gastrointestinal fluid (1 liter), and this taken as the putative gastrointestinal concentration. Daily dosages of herbal extracts were defined by referring to the recommendation in the Physicians' Desk Reference for herbal medicines (Gruenwald et al., 2000) or the recommendation by manufacturers of dietary supplements in Japan. The buffer was adjusted to pH 7.4 and approximately 300 mOsm. Each extract was added simultaneously with the radiolabeled substrate in the uptake experiments.
We investigated whether the inhibition of OATP-B-mediated uptake by ginkgo leaf extract is reversible or not. Cells were preincubated with 1, 10, or 100 μg/ml ginkgo leaf extract for 10 min. Then cells were washed five times with uptake buffer before the addition of 10 nM [3H]estrone-3-sulfate. Uptake experiments were performed according to the procedure described under Uptake Experiments.
After these experiments, we investigated the effects of ginkgo leaf extract constituents, terpenoids (ginkgolide A, ginkgolide B, ginkgolide C, and bilobalide), and flavonol glycosides (kaempferol-3-glucoside, kaempferol-3-rutinoside, quercetin-3-rhamnoside, quercetin-3-rutinoside, and isorhamnetin-3-glucoside), and green tea extract constituents, (+)-catechin, EC, EGC, ECG, and EGCG. Each compound was dissolved in DMSO, diluted with buffer to give 0.5% final DMSO concentration, and added at a concentration of 1, 10, or 100 μM simultaneously with the radiolabeled substrate in the uptake experiments.
Data Analysis. The uptake value (microliters per milligram of protein) was obtained by dividing the radioactivity taken up into the cells per cellular protein content by the concentration of radioactivity of the test compound in the buffer. The OATP-B-mediated uptake was evaluated after subtracting the initial uptake of estrone-3-sulfate (20 s) or glibenclamide (1 min) into HEK/Mock cells from that into HEK/OATP-B cells. Inhibitory effect (percentage of control) was evaluated as the ratio of the OATP-B-mediated uptake in the presence of the herbal extracts or constituents to that in the absence of the herbal extracts or constituents (control).

Time courses of the uptake of estrone-3-sulfate (A) or glibenclamide (B) by human OATP-B. The uptakes of 10 nM [3H]estrone-3-sulfate or 10 nM [3H]glibenclamide into HEK/OATP-B cells (closed circles) and HEK/Mock cells (open circles) were measured at 37°C and pH 7.4. Each point represents the mean ± S.E.M. of four experiments.
To determine the half-maximal inhibitory concentration (IC50; micrograms per milliliter) of the extracts for the uptake of estrone-3-sulfate, following equation was fitted to the observed date for the OATP-B-mediated uptake in the presence of various concentrations of ginkgo leaf and green tea extracts by using a nonlinear least-squares regression analysis program, MULTI (Yamaoka et al., 1981); V/V0 = 1 – [I]/(IC50 + [I]), where V and V0 are the OATP-B-mediated uptake rates in the presence or absence of the extracts, respectively, and [I] is the concentration of the extracts (micrograms per milliliter). In the statistical analysis, the significance of differences between the mean values was determined by analysis of variance (ANOVA), followed by Dunnett's test, and a p value of less than 0.05 was considered statistically significant.
Results
OATP-B-Mediated Uptake of Estrone-3-Sulfate or Glibenclamide. The uptake of 10 nM [3H]estrone-3-sulfate or [3H]glibenclamide into HEK/OATP-B cells was significantly higher than that into HEK/Mock cells (Fig. 1). Because this uptake increased linearly over 20 s (estrone-3-sulfate) or 1 min (glibenclamide), the initial uptake rate was determined at 20 s or 1 min, respectively.
Effects of Herbal Extracts on OATP-B-Mediated Uptake of Estrone-3-Sulfate. The extracts of bilberry, echinacea, green tea, banaba, grape seed, ginkgo, and soybean at the putative gastrointestinal concentration potently inhibited the function of OATP-B by 75.5, 55.5, 82.1, 61.1, 64.5, 85.4, and 66.8%, respectively (P < 0.01). The extracts of mulberry, black cohosh, and Siberian ginseng moderately, but significantly, inhibited the function of OATP-B by 39.1, 47.2, and 49.2%, respectively (P < 0.05). The extracts of barley, Job's tears, rutin, rafuma, and passionflower at the putative gastrointestinal concentrations did not significantly inhibit uptake (Fig. 2).
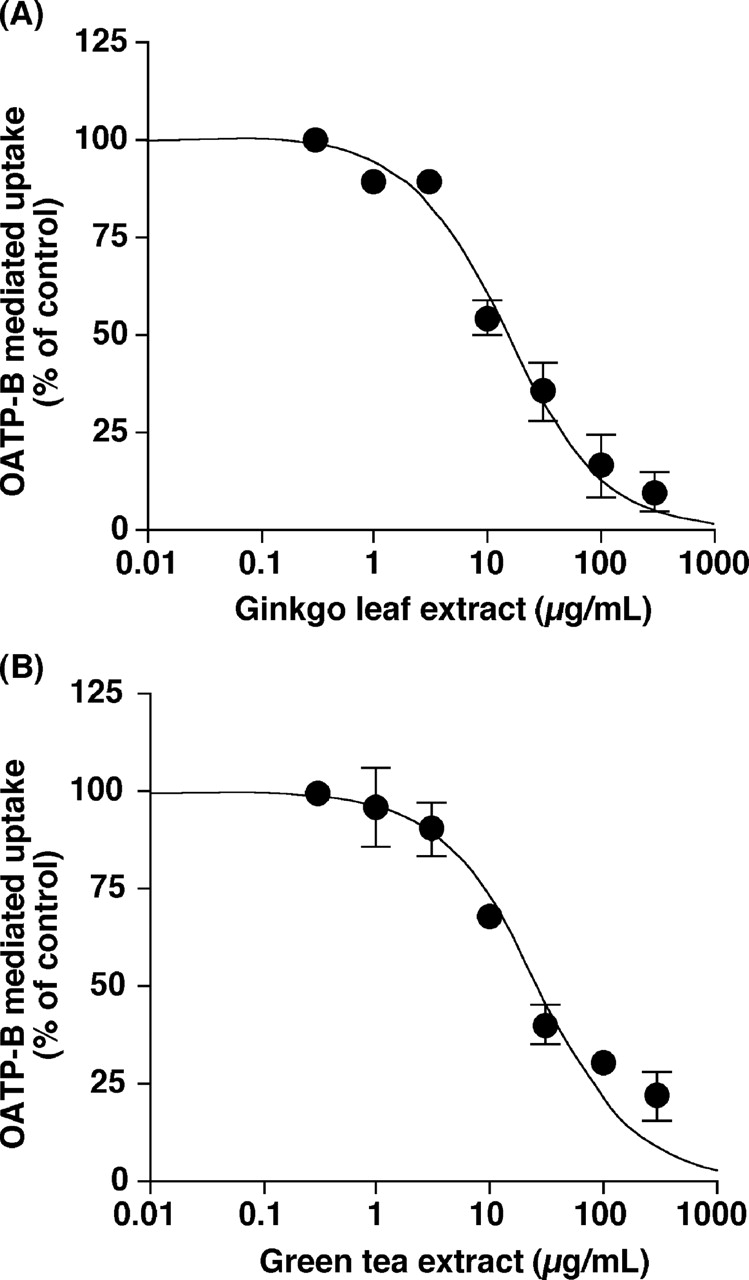
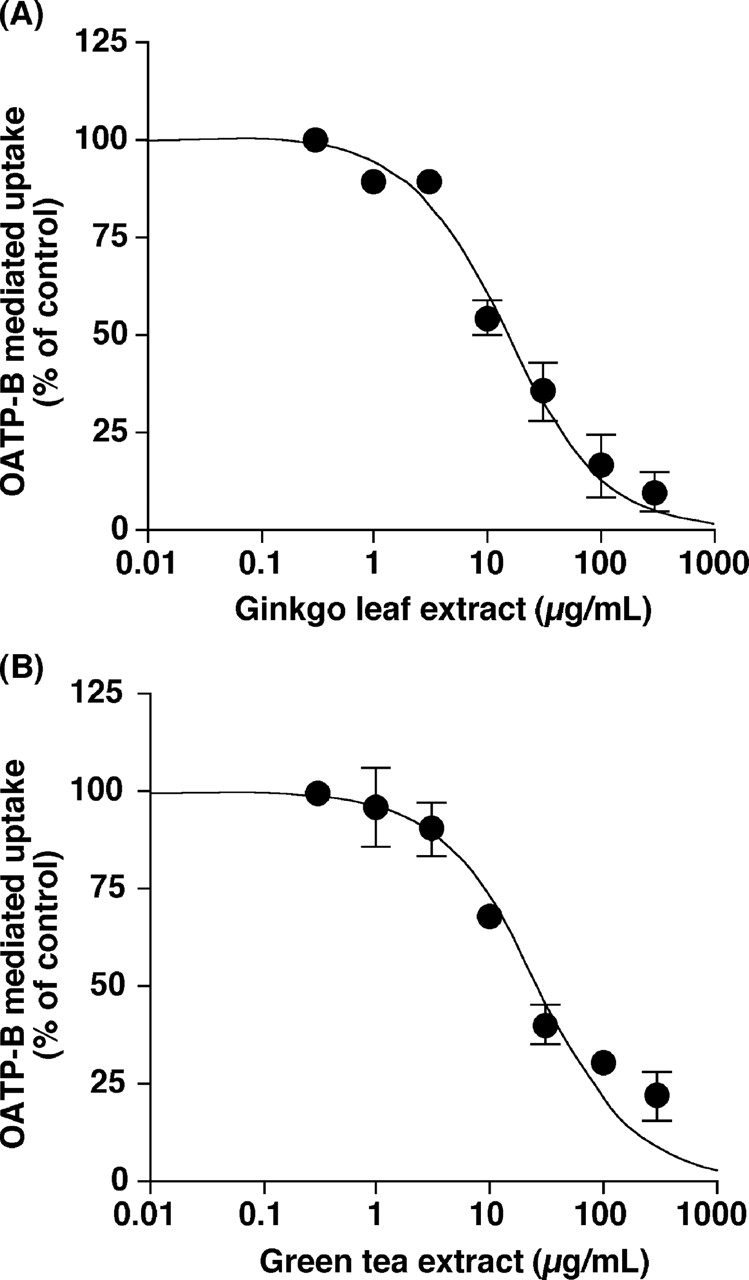
Inhibitory Effects of Ginkgo Leaf and Green Tea Extracts on the Function of OATP-B. Ginkgo leaf and green tea extracts inhibited OATP-B-mediated uptake of estrone-3-sulfate in a concentration-dependent manner (Fig. 3) with the IC50 values of 11.2 ± 3.3 and 22.1 ± 4.9 μg/ml, respectively. Moreover, ginkgo leaf and green tea extracts inhibited OATP-B-mediated uptake of glibenclamide in a concentration-dependent manner (Fig. 4) with the IC50 values of 15.4 ± 2.7 and 24.6 ± 6.1 μg/ml, respectively. When the extracts were washed out, the OATP-B-mediated uptake of estrone-3-sulfate resumed (Fig. 5), suggesting that the inhibition of OATP-B transport by ginkgo leaf extract is reversible.

Inhibitory effects of herbal extracts on the OATP-B-mediated uptake of estrone-3-sulfate. The uptakes of 10 nM [3H]estrone-3-sulfate into HEK/OATP-B cells and HEK/Mock cells were measured for 20 s in the absence (control or 0.5% DMSO) or presence of herbal extracts at 37°C and pH 7.4. The OATP-B-mediated uptake was obtained by subtracting the uptake into HEK/Mock cells from that into HEK/OATP-B cells. The value in parentheses is the tested concentration of each herbal extract (micrograms per milliliter). Each column represents the mean ± S.E.M. of three experiments. The significance of differences from the control (20.4 μl/mg of protein/20 s) or 0.5% DMSO (22.3 μl/mg of protein/20 s) was determined by ANOVA followed by Dunnett's test (*, P < 0.05; **, P < 0.01).

Concentration dependence of the inhibitory effects of ginkgo leaf (A) and green tea (B) extracts on the OATP-B-mediated uptake of estrone-3-sulfate. The uptakes of 10 nM [3H]estrone-3-sulfate into HEK/OATP-B cells and HEK/Mock cells were measured for 20 s in the absence (control) or presence of varying concentrations extracts at 37°C and pH 7.4. The OATP-B-mediated uptake was obtained by subtracting the uptake into HEK/Mock cells from that into HEK/OATP-B cells. Each point represents the mean ± S.E.M. of three experiments.
Concentration dependence of the inhibitory effects of ginkgo leaf (A) and green tea (B) extracts on the OATP-B-mediated uptake of glibenclamide. The uptakes of 10 nM [3H]glibenclamide into HEK/OATP-B cells and HEK/Mock cells were measured for 1 min in the absence (0.5% DMSO) or presence of varying concentrations of the extract at 37°C and pH 7.4. The OATP-B-mediated uptake was obtained by subtracting the uptake into HEK/Mock cells from that into HEK/OATP-B cells. Each point represents the mean ± S.E.M. of three experiments.
Effects of the Major Constituents of Ginkgo Leaf and Green Tea Extracts on the OATP-B-Mediated Uptake of Estrone-3-Sulfate. Terpenoids (ginkgolide A, ginkgolide B, ginkgolide C, and bilobalide), which are contained in ginkgo leaf extract, hardly inhibited the OATP-B-mediated uptake of estrone-3-sulfate (Fig. 6A). On the other hand, kaempferol-3-glucoside and isorhamnetin-3-glucoside at a concentration of 10 μM significantly inhibited estrone-3-sulfate uptake by 54.9 and 47.5%, respectively. Kaempferol-3-rutinoside, quercetin-3-rhamnoside, and quercetin-3-rutinoside at a concentration of 100 μM significantly inhibited the uptake by 45.6, 73.7, and 34.6%, respectively (Fig. 6B). Moreover, EC, ECG, and EGCG, which are contained in green tea extract, at a concentration of 10 μM significantly inhibited estrone-3-sulfate uptake by 27.2, 66.6, and 29.5%, respectively. (+)-Catechin and EGC at a concentration of 100 μM significantly inhibited the uptake by 31.7 and 29.1%, respectively (Fig. 7).
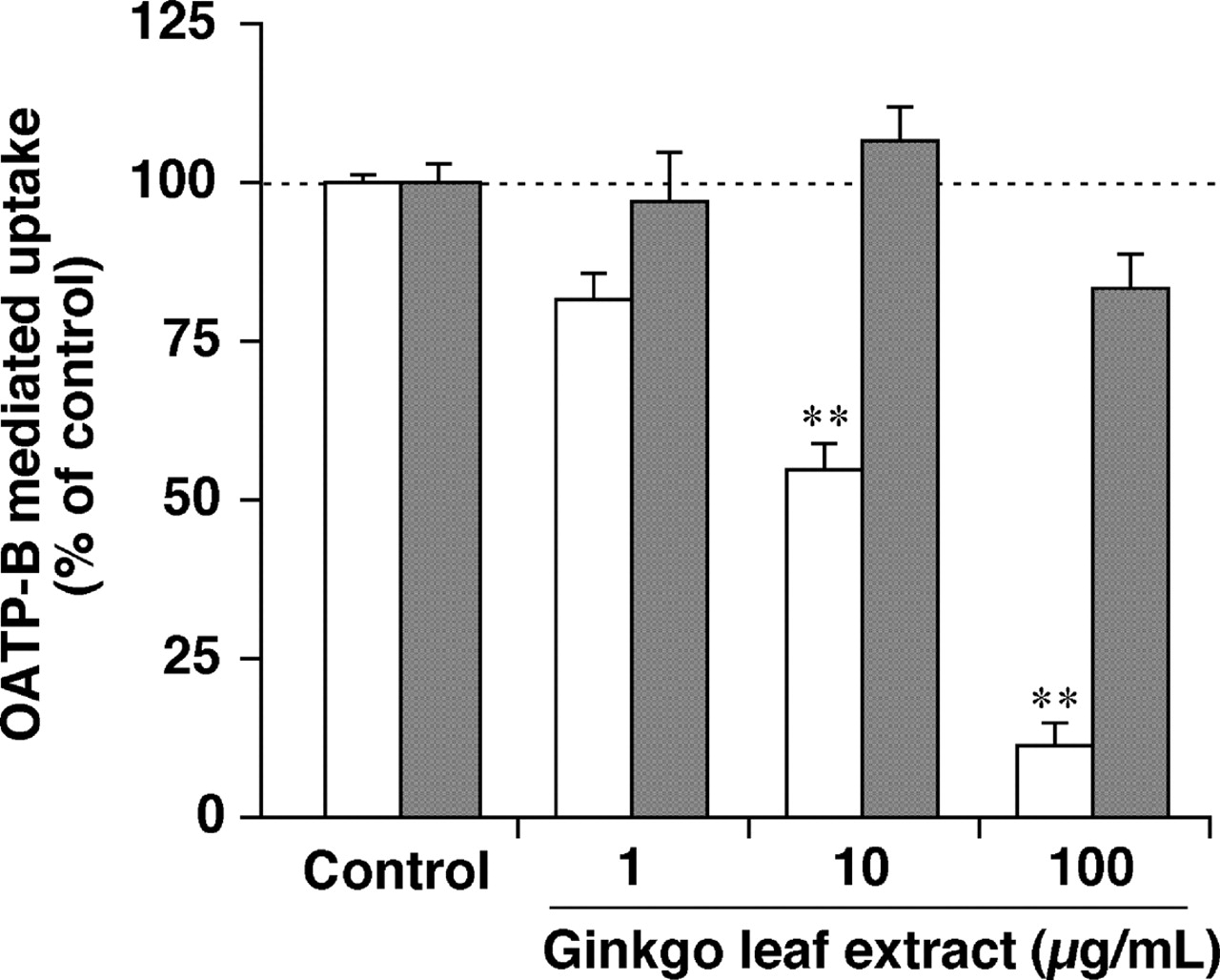
Reversible inhibition of the OATP-B-mediated uptake of estrone-3-sulfate by ginkgo leaf extract. To determine whether the inhibitory effects of ginkgo leaf extract is reversible, the uptakes of 10 nM [3H]estrone-3-sulfate into HEK/OATP-B cells and HEK/Mock cells were measured under the standard conditions (open columns) and the washout conditions, which is washing the cells to remove ginkgo leaf extracts (closed columns). Cells were preincubated with 0.5% DMSO (control) or 1, 10, and 100 μg/ml ginkgo leaf extracts for 10 min, washed five times with uptake buffer, and the uptakes of 10 nM [3H]estrone-3-sulfate into HEK/OATP-B cells and HEK/Mock cells were measured for 20 s at 37°C and pH 7.4. The OATP-B-mediated uptake was obtained by subtracting the uptake into HEK/Mock cells from that into HEK/OATP-B cells. Each column represents the mean ± S.E.M. of three experiments. The significance of differences from the control (21.4 μl/mg of protein/20 s) was determined by ANOVA followed by Dunnett's test (**, P < 0.01).

Inhibitory effects of the major constituents [terpenoids (A) or flavonol glycosides (B)] of ginkgo leaf extract on the OATP-B-mediated uptake of estrone-3-sulfate. The uptakes of 10 nM [3H]estrone-3-sulfate into HEK/OATP-B cells and HEK/Mock cells were measured for 20 s in the absence (control) or presence of 1 μM (open columns), 10 μM (gray columns), or 100 μM (closed columns) constituents at 37°C and pH 7.4. The OATP-B-mediated uptake was obtained by subtracting the uptake into HEK/Mock cells from that into HEK/OATP-B cells. Each column represents the mean ± S.E.M. of three experiments. The significance of differences from the control (24.4 μl/mg of protein/20 s) was determined by ANOVA followed by Dunnett's test (*, P < 0.05; **, P < 0.01).
Discussion
The extracts of barley, Job's tears, rutin, rafuma, and passionflower did not inhibit estrone-3-sulfate uptake by HEK293 cells stably expressing OATP-B at concentrations considered likely to be attainable in the human intestine, whereas the extracts of mulberry, black cohosh, and Siberian ginseng relatively weakly inhibited OATP-B (39.1–49.2%). Therefore, the inhibitory effects of these herbal extracts on the function of OATP-B are unlikely to be clinically important (Fig. 2). On the other hand, the extracts of bilberry, echinacea, green tea, banaba, grape seed, ginkgo, and soybean potently inhibited the OATP-B-mediated uptake of estrone-3-sulfate (55.5–85.4%) at the putative gastrointestinal concentrations, suggesting that routine ingestion of these herbal extracts, especially ginkgo, may significantly affect the function of OATP-B expressed in the intestine and may limit the absorption of drugs transported via OATP-B in humans.
No herbal extracts affected the uptake of estrone-3-sulfate into HEK/Mock cells and showed cytotoxicity at any concentration used in this study (data not shown). Therefore, the permeability of cell membrane was not likely to be affected by these extracts. In addition, ginkgo leaf and green tea extracts inhibited OATP-B-mediated uptake of estrone-3-sulfate in a concentration dependent manner (Fig. 3). Thus, the decreased estrone-3-sulfate uptake may reflect the inhibition of the OATP-B function by extracts. Inhibitory effects (IC50 values) of ginkgo leaf and green tea extracts on glibenclamide uptake were comparable with those on estrone-3-sulfate uptake (Figs. 3 and 4), suggesting that the inhibitory effects of the extracts might not be dependent upon the substrates.

Inhibitory effects of the major constituents of green tea extract on the OATP-B-mediated uptake of estrone-3-sulfate. The uptakes of 10 nM [3H]estrone-3-sulfate into HEK/OATP-B cells and HEK/Mock cells were measured for 20 s in the absence (control) or presence of 1 μM (open columns), 10 μM (gray columns), or 100 μM (closed columns) constituents at 37°C and pH 7.4. The OATP-B-mediated uptake was obtained by subtracting the uptake into HEK/Mock cells from that into HEK/OATP-B cells. Each column represents the mean ± S.E.M. of three experiments. The significance of differences from the control (28.7 μl/mg of protein/20 s) was determined by ANOVA followed by Dunnett's test (**, P < 0.01).
Both flavonol glycosides contained in ginkgo leaf extract and catechins contained in green tea extract significantly inhibited the OATP-B-mediated uptake of estrone-3-sulfate (Figs. 6 and 7). Ginkgo leaf extract contains 25% flavonol glycosides (Kressmann et al., 2002), and green tea extract contains 77% catechins and 13% flavonol (Del Rio et al., 2004). In addition, proanthocyanidins, which account for 90% of grape seed extract, are also catechin derivatives (Singh et al., 2004). Bilberry extract contains 25% anthocyanidin glycosides, which are flavonoids (Ichiyanagi et al., 2004). Soybean extract, which showed a moderate inhibitory effect, contains high levels of isoflavone glycosides (Klejdus et al., 2005). Moreover, we have reported that flavonoids, such as naringin, its aglycone (naringenin), and quercetin, inhibit the function of OATP-B (Satoh et al., 2005). Taken together, the inhibitory effects of the tested herbal extracts on OATP-B may be primarily attributable to flavonoid components.
Although grape seed extract contains a higher amount of catechins than green tea extract, the IC50 value for grape seed extract (22.4 μg/ml; data not shown) was comparable with green tea extract (22.1 μg/ml). It may be due to the differences in the class, number, and position of substituents and glycosylation of flavonoid among the extracts. Indeed, ECG and EGCG (having gallate moiety) exhibited more potent inhibition than (+)-catechin, EC, and EGC (not having gallate moiety).
Recently, Wang et al. (2005) reported that flavonoids such as naringenin and EGCG inhibited the function of OATP-C (OATP1B1, gene symbol SLCO1B1/SLC21A6), a member of the OATP family, which is consistent with our present study. With respect to some compounds, however, their inhibitory characteristics on OATP-C differ from those on OATP-B in our present study. For example, they have reported that quercetin-3-rutinoside (rutin) increased the OATP-C-mediated uptake of DHEAS, a substrate of OATP-C, whereas OATP-B-mediated estrone-3-sulfate uptake was decreased by quercetin-3-rutinoside in the present study. The inhibitory profiles of the inhibitors on OATP-B are not likely to agree with those on OATP-C.
OATP family members have broad substrate specificity and transport bile salts, sulfate or glucuronate derivatives, anionic peptides, and even some cationic compounds (Tirona and Kim, 2002; Hagenbuch and Meier, 2003). However, only estrone-3-sulfate, sulfobromophthalein, and DHEAS have been identified as typical substrates for OATP-B, suggesting that it has high substrate specificity (Kullak-Ublick et al., 2001). It is of interest to know whether OATP-B transports therapeutic drugs. We have previously reported that OATP-B transports a hypoglycemic drug, glibenclamide (Satoh et al., 2005). Other investigators have reported that OATP-B also transports a lipid-lowering drug, pravastatin, and an antihistamine, fexofenadine (Kobayashi et al., 2003; Nozawa et al., 2004).
It has been reported that grapefruit juice and other citrus juices reduce the bioavailability of orally administered fexofenadine, and OATP-B expressed in the intestine is considered to be involved in this juice-drug interaction (Banfield et al., 2002; Dresser et al., 2002). With regard to this interaction, we have reported that 5% GFJ inhibits the function of OATP-B by 82% in vitro (Satoh et al., 2005). Assuming that the volume of gastrointestinal fluid is 1 liter, the concentration of GFJ in the intestinal lumen would be 20 to 33% after taking a glass of juice (250–500 ml). Under this condition, OATP-B is considered to be inhibited by more than 80%. On the other hand, the present study suggested that usual doses of some herbal extracts significantly inhibited OATP-B-mediated transport by 55.5 to 85.4% at the putative gastrointestinal concentration. Thus, the inhibitory effects of these herbal extracts on the function of OATP-B may be comparable with, or a little weaker than, that of GFJ so that it is conceivable that the ingestion of readily available preparations containing these extracts might lead to a decrease in the bioavailability of OATP-B substrate drugs, such as fexofenadine. Recently, it was shown that not only OATP-B but also OATP-A (OATP1A2, gene symbol SLC21A3/SLCO1A2) is expressed on intestinal epithelial cells (Smith et al., 2003). Moreover, Dresser et al. (2005) have shown that the affinity of fexofenadine for OATP-A is higher than that for OATP-B, and they suggested that OATP-A is primarily involved in the decrease in the bioavailability of fexofenadine caused by GFJ. Although both the expression level and affinity of each transporter should be taken into account to estimate the quantitative contributions to drug absorption, OATP-A may be, in part, involved in the absorption of anionic drugs in the intestine as well as OATP-B. Therefore, further studies should be carried out to clarify the expression levels of the OATP isoforms, the affinities of OATP-A and OATP-B for various drugs, and the influence of herbal ingredients on the function of OATP-A.
In conclusion, the extracts of bilberry, echinacea, green tea, banaba, grape seed, ginkgo leaf, and soybean potently inhibited the function of OATP-B at concentrations that seem to be physiologically relevant, suggesting that these extracts may decrease the plasma concentrations of concomitantly ingested OATP-B substrate drugs and may lead to failure of drug therapy.
Acknowledgments
We thank Tokiwa Phytochemical Co., Ltd. for the gifts of herbal extracts.
Footnotes
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.105.007872.
-
ABBREVIATIONS: P450, cytochrome P450; P-gp, P-glycoprotein; OATP, organic anion-transporting polypeptide; EC, (–)-epicatechin; EGC, (–)-epigallocatechin; ECG, (–)-epicatechin gallate; EGCG, (–)-epigallocatechin gallate; DHEAS, dehydroepiandrosterone sulfate; GFJ, grapefruit juice; HEK, human embryonic kidney; DMSO, dimethyl sulfoxide; ANOVA, analysis of variance.
- Received October 19, 2005.
- Accepted January 9, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}