


Abstract
During fetal development, the liver serves as the primary hematopoietic organ in which hematopoietic stem cells (HSC) capable of initiating long-term hematopoiesis comprise a large proportion of the hepatic cell population. Although HSC are potential targets for transplacental chemicals, little is known regarding their xenobiotic biotransformation ability. We quantitated the steady-state mRNA expression of six cytochrome P450 (P450) and 11 glutathione S-transferase (GST) isoforms in CD34+-selected HSC isolated from second trimester human fetal liver donors, genotyped donors for polymorphic hGSTM1 and hGSTT1 status, and analyzed gene expression in HSC relative to total liver from donors of similar gestational ages. Several P450 isoforms, including CYP1A1, CYP2E1, CYP3A4, and CYP3A5, were expressed at low levels in HSC (relative mRNA expression CYP3A5 > CYP1A1 > CYP2E1 > CYP3A4). CYP1A2 and CYP3A7 were not detected in HSC. The CYP3A4/5 mRNA expression in HSC was accompanied by detectable CYP3A protein and low midazolam oxidation activity. Several GST isoforms, including hGSTM1, hGSTM2, hGSTM4, and hGSTP1, were significantly higher in HSC as compared with total fetal liver. With the exception of hGSTA4, alpha class GST were not detected in HSC. GST expression in HSC was accompanied by substantial GST catalytic activity toward 1-chloro-2,4-dinitrobenzene. In summary, our data indicate that fetal liver CD34+-derived HSC constitutively express several P450 isoforms at low levels relative to total hepatic cell populations but have a higher capacity for GST conjugation reactions through mu and pi class isoforms. The functional ramifications of these observations are discussed relative to the sensitivity of human fetal HSC to transplacental chemical injury.
A number of environmental chemicals and drugs have the ability to cross the placental barrier during pregnancy and interact with fetal cell targets (Shu, 1997; Alexander et al., 2001; Woodruff et al., 2004). Because the fetal liver receives a substantial proportion of maternal blood flow during pregnancy, it can be an important target for the exposure to agents that elicit cellular toxicity (Hakkola et al., 1998). However, unlike in adults, the developing fetal liver is not dominated by the presence of hepatocytes but instead by the presence of hematopoietic cells and precursors, and thus serves as the major site of multilineage hematopoiesis (Morrison et al., 1995). Accordingly, a large proportion of the hepatic fetal cell population includes hematopoietic progenitors and hematopoietic stem cells (HSC), which express primitive markers such as CD34+, CD38+, and CD33+, and that can be cell targets for maternally transferred compounds (Sancewicz-Pach et al., 1997; Moneypenny and Gallagher, 2005). It has been proposed that fetal liver HSC injury during pregnancy may underlie the development of certain hematopoietic disorders that develop after birth (Tavassoli, 1991; Shu, 1997; Alexander et al., 2001; Woodruff et al., 2004).
Our laboratory has shown that CD34+ HSC cultured from human fetal liver are extremely sensitive to the toxicity of certain transplacental chemicals or their metabolites, including certain pesticides and anticancer drugs (Moneypenny et al., 2006), and products of oxidative stress (Moneypenny and Gallagher, 2005). In the case of 4-hydroxynonenal, a reactive α,β-unsaturated aldehyde produced in vivo and in utero, the sensitivity of fetal liver HSC greatly exceeds that of other cell types (Moneypenny and Gallagher, 2005). Furthermore, we have recently shown that exposure to low levels of etoposide, a potent DNA topoisomerase II inhibitor, can cause DNA damage and genetic rearrangements in the mixed lineage leukemia (MLL) gene in cultured human fetal liver HSC (Moneypenny et al., 2006). This observation is of particular significance because MLL rearrangements are signature lesions in the infant acute leukemias that have been associated with stem cell injury during pregnancy (Armstrong et al., 2002). Collectively, these observations suggest that human fetal HSC may be relevant targets for transplacental chemicals or their metabolites during pregnancy.
Although the mechanistic basis underlying the susceptibility of HSC to chemical toxicity has not been established, given their primitive nature, it is possible that HSC may have a poor ability to detoxify environmental chemicals and drugs via enzymatic biotransformation pathways. In particular, the selective or polymorphic expression of certain cytochrome P450 (P450) and glutathione S-transferase (GST) isoforms has been associated with susceptibility to a number of drugs and chemicals. For example, the alpha class GST isoform mGSTA3-3 in mice is largely responsible for conferring resistance to the carcinogenic intermediate aflatoxin B1 8-9 epoxide (Buetler et al., 1992; Eaton and Gallagher, 1994). In addition, elevated fetal CYP2E1 expression has been associated with susceptibility to alcohol teratogenesis (Brzezinski et al., 1999), whereas high CYP1A1 protein expression in lung is associated with increased formation of bulky DNA adducts from tobacco smoke carcinogens and susceptibility for smoking-related lung cancer (Mollerup et al., 2006). Although we know that the selective expression of certain xenobiotic metabolizing isoforms can affect in utero organotoxicity (Hakkola et al., 1998), there is little information regarding the ability of sensitive CD34+ HSC from human fetal liver to biotransform xenobiotics. The lack of this information represents a data gap in the area of human developmental toxicology and, in particular, the origins of hematopoietic disorders that may have an etiology based in part on exposures during pregnancy.
In the current study, we have characterized the expression of the predominant P450 and GST genes in HSC prepared from a panel of second trimester human fetal livers. Functionality of mRNA expression was confirmed for several gene subfamilies by catalytic activity assays and/or Western blot analysis. In addition, comparisons of gene expression in CD34+ HSC were made to gene expression in the overall fetal liver containing hematopoietic cells and their progenitors and hepatocytes to better understand the cell origins of P450 and GST gene expression during human development, as well as the ability for sensitive stem cells to carry out P450 and GST-mediated biotransformation.
Materials and Methods
Chemicals and Biochemicals. Iscove's modified Dulbecco's medium, penicillin, streptomycin, and heat-inactivated fetal bovine serum were purchased from Invitrogen (Carlsbad, CA). Recombinant human interleukin 3, recombinant human granulocyte colony stimulating factor, and recombinant human stem cell factor were obtained from Research Diagnostics Inc. (Flanders, NJ). CD34+ magnetic bead isolation columns were purchased from Miltenyl Biotec (Auburn, CA). Western blotting luminol reagent and horseradish peroxidase-linked goat anti-rabbit IgG antibody were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). TRIzol reagent, superscript first-strand synthesis kit, formamide, Denhardt's solution, CotI DNA, polyA(72) primer high-performance liquid chromatography grade water, TaqMan polymerase, Taq antibody, sequence-specific quantitative reverse transcriptase (RT)-polymerase chain reaction (PCR) (Q-PCR) primers and probes, and other molecular biology reagents were purchased from Invitrogen. All the other chemicals were obtained from Sigma Chemical Co. (St. Louis, MO) or Fisher Scientific (Pittsburgh, PA). Vented culture flasks and 96-well plates were purchased from Corning Inc. (Corning, NY). Trypan blue was purchased from Sigma Chemical Co.
Human Liver Tissues and Isolation of CD34+HSC. All use of human tissues was approved by the University of Washington Institutional Research Board. Briefly, the human fetal liver tissues (10–20 weeks gestational age) were provided by the University of Washington Birth Defects Research Laboratory and were obtained by elective termination of pregnancy. Some tissues were rinsed in phosphate-buffered saline (PBS) and frozen at –80° before total RNA and DNA isolation as described below. Other liver specimens used for CD34+ cell isolations were dissociated under sterile conditions, and the resultant total cell population was washed several times in PBS buffer supplemented with 0.3% bovine serum albumin, 2.5 μg/ml amphotericin B/Fungizone, and 50 μg/ml gentamicin sulfate. Human fetal liver-derived CD34+ HSC were isolated using CD34+ magnetic bead-based separation (Miltenyi Biotec). The mononuclear layer was collected by centrifuging over 1.077 g/ml Nycoprep (Invitrogen, Carlsbad, CA) at 400g for 30 min. The CD34+ cells were enriched by magnetic bead separation using the magnetic cell sorting CD34+ isolation (de Wynter et al., 1998), and the purity of the CD34+ HSC averaged >98% as verified by flow cytometry.
After isolation, the CD34+ HSC were seeded at approximately 6250 cells/ml of Iscove's modified Dulbecco's medium containing 15% heat-inactivated fetal bovine serum, 2 ng/ml interleukin 3, 1 ng/ml granulocyte colony stimulating factor, 20 ng/ml stem cell factor, 100 U/ml penicillin, and 100 μg/ml streptomycin (Moneypenny and Gallagher, 2005). The cells were maintained in culture for 7 days, a timeframe which allows for moderate increases in cell number while maintaining a relatively primitive undifferentiated status in a humid chamber at 37°C in 95% O2/5% CO2. HSC under these conditions undergo moderate proliferation and differentiate as a mixed population of several hematopoietic cell lineages. The typical percentage of CD34+, CD33+, and CD38+ on day 7 is approximately 17, 19, and 68% of total cell populations, respectively, and these populations remain dynamic over the culture period (Moneypenny and Gallagher, 2005). Cell counts and viability were determined over the culture period using a hemocytometer and trypan blue exclusion, respectively.
Isolation of DNA, RNA, and Subcellular Fractions. Total genomic DNA from cultured HSC and second trimester fetal livers was extracted using a new Qiagen (Valencia, CA) DNA extraction kit. For the RNA extractions, approximately 4 × 106 HSC or 200 mg of snap-frozen fetal liver tissue was used for isolation of total RNA using the TRIzol reagent according to the manufacturer's instructions. The aqueous layer containing RNA was collected with chloroform, mixed with 70% ethanol, and transferred to a Qiagen RNA isolation column. The integrity of the eluted total RNA was verified using an Agilent (Palo Alto, CA) 2100 Bioanalyzer before we performed Q-PCR. For preparation of subcellular fractions, the cultured HSC were collected by centrifugation, washed twice in PBS, and resuspended in 300 μl of prechilled homogenization buffer containing 250 mM sucrose, 10 mM Tris, 1 mM EDTA, 0.2 mM DTT, and 0.1 mM phenylmethylsulfonyl fluoride, pH 7.4 (added immediately before homogenization). The cells were sonicated for 3- to 5-s bursts, and the homogenates were centrifuged for 15 min at 16,000g at 4°C. The resulting supernatants (S9 fractions containing cytosol and microsomes) were collected and stored at –80°C. The protein concentrations were measured by Bradford Assay using a commercial kit (Sigma Inc.), with bovine serum albumin used as a standard.
Evaluation ofhGSTM1andhGSTT1Gene Deletions. Before Q-PCR analysis, HSC and second trimester fetal liver samples were genotyped for hGSTM1 and hGSTT1 polymorphic expression/deletion using a multiplexed PCR assay (Sweeney et al., 2000). Briefly, two sets of primers were used to amplify a 215-base pair (bp) segment of the hGSTM1 gene and a 480-bp segment of the hGSTT1 gene. Primers were also included to amplify a 268-bp segment of the β-globin gene, which was used as a positive control for the PCR. The products were separated by electrophoresis with ethidium bromidestained 2.5% agarose gel (ISC BioExpress, Kaysville, UT) and genotyped by visual inspection.
Q-PCR Analysis of P450 and GST mRNA Expression. Reverse transcription of 1 to 2 μg of total RNA using oligo(dT)15 primer and Superscript II RNaseH– (Invitrogen) was performed according to the manufacturer's instructions. PCR primers and the dual-labeled probes for the specific genes were designed using the primer design software Primer Express (Applied Biosystems, Foster City, CA) and verified for specificity using BLAST software. For CYP2E1 mRNA expression, the gene-specific primers and probe were designed and verified using software by Roche Applied Science (Indianapolis, IN). The PCR primers and/or the corresponding dual-labeled probes were designed to span intron/exon boundaries and are listed in Tables 1 and 2. The probes were labeled at the 5′ end with 6-carboxy-fluorescein and at the 3′ end with the quenching molecule 6-carboxy-tetramethyl-rhodamine. The PCR mixtures (20 μl final volume) consisted of the appropriate forward and reverse primers (0.35 μM each), 150 nM TaqMan probe, and 1× FAST Universal PCR Master Mix. Gene amplification and detection were measured using the Applied Biosystems 7900HT FAST Real-Time PCR System with the following PCR reaction profile: 1 cycle of 95°C for 15 s, 40 cycles of 95°C for 1 s, and 62°C for 20 s. The relative quantity of target cDNA in each sample was calculated from the experimentally determined CT value using a standard curve.
Primer pairs and probes used to measure human P450 genes by Q-PCR
Primer pairs and probes used to measure human GST genes by Q-PCR
Several commonly used housekeeping genes, including glyceraldehyde-3-phosphate dehydrogenase, β-actin, and 18S RNA, were evaluated for mRNA normalization. A comparison of the steady-state mRNA expression of β-actin and glyceraldehyde-3-phosphate dehydrogenase revealed that these genes are expressed at markedly higher concentrations in the rapidly dividing HSC relative to total fetal liver; therefore, they were not suitable for normalization. The levels of ribosomal 18S RNA expression did not differ among the two groups; therefore, it was used for subsequent normalization of all the P450 and GST genes. The measured relative expression levels for the target P450 and GST genes were divided by the sample's 18S RNA level and multiplied by 1000 to obtain the normalized mRNA expression values presented in the figures.
P450 and GST Catalytic Assays. Several P450 and GST catalytic activities were assayed as a means to determine functionality of P450 and GST proteins encoded by the measured mRNA. Specifically, CYP1A1-dependent ethoxyresorufin O-deethylase (EROD) activity in HSC S9 fractions was measured using a fluorescent microplate reader (Hahn et al., 1993). To determine the functionality of CYP3A5 and CYP3A7 mRNA expression, human fetal HSC were incubated with 12 μM midazolam for 6 h in a shaking water bath at 37°C. Each incubation tube contained approximately 20 × 106 cells in culture media. Reactions were terminated with the addition of an equal volume of 0.1 M Na2CO3, pH11. 15N-labeled metabolite internal standards were added to the samples and standards before extraction using ethyl acetate. The organic phase was transferred to a clean tube, evaporated to dryness, reconstituted in acetonitrile, and derivatized using N-methyl-N-(t-butyl-dimethylsilyl) trifluoroacetamide. Samples and standards were analyzed for 1′-hydroxymidazolam and 4-hydroxymidazolam by negative chemical ionization gas chromatography-mass spectroscopy (Paine et al., 1997). GST activity toward 1-chloro-2,4-dinitrobenzene (CDNB) in S9 fractions was spectrophotometrically measured using 5 mM substrate concentrations in a 96-well microplate reader (Gallagher and Gardner, 2002).
Western Blot Analysis. Western blot analysis for the presence of CYP1A- and 3A-reactive proteins in HSC S9 fractions was conducted using human CYP1A2 and CYP3A4 polyclonal antibodies (Oxford Biomedical Research, Oxford, MI) as described (Doi et al., 2002). Forty micrograms of protein from each treatment group was loaded onto a Bio-Rad (Hercules, CA) Criterion 8 to 16% gradient polyacrylamide gel and size-fractionated. The separated proteins were transferred to a polyvinylidene difluoride membrane and probed with anti-CYP1A2 and anti-CYP3A4 polyclonal antibodies, followed by visualization with horseradish peroxidase-linked goat anti-rabbit IgG secondary antibody (1:2000 in 1% dry milk/Tris-buffered saline/Tween 20). Imaging of the reactive proteins was performed using a Bio-Rad Fluor-S imager and densitometric scanning.
Statistical Analysis. Gene expression data are presented as the mean ± S.E.M. of six HSC and six fetal liver samples, unless otherwise indicated in those donors that genotyped negative for hGSTM1 or hGSTT1. Differences in P450 and GST gene expression among the six second trimester liver donors in which tissues were used to isolate CD34+ HSC, as well as for those donors in which RNA was isolated from total liver, were compared using an unpaired Student's t test (Statview 4.5, Abacus Concepts, Berkeley, CA). Differences in gene expression among the two groups were considered significant at p ≤ 0.05 or p ≤ 0.001.
Results
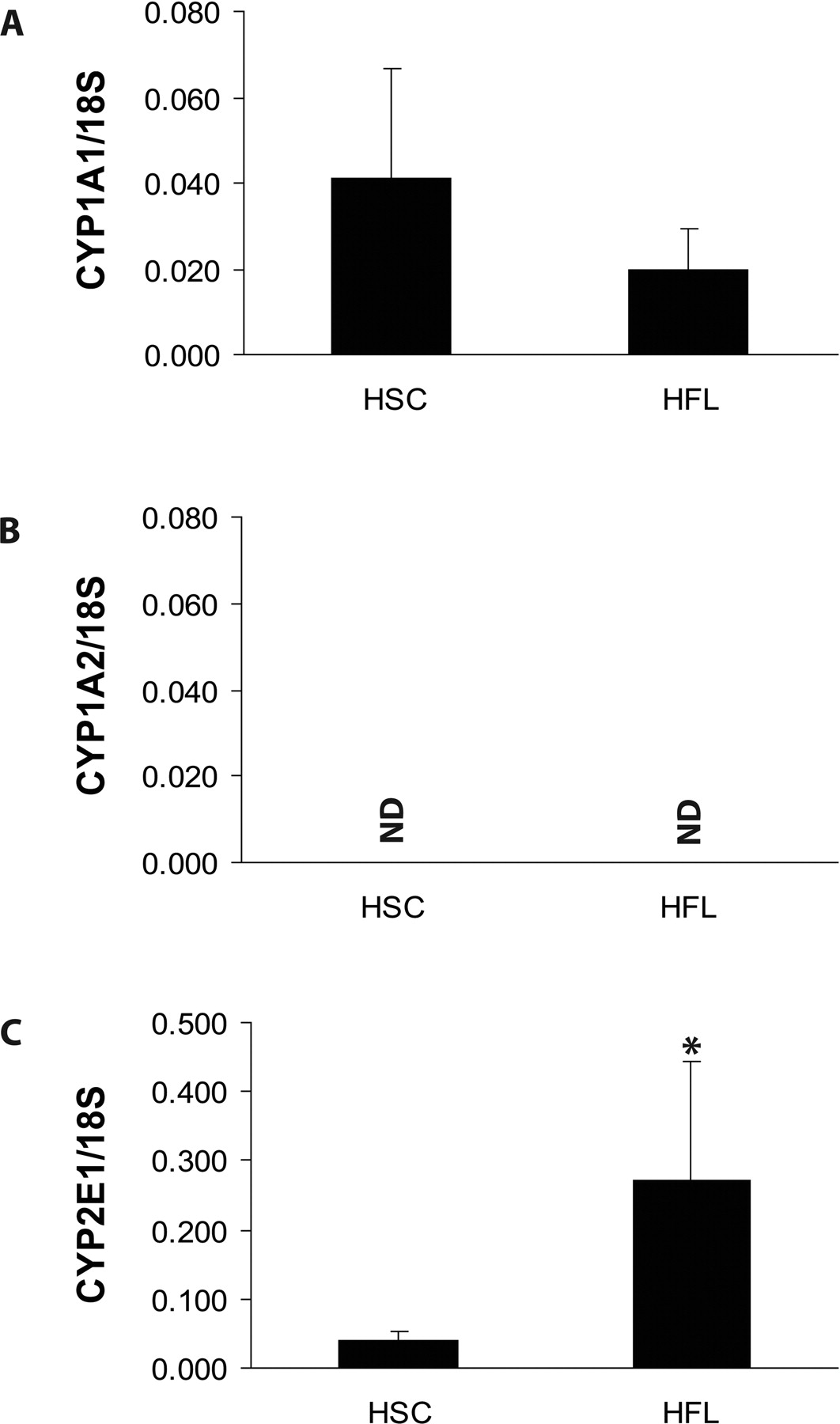
P450 mRNA Expression and Activity in HSC and Fetal Liver. As observed in Figs. 1 and 2, Q-PCR analysis revealed extremely low overall P450 gene expression in the HSC fractions. We were unable to detect CYP1A2 and CYP3A7 transcripts, and transcripts encoding CYP1A1, CYP2E1, CYP3A4, and CYP3A5 mRNA were present at extremely low levels that marginally exceeded the detection limits of Q-PCR. Consistent with other studies, CYP1A1 mRNA was expressed at extremely low levels in total fetal liver (Yang et al., 1995), and also as previously reported, CYP1A2 mRNA was not detected in any of the fetal liver preparations (Yang et al., 1995; Hines and McCarver, 2002). As anticipated, CYP3A7 was the predominant P450 isoform accounting for the majority of P450 content in the fetal liver samples (Fig. 2). Transcripts encoding CYP3A5 were expressed at appreciable levels in the human fetal livers analyzed, whereas transcripts encoding CYP3A4 were barely detectable (Fig. 2). As shown in Fig. 2, CYP3A5 mRNA levels in fetal liver were 70-fold higher than in HSC, whereas CYP3A4 mRNA levels were 20-fold higher in fetal liver than in HSC.
As observed in Fig. 3, the low CYP1A1 mRNA expression in HSC was not accompanied by the presence of detectable CYP1A-reactive proteins (Fig. 3) or EROD activity (Table 3). In contrast, the extremely low CYP3A4 and CYP3A5 expression in HSC was accompanied by low but detectable CYP3A protein cross-reactivity (Fig. 3). Detectable CYP3A-mediated midazolam oxidation was observed in S9 fractions from two HSC donors analyzed, and the rates were 0.094 and 0.111 pmol/h/106 cells (Table 3). A human adult liver microsomal preparation was used as a positive control in the midazolam oxidation studies and yielded a rapid rate of CYP3A-mediated oxidation of 729 pmol/min/mg protein. It should be noted, however, that the midazolam oxidation rates in HSC and adult microsomal samples cannot be directly compared because of unit differences (i.e., cells versus milligram of microsomal protein).
P450 and GST catalytic activities in HSC from two donors

Comparative CYP1A1, CYP1A2, and CYP2E1 gene expression in human HSC and fetal livers. CYP1A1 (A), CYP1A2 (B), and CYP2E1 (C) mRNA expression. Data represent mean ± S.E.M. of six individuals with all the gene expression data normalized to the levels of 18S RNA in each sample. Asterisks denote significant differences in gene expression among HSC and fetal liver samples at p ≤ 0.001.
GST mRNA Expression and Activity in HSC and Fetal Liver. As observed in Fig. 4, HSC did not appreciably express alpha class isoforms hGSTA1, hGSTA2, or hGSTA3, whereas low levels of hGSTA4 were detected by Q-PCR. hGSTA4 mRNA expression was 5-fold higher in total fetal liver compared with HSC. All the alpha class GST isoforms analyzed were readily detected in the fetal livers, with the alpha GST expression levels being hGSTA1 > hGSTA2 > hGSTA4 > hGSTA3. All five mu class hGST were detected in both HSC and fetal liver, and for the most part the mu class hGST were expressed at higher concentrations in HSC (an exception being the relatively low expression of hGSTM5) (Fig. 5). All the HSC and fetal liver samples were genotyped for deletion polymorphism in hGSTM1 gene, and three HSC donors and two fetal liver donors were negative for the presence of hGSTM1 alleles and not used in quantitative comparisons of hGSTM1 mRNA expression among the two groups. For individuals expressing at least one copy of hGSTM1, we did not distinguish the existence of one allele or both alleles. The level of hGSTM1 in HSC was 32-fold higher than in fetal liver. The level of hGSTM2 in HSC was 3-fold higher than in fetal liver, whereas hGSTM3 mRNA transcripts were relatively abundant and similar in HSC and fetal liver (Fig. 5). Interestingly, the expression of hGSTM4 in HSC was the highest among all five mu class hGST analyzed. In contrast, hGSTM5 was the least abundant mu class isoform, and the level of its expression in HSC was 3.3-fold lower than in fetal liver (Fig. 5). Collectively, the level of expression for the five mu class hGST in HSC was hGSTM4 > hGSTM1 ≈ hGSTM3 > hGSTM2 > hGSTM5 (Fig. 5).

Comparative CYP3A4, CYP3A5, and CYP3A7 gene expression in human HSC and fetal livers. CYP3A4 (A), CYP3A5 (B), and CYP3A7 (C) mRNA expression. Data represent mean ± S.E.M. of six individuals with all the gene expression data normalized to the levels of 18S RNA in each sample. Asterisks denote significant differences in gene expression among HSC and fetal liver samples at p ≤ 0.001.
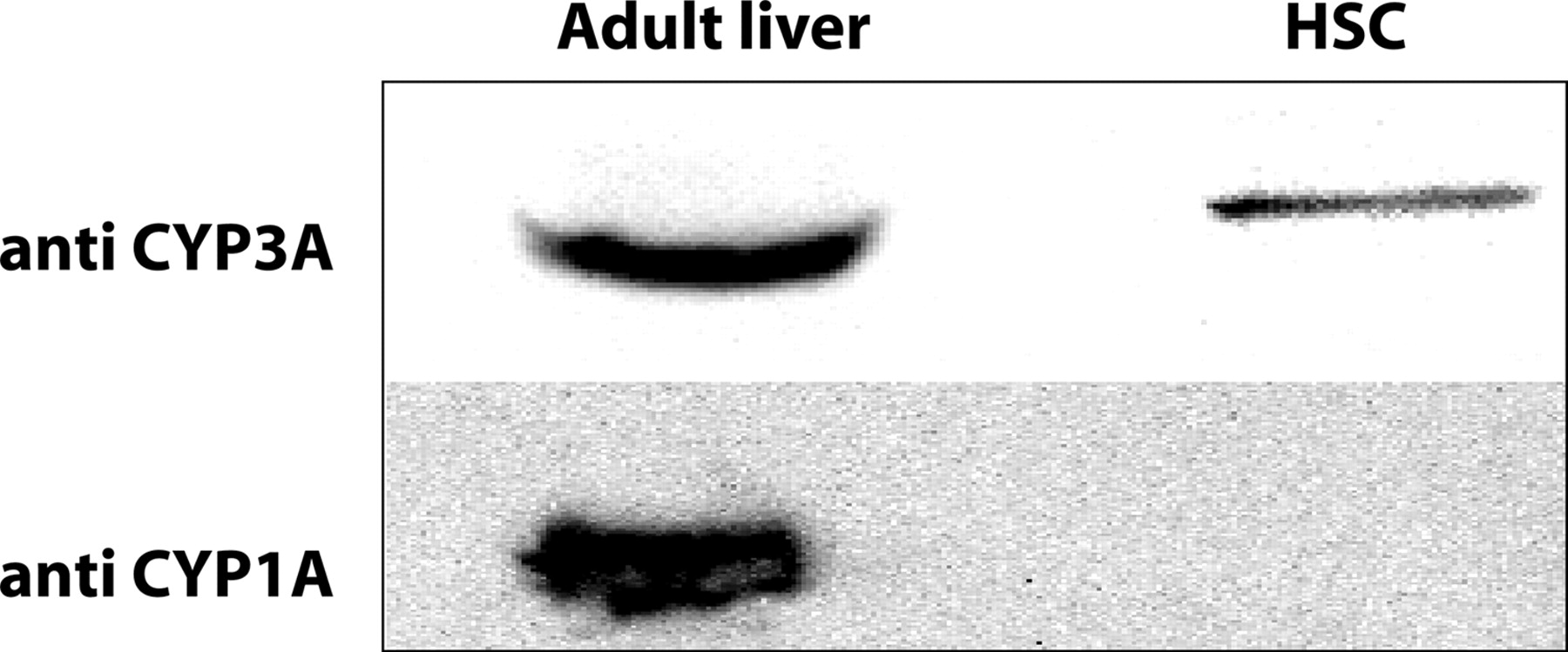
Western blot analysis of CYP1A- and CYP3A-immunoreactive proteins in HSC. HSC S9 (40 μg of each blot) were probed using anti-human CYP1A2 and CYP3A4 antibodies. Liver microsomes from an adult donor (HAL, 5 μg for CYP3A and 20 μg for CYP1A detection) were used as a positive control for the Western blots.
The comparative expression of hGSTP1 and hGSTT1 mRNA in HSC and fetal liver is presented in Fig. 6. hGSTP1 mRNA was expressed at approximately 6-fold higher levels in HSC relative to the fetal liver. Genotyping for the presence/absence of the hGSTT1 gene revealed that at least one hGSTT1 allele was present in all the samples analyzed. As with hGSTM1, we did not distinguish the number of hGSTT1 alleles that were present in those individuals. As observed, comparative expression of hGSTT1 mRNA was similar among HSC and in the total fetal liver. The presence of many of the GST mRNA isoforms in HSC was accompanied by readily detectable GST activity toward the overall GST substrate CDNB in S9 fractions from the two donors analyzed. As shown in Table 3, the GST-CDNB activity for the two donors was 93 and 166 nmol/min/mg protein.
Discussion
There are very few reports in the literature of biotransformation enzyme expression in HSC, with a few studies centering on gene expression and catalytic activities using CD34+ cells from bone marrow or peripheral blood (Czerwinski et al., 1997; Bernauer et al., 2000; Kousalova et al., 2004; Czekaj et al., 2005). For example, CYP2E1 is present in CD34+ stem cells derived from human, rat, and rabbit bone marrow, and human CD34+ cells from blood have functional CYP2E1 activity toward chlorzoxazone (Kousalova et al., 2004). In contrast, CYP3A4 protein or CYP3A-dependent catalytic activities are not present in human CD34+ cells from peripheral blood (Kousalova et al., 2004). However, differences in gene expression, function, and phenotype exist among CD34+ cells isolated from various human tissue sources, thus necessitating caution in extrapolating drug metabolism capabilities of CD34+-derived cells. Although we did not analyze for the expression of all the P450 and GST isoforms, our goal was to characterize the major isoforms in the cells that participate in xenobiotic metabolism.
CYP3A7 predominates in human fetal liver from early gestation to 6 months postnatally (de Wildt et al., 1999; Stevens et al., 2003) and contributes to the regulation of retinoic acid metabolism and estriol levels (Kitada et al., 1987; Marill et al., 2000). The lack of detectable CYP3A7 mRNA in HSC suggests that fetal hepatocytes or other fetal hepatic cell types are the major source of this P450 isoform. CYP3A4 expression is extremely low in human fetal liver but increases postnatally (de Wildt et al., 1999; Stevens et al., 2003). We confirmed that CYP3A4 mRNA is expressed in HSC at lower levels than in the fetal liver, suggesting a poor capacity of CYP3A4-mediated metabolism in HSC. The relatively high expression of CYP3A5 in HSC, whose protein product shares some substrate similarities with CYP3A4 (Gorski et al., 1994; Huang et al., 2004), indicates that CYP3A5 may be more important for drug metabolism in fetal liver CD34+-derived HSC. However, the extremely low CYP3A-mediated midazolam oxidation activity observed in two HSC donors, along with the relatively low CYP3A mRNA and protein expression, suggests a poor capacity for CYP3A-dependent oxidation of transplacental drugs and chemicals in these hematopoietic cells.

Comparative alpha class GST gene expression in human HSC and fetal livers. hGSTA1 (A), hGSTA2 (B), hGSTA3 (C), and hGSTA4 (D) mRNA expression. Data represent mean ± S.E.M. of six individuals with all the gene expression data normalized to the levels of 18S RNA in each sample. Asterisks denote significant differences in gene expression among HSC and fetal liver samples at p ≤ 0.001.

Comparative mu class GST gene expression in human HSC and fetal livers. hGSTM1 (A), hGSTM2 (B), hGSTM3 (C), hGSTM4 (D), and hGSTM5 (E) mRNA expression. Data represent mean ± S.E.M. of six individuals, except for polymorphic hGSTM1 where at least one allele was present in n = 3 HSC donors and n = 4 fetal liver donors. All the gene expression data were normalized to the levels of 18S RNA. Asterisks denote significant differences in gene expression among HSC and fetal liver samples at p ≤ 0.001.

Comparative hGSTP1 and hGSTT1 gene expression in human HSC and fetal livers. hGSTP1 (A) and hGSTT1 (B) mRNA expression. Data represent the mean ± S.E.M. of six individuals, and all the gene expression data were normalized to the levels of 18S RNA in each sample. Asterisks denote significant differences in gene expression among HSC and fetal liver samples at p ≤ 0.05.
CYP1A1 and CYP1A2 proteins are active in the bioactivation of a number of carcinogens and are typically expressed at low levels during development (Yang et al., 1995; Hines and McCarver, 2002). The fact that the low CYP1A1 mRNA levels in fetal HSC were not accompanied by detectable CYP1A-related proteins by Western blotting or CYP1A1-dependent EROD activity indicates that the low level of CYP1A1 mRNA does not result in functional protein in these cells. Alternatively, it is possible that the lack of detectable CYP1A protein in fetal HSC could be explained by alternative CYP1A1 gene splicing during translation that occurs in rat and human brain (Chinta et al., 2005). Similar to CYP1A2, CYP2E1 is expressed at low levels in fetal liver with up-regulation of expression occurring at birth (Mori et al., 2002). CYP2E1 is the high-affinity liver P450 isoform involved in the metabolism and activation of several aliphatic and aromatic halogenated hydrocarbons (Hines and McCarver, 2002; Gonzalez, 2005). CYP2E1-null mice develop normally and show no overt deleterious phenotype (Gonzalez, 2005); thus, the physiological function of this isoform during development is unknown. Although we did not analyze for CYP2E1-dependent catalytic activities in HSC, the low CYP2E1 mRNA expression in fetal liver HSC suggests a poor ability of these cells to metabolize and bioactivate CYP2E1 substrates of toxicological significance. Considering that a host of P450 isoforms involved in the bioactivation of chemical carcinogens (e.g., CYP1A1, CYP1A2, CYP3A7, and CYP2E1) display either low expression or are not detectable in fetal liver HSC, we would hypothesize that these cells have a poor capacity to bioactivate chemical carcinogens through these pathways. We are currently exploring these questions in catalytic studies.
Our laboratory, as well as others, have shown that several alpha, mu, and pi class GST proteins are abundant in human fetal liver tissues (Mera et al., 1994; Gallagher and Gardner, 2002). In the current study, GST mRNA expression profiles in HSC markedly contrasted those observed for the various P450s in that a number of GST were expressed at higher levels than in fetal liver. Several of the human fetal donors analyzed for hGSTM1 polymorphisms exhibited gene deletions, with the percentage of gene deletion consistent with the general Caucasian population (50%) (Dagnino-Subiabre et al., 2000; Hayes et al., 2005). In hGSTM1-null individuals, the absence of functional hGSTM1-1 protein could potentially impact the ability of cells to metabolize polycyclic aromatic hydrocarbon metabolites formed in utero. Of particular note was the relatively higher expression of hGSTM1, hGSTM2, and hGSTM4 mRNA in HSC relative to the total fetal liver. In fact, CD34+ HSC are likely major origins of mu class GST during the second trimester in fetal liver, thus indicating an important function for these proteins in HSC. Little is known regarding the substrate specificity for mu class GST isoforms other than hGSTM1-1. hGSTM2-2 conjugates 1,2-dichloro-4-nitrobenzene, aminochrome, and dopa O-quinone (Dagnino-Subiabre et al., 2000; Hayes et al., 2005) with glutathione. There is less known about the preferred substrates of hGSTM3-3, hGSTM4-4, and hGSTM5-5, although these isoforms can conjugate the general GST substrate CDNB (Patskovsky et al., 2000).
Of note was our observation of high expression of hGSTP1 mRNA in HSC, which exceeded that observed in fetal liver. As discussed, hGSTP1-1 and hGSTA1-1 are the major human fetal liver GST isoforms. Interindividual differences in hGSTP1-1 protein expression have been linked to susceptibility against the mutagenic action of a number of carcinogens and their intermediates (reviewed in Hayes and Pulford, 1995; Eaton and Bammler, 1999). When considered in the context of the absence or relatively low expression of CYP1A1 and other bioactivating P450 isoforms, we would hypothesize that human HSC would not form bulky DNA adducts from certain carcinogens. It was also previously reported that at least one θ-like GST is present at the second trimester in human fetal liver (Mera et al., 1994). Our results show that hGST1 mRNA is expressed at appreciable levels in the total fetal liver and also in the HSC subpopulations. The hGSTT1-1 protein is active in the metabolism of several pesticides and may partially protect fetal liver HSC from transplacental pesticide exposure (Hayes and Pulford, 1995). However, there does not appear to be a single class of chemicals preferentially metabolized by hGST1-1, and this isoform bioactivates certain industrial agents such as methyl bromide and methylene chloride to genotoxic metabolites, which may affect susceptibility to genotoxic injury from these compounds (Hallier et al., 1993; Kempkes et al., 1996).
With the exception of hGSTA4, the alpha class GST were not appreciably expressed in HSC. The low but detectable hGSTA4 expression in HSC is consistent with our earlier report of low expression of hGSTA4 mRNA in 70 human tissues and human cell lines (Gallagher and Gardner, 2002). Of the alpha class GST, hGSTA4-4 is the high affinity isoform that conjugates 4-hydroxynonenal (Hubatsch et al., 1998). The extremely low expression of hGSTA4 mRNA in HSC is consistent with the sensitivity of these cells to 4-hydroxynonenal toxicity (Moneypenny and Gallagher, 2005). We also previously reported relatively high expression of hGSTA1 mRNA and hGSTA1-1 catalytic activities in second trimester human fetal liver (Gallagher and Gardner, 2002). It is of interest that hGSTA1 is not expressed in fetal liver HSC, as this alpha class GST has comparatively high constitutive expression in most human tissues (Gallagher and Gardner, 2002). It has been reported that hGSTA1 mRNA is not expressed in CD34+ cells derived from human bone marrow (Czerwinski et al., 1997). Because the hGSTA1-1, hGSTA2-2, and hGSTA4-4 proteins are the dominant GST isoforms that protect against oxidative damage (Hayes and Pulford, 1995; Hubatsch et al., 1998), lack of these isoforms in HSC suggests that GST may not constitute an important pathway of protection against byproducts of oxidative stress in these cells. hGSTA3-3 is expressed in steroidogenic tissues and catalyzes the glutathione-dependent isomerization of steroids (Johansson and Mannervik, 2001). Thus, hGSTA3-3 may play a role in regulation of steroid hormone biosynthesis. Polymorphisms in the coding region of hGSTA3 may modulate the effectiveness of hGSTA3-3 in steroid hormone synthesis (Johansson and Mannervik, 2001; Tetlow et al., 2004). If the primary role of this GST isoform lies in steroid biosynthesis, we would not expect to find appreciable hGSTA3 mRNA expression in primitive HSC as a result of the hematopoietic functionality of these cells.
In summary, our studies indicate an overall low expression profile in human fetal liver HSC for many P450 isoforms that confer chemical metabolic capabilities in other tissues. In contrast, we observed relatively high expression of certain GST isoforms and functional GST catalytic activity, indicating that HSC have the potential to conjugate reactive intermediates and thus reduce the potential for DNA damage in these sensitive cells. Ongoing studies in our laboratory are directed toward a better understanding of the role of xenobiotic metabolizing and conjugating enzymes in the susceptibility of HSC to chemicals of relevance in utero.
Footnotes
-
This work was supported in part by grants from National Institutes of Health (R01-ES09427, P30-ES07033), the United States Environmental Protection Agency STAR program (R-827441), the National Oceanic and Atmospheric Administration Coastal Ocean Program (NA05NS4781253), the University of Washington Royalty Research Fund, the University of Washington Birth Defects Research (NIH-HD000836), and by the National Institutes of Health Roadmap/National Center for Research Resources (K12-8K12RR023265-02).
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.106.012757.
-
ABBREVIATIONS: HSC, hematopoietic stem cell(s); MLL, mixed lineage leukemia; P450, cytochrome P450; GST, glutathione S-transferase; RT, reverse transcriptase; PCR, polymerase chain reaction; Q-PCR, quantitative reverse transcriptase-polymerase chain reaction; PBS, phosphate-buffered saline; bp, base pair; EROD, ethoxyresorufin O-deethylase; CDNB, 1-chloro-2,4-dinitrobenzene.
- Received September 15, 2006.
- Accepted October 17, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}