


Abstract
[3R,4R,5S]-4-Acetamido-5-amino-3-(1-ethylpropoxy)-1-cyclohexene-1-carboxylate phosphate (Ro 64-0802) is a pharmacologically active form of the anti-influenza virus drug oseltamivir. Abnormal behavior is a suspected adverse effect of oseltamivir on the central nervous system. This study focused on the transport mechanisms of Ro 64-0802 across the blood-brain barrier (BBB). Ro 64-0802 was found to be a substrate of organic anion transporter 3 (OAT3/SLC22A8) and multidrug resistance-associated protein 4 (MRP4/ABCC4). Human embryonic kidney 293 cells expressing OAT3 exhibited a greater intracellular accumulation of Ro 64-0802 than mock-transfected cells (15 versus 1.2 μl/mg protein/10 min, respectively). The efflux of Ro 64-0802 was 3-fold greater when MRP4 was expressed in MDCKII cells and was significantly inhibited by indomethacin. After its microinjection into the cerebrum, the amount of Ro 64-0802 in brain was significantly greater in both Oat3–/– mice and Mrp4–/– mice compared with the corresponding wild-type mice (0.36 versus 0.080 and 0.32 versus 0.060 nmol at 120 min after injection, respectively). The brain/plasma concentration ratio (Kp,brain) of Ro 64-0802, determined in wild-type mice after subcutaneous continuous infusion for 24 h, was close to the capillary volume (approximately 10 μl/g brain). Although the Kp,brain of Ro 64-0802 was unchanged in Oat3–/– mice, it was significantly greater in Mrp4–/– mice (41 μl/g of brain). These results suggest that Ro 64-0802 can cross the BBB from the blood, but its brain distribution is limited by its active efflux by Mrp4 and Oat3 across the BBB. The transporter responsible for the brain uptake of Ro 64-0802 remains unknown, but Oat3 is a candidate transporter.
Oseltamivir is an ester-type prodrug of Ro 64-0802, a potent and selective inhibitor of viral neuraminidase, a key enzyme involved in the release of influenza virus from host cells. Oseltamivir is used for the treatment and prophylaxis of infectious diseases caused by both Influenzavirus A and Influenzavirus B (Bardsley-Elliot and Noble, 1999). In recent studies, abnormal behavior, such as jumping and falling from balconies, has been reported in teenagers or younger people who are taking oseltamivir (http://www.fda.gov/cder/drug/infopage/tamiflu/QA20051117.htm; Fuyuno, 2007). In response to these reports, the Ministry of Health, Labor and Welfare has issued a warning regarding the use of oseltamivir as a medication for teenagers or younger people and has prohibited the prescribing of oseltamivir for them in Japan.
The pharmacological actions of oseltamivir on the central nervous system have been reported in several animal studies (Izumi et al., 2007; Satoh et al., 2007; Usami et al., 2008; Yoshino et al., 2008), although the association between such pharmacological actions and abnormal behavior remains an open question. The systemic administration of oseltamivir increases dopamine levels in the rat medial prefrontal cortex (Yoshino et al., 2008), and oseltamivir and Ro 64-0802 enhance spike synchronization between hippocampal CA3 pyramidal cells and evoked synchronized population bursts, which recruit virtually all of the neurons in the network (Usami et al., 2008). It has also been demonstrated that oseltamivir and Ro 64-0802 affect neuronal excitability in rat hippocampal slices and that Ro 64-0802 is 30 times more potent than oseltamivir (Izumi et al., 2007).
Whether oseltamivir and Ro 64-0802 cross the blood-brain barrier (BBB) is an important issue, considering their pharmacological actions on the central nervous system. In clinical studies, both oseltamivir and Ro 64-0802 were detected in the plasma after oral administration of oseltamivir. Oseltamivir is converted to Ro 64-0802 by carboxylesterase 1A1 (CES1A1) in the liver (Shi et al., 2006). Most of the administered dose is recovered in the urine as Ro 64-0802 by glomerular filtration and tubular secretion by organic anion transporters in the kidney (He et al., 1999). The penetration of drugs to the brain from the circulating blood is limited by the BBB, which is formed by endothelial cells connected tightly to adjacent cells. It has been shown that oseltamivir can cross the BBB, but P-glycoprotein (P-gp) limits its brain penetration at the BBB (Morimoto et al., 2008; Ose et al., 2008). In contrast, Ro 64-0802 exhibits only a limited distribution in the brain because it is close to the brain capillary volume. Therefore, the permeability of Ro 64-0802 across the BBB has been considered to be quite low because of its hydrophilic nature and anionic charge at neutral pH.
In this study, we hypothesized that the low distribution of Ro 64-0802 in the brain is attributable to active efflux at the BBB. We focused on two organic anion transporters—organic anion transporter 3 (OAT3/SLC22A8) and multidrug resistance-associated protein 4 (MRP4/ABCC4)—as the candidate transporters involved. Oat3 is expressed on the abluminal membrane of the brain capillary endothelial cells in rodents (Kikuchi et al., 2003; Mori et al., 2003; Roberts et al., 2008). Cumulative in vivo studies suggest that Oat3 plays a significant role in the uptake of hydrophilic organic anions into the endothelial cells, the first step in its overall elimination from the brain to the blood (Ohtsuki et al., 2002; Kikuchi et al., 2003, 2004; Mori et al., 2003, 2004). Ro 64-0802 has been identified as a substrate of OAT1/SLC22A6 (Hill et al., 2002). Considering the overlapping substrate specificities of OAT1 and OAT3, it is possible that OAT3 accepts Ro 64-0802 as substrate. MRP4 is an ATP-binding cassette transporter localized in the luminal membrane of the brain capillary endothelial cells (Leggas et al., 2004). MRP4 accepts anionic drugs as substrates (Ci et al., 2007; Hasegawa et al., 2007; Imaoka et al., 2007) and mediates their unidirectional efflux into the circulating blood (Leggas et al., 2004). It has been demonstrated that the elimination of topotecan from the brain was delayed and that the concentration of topotecan in the cerebrospinal fluid was greatly enhanced in Mrp4–/– mice compared with the corresponding wild-type mice (Leggas et al., 2004). It has also been demonstrated that the brain/plasma concentration ratio of 9′-(2′-phosphonylmethoxyethyl)-adenine 3 h after intravenous administration was greater in Mrp4–/– mice than in wild-type mice (Belinsky et al., 2007).
In this study, in vivo experiments were undertaken using wild-type, Oat3–/–, and Mrp4–/– mice to examine the involvement of Oat3 and Mrp4 in the uptake and efflux of Ro 64-0802 across the BBB.
Materials and Methods
Reagents. Oseltamivir phosphate and its active metabolite, Ro 64-0802 (purity >95%), were synthesized according to a previous report (Yamatsugu et al., 2007). All other chemicals used in the experiments were of analytical grade.
Animals.Mrp4–/– mice had been established previously (Leggas et al., 2004). Oat3–/– mice were obtained from Deltagen, Inc. (San Carlos, CA). Male C57BL/6J, Mrp4–/–, and Oat3–/– mice were maintained by CLEA Japan, Inc. (Tokyo, Japan). All mice (10–18 weeks old) were maintained under standard conditions with a reverse dark-light cycle. Food and water were available ad libitum. All experiments using animals in this study were performed according to the guidelines provided by the Institutional Animal Care Committee (Graduate School of Pharmaceutical Sciences, University of Tokyo).
Uptake of Ro 64-0802 by Human OAT3-Expressing HEK293 Cells. An in vitro transport experiment was performed as described previously (Deguchi et al., 2004). After the cells had been washed twice and preincubated with Krebs-Henseleit buffer at 37°C for 15 min, drug uptake was initiated by the addition of Krebs-Henseleit buffer containing Ro 64-0802 (10 μM). The Krebs-Henseleit buffer consisted of 118 mM NaCl, 23.8 mM NaHCO3, 4.8 mM KCl, 1.0 mM KH2PO4, 1.2 mM MgSO4, 12.5 mM HEPES, 5.0 mM glucose, and 1.5 mM CaCl2, adjusted to pH 7.4. Uptake was terminated at the designated times by the addition of ice-cold Krebs-Henseleit buffer after the removal of the incubation buffer. The cells were then washed twice with 1 ml of ice-cold Krebs-Henseleit buffer, solubilized in 500 μl of 1 mM Tris-HCl buffer (pH 7.4), and stored overnight at 4°C. After sonication, aliquots (250 μl) were subjected to liquid chromatography (LC)-mass spectrometry (MS) analysis. The remaining 20 μl of cell lysate was used to determine the protein concentration by the method of Lowry et al. (1951), with bovine serum albumin as the standard.
Construction of Human MRP4/CES1A1-Expressing MDCKII Cells. CES1A1 cDNA was subcloned into the pTARGET vector (Promega, Madison, WI) (Mori et al., 1999) and transfected into MDCKII cells with Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA), according to the manufacturer's protocol. The transfectants were selected by culturing them in the presence of neomycin (1600 μg/ml) (Invitrogen) and were maintained in Dulbecco's modified Eagle's medium (Invitrogen) supplemented with 10% fetal bovine serum, 1% antibiotic-antimycotic (Invitrogen), and neomycin (400 μg/ml) at 37°C with 5% CO2 and 95% humidity. MDCKII cells with sufficient CES1A1 activity (CES1A1-MDCKII) were cloned and used as the hosts for infection with recombinant adenovirus carrying the human MRP4 gene, which had been established previously (Ci et al., 2007; Hasegawa et al., 2007; Imaoka et al., 2007). CES1A1-MDCKII cells were infected with recombinant adenovirus containing human MRP4 transporter cDNA at a multiplicity of infection of 10 for 48 h to overexpress human MRP4 (MRP4/CES1A1-MDCKII). Green fluorescent protein (GFP) was used as the negative control (GFP/CES1A1-MDCKII).
The expression of MRP4 protein was confirmed by Western blotting. The cell lysates were loaded onto a SDS-polyacrylamide gel (7.5%) with a 3.75% stacking gel. N-Linked carbohydrate groups were cleaved from the MRP4 protein in the cell lysates with N-glycosidase F (PNGase F; New England Biolabs, Ipswich, MA). Digestion was performed according to the manufacturer's instructions, except that the samples were incubated for 30 min at 37°C in denaturing buffer rather than for the recommended 10 min at 100°C. To minimize protein degradation, protease inhibitors were included in all the steps. After incubation at 37°C for 30 min, the samples were separated by SDS-polyacrylamide gel electrophoresis (7.5%). The proteins were electroblotted on to a polyvinylidene difluoride membrane (Pall Corporation, East Hills, NY). The membrane was blocked with blocking buffer [Tris-buffered saline containing 0.05% Tween 20 (TTBS) and 3% skimmed milk] for 1 h at room temperature. After it had been washed with TTBS, the membrane was incubated overnight at 4°C with monoclonal anti-MRP4 M4I-10 antibody (1:1000 in blocking buffer; Abcam Inc., Cambridge, MA). The protein was detected by binding horseradish peroxidase-labeled anti-rat IgG antibody (1:5000 in blocking buffer; GE Healthcare, Little Chalfont, Buckinghamshire, UK). Immunoreactivity was detected with an ECL Plus Western Blotting Detection Kit (GE Healthcare).
Efflux of Ro 64-0802 Formed Intracellularly from Oseltamivir in MRP4/CES1A1-MDCKII and GFP/CES1A1-MDCKII Cells. After the cells had been washed twice and preincubated with Krebs-Henseleit buffer at 37°C for 15 min, oseltamivir (10 μM) was added to the incubation buffer in the presence or absence of indomethacin (50 μM). At the designated times, the incubation buffer was collected. Ice-cold Krebs-Henseleit buffer was then added, and the cells were washed four times. After the cells had been collected, they were frozen in liquid nitrogen and stored at –80°C until use. The efflux clearance of Ro 64-0802 from the transfectants was determined using the integration plot method. The amount of Ro 64-0802 effluxed to the buffer at time t [Xbuffer (t), nanomoles per milligram of protein] can be described by the following equation:  where CLefflux (microliters per minute per milligram of protein) represents the efflux clearance of Ro 64-0802 from the cells, and Ccell (t) (micromolar concentration) is the cellular concentration of Ro 64-0802. Cellular volume was assumed to be 4 μl/mg protein. The integration of this equation from time 0 to time t yields the following equation:
where CLefflux (microliters per minute per milligram of protein) represents the efflux clearance of Ro 64-0802 from the cells, and Ccell (t) (micromolar concentration) is the cellular concentration of Ro 64-0802. Cellular volume was assumed to be 4 μl/mg protein. The integration of this equation from time 0 to time t yields the following equation:  where AUCcell (0 –t) (micromolar concentration × min) represents the area under the cellular concentration-time curve for Ro 64-0802 from time 0 to time t. Because the amount of Ro 64-0802 in buffer [Abuffer (t), nanomoles per milligram of protein] is given by the sum of Xbuffer (t) and the amount of Ro 64-0802 existing in the buffer at time 0 (X0), Abuffer (t) is described by the following equation:
where AUCcell (0 –t) (micromolar concentration × min) represents the area under the cellular concentration-time curve for Ro 64-0802 from time 0 to time t. Because the amount of Ro 64-0802 in buffer [Abuffer (t), nanomoles per milligram of protein] is given by the sum of Xbuffer (t) and the amount of Ro 64-0802 existing in the buffer at time 0 (X0), Abuffer (t) is described by the following equation:  Thus, the CLefflux value can be obtained by fitting Abuffer (t) versus AUCcell (0 –t) using a least-squares regression program (MULTI) (Yamaoka et al., 1981).
Thus, the CLefflux value can be obtained by fitting Abuffer (t) versus AUCcell (0 –t) using a least-squares regression program (MULTI) (Yamaoka et al., 1981).
Efflux of Ro 64-0802 from the Cerebral Cortex of Wild-Type,Oat3–/–,andMrp4–/–Mice after Microinjection. The efflux of the test compounds from the brain after their microinjection into the cerebral cortex was investigated using the brain efflux index method, as described previously (Kakee et al., 1996). Ro 64-0802 (1 mM) in 0.5 μl of ECF buffer (122 mM NaCl, 25 mM NaHCO3, 10 mM d-glucose, 3 mM KCl, 1.4 mM CaCl2, 1.2 mM MgSO4, 0.4 mM K2HPO4, and 10 mM HEPES, pH 7.4) was injected into the cerebral cortex (4.5 mm lateral to the bregma and 2.5 mm in depth). After the intracerebral microinjection, the mice were decapitated, and the amount of Ro 64-0802 that remained in the ipsilateral cerebrum was determined with LC-MS analysis.
Brain/Plasma Concentration Ratio of Ro 64-0802 after Subcutaneous Infusion of Oseltamivir or Ro 64-0802 in Wild-Type,Oat3–/–, andMrp4–/–Mice. Male C57BL/6J, Mrp4–/–, and Oat3–/– mice (10–18 weeks old), weighing approximately 25 to 30 g, were used for these experiments. An osmotic pump (8 μl/h; Alzet, Cupertino, CA) was implanted under the skin in the backs of the mice under pentobarbital anesthesia (30 mg/kg). The mice received a continuous subcutaneous infusion of oseltamivir or Ro 64-0802 at doses of 400 or 80 nmol/h/mouse, respectively. Blood samples were collected from the postcaval vein at 24 h after treatment under pentobarbital anesthesia, and the brain was excised immediately. Plasma was obtained by centrifugation of the blood samples (10,000g). The esterase inhibitor, dichlorvos (200 μg/ml), was used to prevent ex vivo hydrolysis of the oseltamivir to Ro 64-0802 in the blood and plasma (Wiltshire et al., 2000; Lindegardh et al., 2006). The plasma and brain concentrations of Ro 64-0802 were determined with LC-MS analysis.
Quantification of Ro 64-0802 in Plasma and Brain Specimens. The brain was homogenized with a 4-fold volume of phosphate-buffered saline to obtain a 20% brain homogenate. Plasma specimens (10 μl) were mixed with 40 μl of ethanol, and the brain homogenates (100 μl) were mixed with 400 μl of ethanol. All of these mixed solutions were centrifuged at 15,000g for 10 min. The supernatants of the plasma specimens were mixed with an equal volume of water and subjected to LC-MS analysis. The supernatants of the brain specimens (350 μl) were evaporated, and the pellets were reconstituted with 50 μl of 20% ethanol. The reconstituted specimens were centrifuged at 15,000g for 10 min, and an aliquot of the supernatant was subjected to LC-MS analysis.
An LCMS2010EV equipped with a Prominence LC system (Shimadzu, Kyoto, Japan) was used for the analysis. Samples were separated on a CAP-CELL PAK C18 MGII column (3 μm, 2 mm × 50 mm; Shiseido, Tokyo, Japan) in binary gradient mode at a flow rate of 1 ml/min. Formic acid (0.05%) and acetonitrile were used for the mobile phase. The acetonitrile concentration was initially 10% and then increased linearly to 40% over 2 min. Finally, the column was reequilibrated at an acetonitrile concentration of 10% for 3 min. The total run time was 5 min. Ro 64-0802 was eluted at 2.5 min. In the mass analysis, Ro 64-0802 was detected at a mass-to-charge ratio of 285.15 under positive electron spray ionization conditions. The interface voltage was –3.5 kV, and the nebulizer gas (N2) flow was 1.5 l/min. The heat block and curved desolvation line temperatures were 200 and 150°C, respectively.
Statistical Analysis. Data are presented as means ± S.E. of three to six animals, unless otherwise specified. Student's two-tailed unpaired t test and one-way ANOVA followed by Tukey's multiple comparison test were used to identify significant differences between groups when appropriate. Statistical significance was set at P < 0.05.
Results
Uptake of Ro 64-0802 into Human OAT3-Expressing HEK293 Cells. To show that Ro 64-0802 is an OAT3 substrate, an in vitro transport experiment was performed. The intracellular accumulation of Ro 64-0802 was significantly greater in HEK293 cells expressing human OAT3 than in mock-transfected cells (1.2 ± 0.2 and 15.1 ± 0.2 μl/10 min/mg protein for vector-transfected and OAT3-expressing HEK293 cells, respectively).
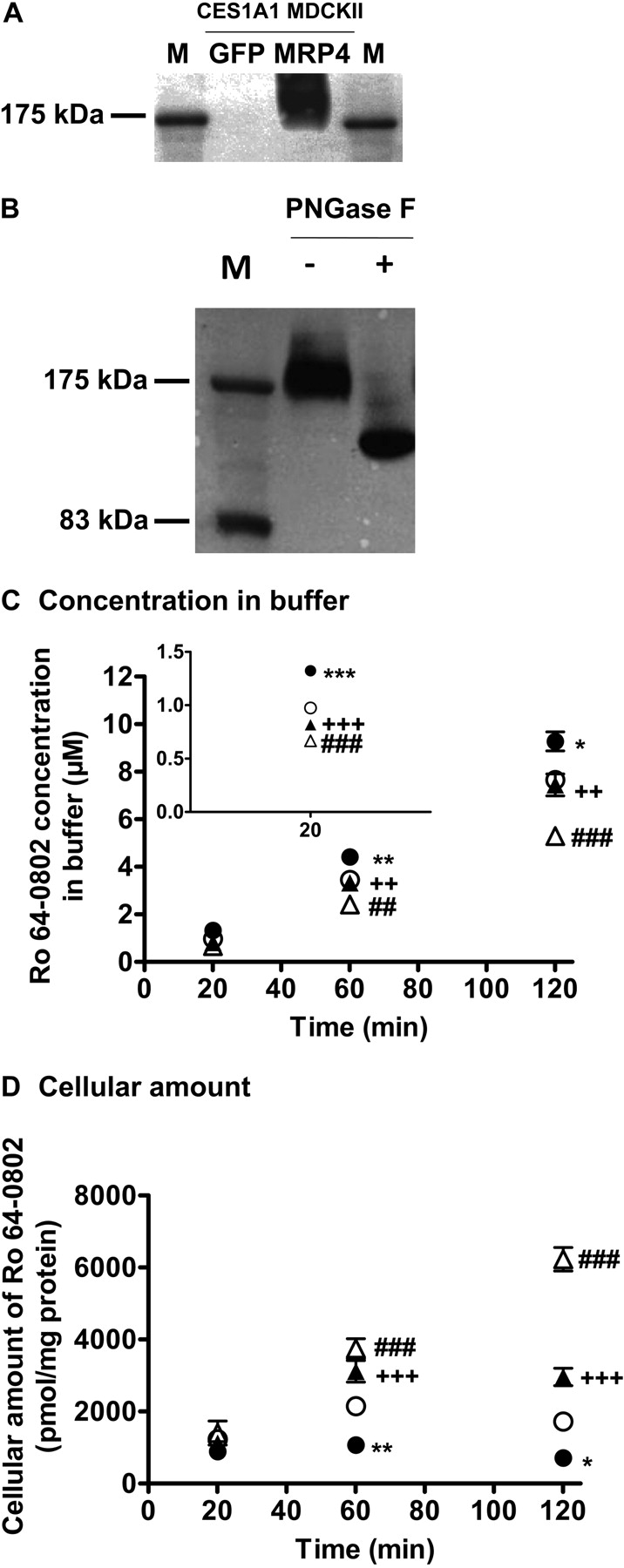
Construction of Human MRP4/CES1A1-Expressing MDCKII Cells. A clone of the MDCKII cells exogenously expressing CES1A1 was selected by measuring the hydrolytic activity against p-nitrophenyl acetate (data not shown). The subsequent study was performed using this clone as the host. After infection with the recombinant adenovirus, the protein expression of MRP4 in the MRP4/CES1A1-expressing MDCKII cells was confirmed by Western blot analysis (Fig. 1A). An anti-MRP4 monoclonal antibody recognized a 175-kDa protein, which is larger than the molecular mass for MRP4 (149 kDa) predicted from the sequence in the Swiss-Prot database. Deglycosylation of the cell lysates with N-glycosidase F resulted in a reduction in the molecular mass, suggesting that the molecular mass of MRP4 in the MRP4/CES1A1-expressing MDCKII cells was increased by glycosylation (Fig. 1B).
Efflux of Ro 64-0802 Formed Intracellularly from Oseltamivir in Human GFP/CES1A1 and MRP4/CES1A1-Expressing MDCKII Cells. An efflux transport experiment was conducted using MRP4/CES1A1- and GFP/CES1A1-MDCKII cells. When MRP4/CES1A1- and GFP/CES1A1-MDCKII cells were incubated with oseltamivir, Ro 64-0802 was detected in both the buffer and the cells in a time-dependent manner (Fig. 1, C and D). The concentration of Ro 64-0802 in the buffer was higher for the MRP4/CES1A1-MDCKII cells than for the GFP/CES1A1-MDCKII cells, and cellular Ro 64-0802 was lower in the MRP4/CES1-MDCKII cells (Fig. 1, C and D). Indomethacin, an inhibitor of MRP4 (Reid et al., 2003; Nozaki et al., 2007), reversed the effects of exogenous MRP4 expression. Integration plots of the efflux transport of Ro 64-0802 are shown in Fig. 2A. The efflux clearance of Ro 64-0802 from MRP4/CES1A1-MDCKII cells was 3.1-fold greater than that from GFP/CES1A1-MDCKII cells and was significantly inhibited by indomethacin (Fig. 2B).
Efflux of Ro 64-0802 from the Cerebral Cortex of Wild-Type andOat3–/–Mice after Microinjection. Real-time PCR was performed to check the adaptive regulation of efflux transporters at the BBB of Oat3–/– mice. There were no significant differences in the mRNA expression of Mdr1a, Bcrp, Mrp4, organic anion transporter peptide 1a4 (Oatp1a4), or Oatp1c1 in the cerebral cortex, quantified by real-time PCR, between wild-type and Oat3–/– mice (data not shown).
To examine the involvement of Oat3 in the efflux transport of Ro 64-0802 across the BBB, Ro 64-0802 was directly injected into the mouse cerebral cortex, and the amount of Ro 64-0802 remaining in the brain was determined at 60 and 120 min after injection. The amount of Ro 64-0802 remaining in the brain was compared between wild-type and Oat3–/– mice (Fig. 3). As shown in Fig. 3, Oat3–/– mice exhibited delayed elimination of Ro 64-0802 from the brain compared with that in wild-type mice.

Efflux of intracellularly formed Ro 64-0802 from mock-transfected and MRP4-expressing MDCKII cells. A and B, Western blotting. A, cell lysates were prepared from mock-transfected (GFP/CES1A1-MDCKII) and MRP4-expressing MDCKII cells (MRP4/CES1A1-MDCKII) and subjected to SDS-polyacrylamide gel electrophoresis (7.5%). B, N-linked carbohydrate groups were cleaved from the MRP4 protein in the cell lysates using N-glycosidase F (PNGase F). MRP4 was detected by the monoclonal anti-MRP4 M4I-10 antibody. C and D, efflux transport study: MRP4/CES1A1-MDCKII (•, ▴) and GFP/CES1A1-MDCKII cells (○, ▵) were incubated with 10 μM oseltamivir in the presence (▴, ▵) or absence (•, ○) of indomethacin (50 μM) at 37°C. Each point represents the mean ± S.E. (n = 6). Statistical significance was calculated by one-way ANOVA followed by Tukey's multiple comparison test. *, P < 0.05; **, P < 0.01; ***, P < 0.001; significantly different between MRP4/CES1A1-MDCKII and GFP/CES1A1-MDCKII cells. ##, P < 0.01; ###, P < 0.001; significantly different in GFP/CES1A1-MDCKII cells with and without indomethacin. ++, P < 0.01; +++, P < 0.001; significantly different in MRP4/CES1A1-MDCKII cells with and without indomethacin.
Efflux of Ro 64-0802 from the Cerebral Cortex of Wild-Type andMrp4–/–Mice after Microinjection. Real-time PCR was used to check the adaptive regulation of efflux transporters at the BBB of Mrp4–/– mice. No significant differences were observed in the mRNA expression levels of Mdr1a, Bcrp, Oat3, Oatp1a4, or Oatp1c1 in the cerebral cortex, quantified by real-time PCR, between wild-type and Mrp4–/– mice (data not shown).
To examine the involvement of Mrp4 in the efflux transport of Ro 64-0802 across the BBB, Ro 64-0802 was directly injected into the mouse cerebral cortex, and the amount of Ro 64-0802 remaining in the brain was determined at 60 and 120 min after injection. The amount of Ro 64-0802 remaining in the brain was compared between wild-type and Mrp4–/– mice (Fig. 4). As shown in Fig. 4, Mrp4–/– mice exhibited delayed elimination of Ro 64-0802 from the brain compared with that in wild-type mice.
Brain/Plasma Concentration Ratio of Ro 64-0802 after Subcutaneous Infusion of Oseltamivir or Ro 64-0802 in Wild-Type,Oat3–/–, andMrp4–/–Mice. To clarify the importance of the Oat3- and Mrp4-mediated efflux of Ro 64-0802 at the BBB, Ro 64-0802 was given to mice by subcutaneous infusion for 24 h, and the Kp,brain of Ro 64-0802 was determined. The concentrations of Ro 64-0802 in the plasma were 4.5 ± 0.6 and 4.5 ± 1.3 μM in wild-type and Oat3–/– mice, respectively. The Kp,brain of Ro 64-0802 in Oat3–/– mice was not significantly different from that in wild-type mice and remained close to the capillary volume in the brain (Fig. 5). After subcutaneous infusions of oseltamivir in wild-type or Mrp4–/– mice, the plasma concentrations of Ro 64-0802 were 6.9 ± 2.3 and 12 ± 5 μM, respectively. The Kp,brain of Ro 64-0802 was 3.8-fold higher in Mrp4–/– mice than that in wild-type mice (Fig. 6). Even after subcutaneous infusions of Ro 64-0802, the Kp,brain of Ro 64-0802 was 6.4-fold greater in Mrp4–/– mice than that in wild-type mice (Fig. 6). The plasma concentrations of Ro 64-0802 were 5.7 ± 0.8 and 3.8 ± 1.7 μM in wild-type and Mrp4–/– mice treated with Ro 64-0802, respectively.

Integration plots of the efflux transport of Ro 64-0802 from MRP4/CES1A1-MDCKII and GFP/CES1A1-MDCKII cells. A, integration plot: the amount of Ro 64-0802 in buffer [Abuffer (t)] was plotted against AUCcell (0 –t) in MRP4/CES1A1-MDCKII (•, ▴) and GFP/CES1A1-MDCKII cells (○, ▵) in the presence (▴, ▵) or absence (•, ○,) of indomethacin (50 μM). The data used for the calculation are cited in Fig. 1, C and D. The slope of the plot represents the efflux clearance. Each point represents a mean ± S.E. (n = 6). B, efflux clearance of Ro 64-0802: The efflux clearance of Ro 64-0802 was calculated from the slope of the integration plot (A). Statistical significance was calculated by one-way ANOVA followed by Tukey's multiple comparison test. ***, P < 0.001; significantly different between MRP4/CES1A1-MDCKII and GFP/CES1A1-MDCKII cells. †††, P < 0.001; significantly different efflux clearance in the presence and absence of indomethacin. Data represent means ± computer-calculated S.D. (microliters per minute per milligram of protein).

Comparison of the amounts of Ro 64-0802 in the ipsilateral cerebrum after intracerebral microinjection of Ro 64-0802 in wild-type (WT) and Oat3–/– mice. Ro 64-0802 (1 mM) in 0.5 μl of ECF buffer was injected into the Par2 region (4.5 mm lateral to the bregma and 2.5 mm in depth). The amount of Ro 64-0802 in the ipsilateral cerebrum was determined at 60 and 120 min after treatment. □, data for wild-type mice; ▪, data for Oat3–/– mice. Each bar represents the mean ± S.E. (n = 3–4). Statistically significant differences between wild-type and Oat3–/– mice: *, P < 0.05; ***, P < 0.001.
Discussion
Abnormal behavior is a suspected adverse effect of oseltamivir on the central nervous system. To understand the pharmacological action of oseltamivir in the brain, its uptake and efflux transport across the BBB were investigated as factors that determine its exposure to the central nervous system. In this study, we focused on Oat3 and Mrp4 as transporters of Ro 64-0802, a pharmacologically active form of oseltamivir, across the BBB.
First, we examined the involvement of Oat3 in the elimination of Ro 64-0802 from the cerebral cortex after microinjection because an in vitro transport study using OAT3-expressing HEK cells identified Ro 64-0802 as a substrate of OAT3. The elimination of Ro 64-0802 from the brain after its microinjection into the cerebral cortex was markedly delayed in Oat3–/– mice compared with wild-type mice (Fig. 3). This suggests that Oat3 plays a significant role in the efflux of Ro 64-0802 from the brain by facilitating its uptake from the brain interstitial space to endothelial cells. For the directional efflux of Ro 64-0802 from the brain to the blood across the BBB, a transporter(s) is also required to facilitate its luminal efflux, considering the hydrophilic nature of Ro 64-0802. In a previous study, we demonstrated that the brain concentrations of Ro 64-0802 in Mdr1a/1b–/– and Abcg2–/– mice are similar to that in wild-type mice (Ose et al., 2008), excluding the possibility that P-gp and Bcrp are involved in the luminal efflux of Ro 64-0802. Therefore, we focused on Mrp4, another ATP-binding cassette transporter at the BBB, as a candidate transporter because it has been reported to mediate the active efflux of topotecan and adefovir across the BBB (Leggas et al., 2004; Belinsky et al., 2007). To show that Ro 64-0802 is a substrate of MRP4, we constructed double transfectant cells expressing both CES1A1, an enzyme producing Ro 64-0802 from oseltamivir, and MRP4 (MRP4/CES1A1-MDCKII). The double transfectant exhibited enhanced efflux of Ro 64-0802 (which was inhibited by an MPR4 inhibitor, indomethacin) compared with GFP/CES1A1-MDCKII (Fig. 2B). It should be noted that the host cells also exhibited indomethacin-sensitive efflux of Ro 64-0802. This is presumably attributable to endogenous canine MRP4 because its mRNA expression was detected by reverse transcription-PCR in MDCKII cells (data not shown). The involvement of Mrp4 in the efflux of Ro 64-0802 across the BBB was then examined using Mrp4–/– mice by microinjection into the cerebral cortex. Mrp4–/– mice exhibited delayed elimination of Ro 64-0802 from the cerebral cortex after its microinjection compared with that in wild-type mice (Fig. 4). Therefore, Mrp4 plays an important role in the luminal efflux of Ro 64-0802 after its cellular uptake by Oat3 from the brain side.

Comparison of the amounts of Ro 64-0802 in the ipsilateral cerebrum after intracerebral microinjection of Ro 64-0802 in wild-type (WT) and Mrp4–/– mice. Ro 64-0802 (1 mM) in 0.5 μl of ECF buffer was injected into the Par2 region (4.5 mm lateral to the bregma and 2.5 mm in depth). The amount of Ro 64-0802 in the ipsilateral cerebrum was determined at 60 and 120 min after treatment. □, data for wild-type mice; ▪, data for Mrp4–/– mice. Each bar represents the mean ± S.E. (n = 3–4). Statistically significant differences between wild-type and Mrp4–/– mice: **, P < 0.01; ***, P < 0.001.

Comparison of the brain/plasma concentration ratio (Kp,brain) of Ro 64-0802 after subcutaneous infusion of Ro 64-0802 in wild-type and Oat3–/– mice. Mice received a continuous subcutaneous infusion of Ro 64-0802 at a dose of 80 nmol/h/mouse for 24 h, with an osmotic pump. The plasma and brain concentrations of Ro 64-0802 were determined at 24 h after treatment. □, data for wild-type mice; ▪, data for Oat3–/– mice. Each bar represents the mean ± S.E. (n = 3–4).

Comparison of the brain/plasma concentration ratio (Kp,brain) of Ro 64-0802 after subcutaneous infusion of either oseltamivir or Ro 64-0802 in wild-type (WT) and Mrp4–/– mice. Mice received a continuous subcutaneous infusion of either oseltamivir or Ro 64-0802 at a dose of 400 or 80 nmol/h/mouse, respectively, for 24 h with an osmotic pump. The plasma and brain concentrations of Ro 64-0802 were determined at 24 h after treatment. □, data for wild-type mice; ▪, data for Mrp4–/– mice. Each bar represents the mean ± S.E. (n = 4–6). Statistically significant differences between wild-type and Mrp4–/– mice: **, P < 0.01; ***, P < 0.001.
To show the importance of the active efflux of Ro 64-0802 at the BBB mediated by Oat3 and Mrp4, the Kp,brain of Ro 64-0802 was determined in Oat3–/– or Mrp4–/– mice given Ro 64-0802 by subcutaneous infusion for 24 h. Our approach was based on the pharmacokinetic concept that reduced efflux across the BBB may lead to an increase in the Kp,brain. Consistent with a previous report (Ose et al., 2008), the Kp,brain of Ro 64-0802 was close to the capillary volume in wild-type mice. The Kp,brain of Ro 64-0802 observed in Oat3–/– mice was also close to the capillary volume (Fig. 5). In contrast, the Kp,brain of Ro 64-0802 was 4- to 6-fold greater in Mrp4–/– mice than that in wild-type mice receiving either oseltamivir or Ro 64-0802 (Fig. 6). These in vivo results using Mrp4–/– mice thus indicate that Ro 64-0802 crosses the BBB from the blood side to the brain, but Mrp4 limits its penetration into the brain by extruding it into the blood. The latter finding is consistent with the results of our microinjection experiment. However, the lack of effect of knockout of Oat3 on the Kp,brain of Ro 64-0802 seems to contradict the results of the microinjection experiment. There are three explanations for this discrepancy. The first explanation is that luminal Mrp4 is more important in preventing brain penetration than abluminal Oat3. The second explanation is that, considering the mediation of bidirectional transport by Oat3 (Bakhiya et al., 2003), it is possible that Oat3 on the abluminal membrane of brain endothelial cells may act in the efflux of Ro 64-0802 from inside the endothelial cells into the brain, as well as in its uptake from the brain to the endothelial cells (Fig. 7). The third explanation is that, considering the hydrophilic character of Ro 64-0802, with an anionic charge, its luminal uptake probably involves transporters. Kikuchi et al. (2003) suggested that Oat3 is expressed on both the luminal and abluminal membranes of rat brain capillaries; however, this localization is controversial (Mori et al., 2003; Roberts et al., 2008). Oat3 may serve as an uptake system for Ro 64-0802 on the luminal membrane from the circulating blood into the brain (Fig. 7). Further studies using Mrp4 and Oat3 double knockout mice can answer these questions and perhaps confirm these speculations.

Schematic representation of the proposed mechanism underlying the brain distribution of oseltamivir and Ro 64-0802 in humans. Oseltamivir crosses the BBB, and Ro 64-0802 is produced by CES1A1 during the penetration of oseltamivir across the BBB. Both oseltamivir and Ro 64-0802 are subjected to active efflux by P-gp and MRP4, respectively. The functional importance of OAT3 at the human BBB has not yet been established, so the hypothetical pathway is shown with a broken line.
Figure 7 summarizes the proposed mechanisms determining the brain distribution of oseltamivir and Ro 64-0802 in humans. P-gp, MRP4, and CES1A1 are factors that determine the brain distribution of oseltamivir and Ro 64-0802 in humans. CES1A1 is predominantly expressed in the brain capillaries in human brain (Yamada et al., 1994), although whether CES1A1 is the only enzyme responsible for the conversion of oseltamivir remains to be examined. MRP4 is also expressed on the luminal membranes of human brain capillaries (Nies et al., 2004; Bronger et al., 2005). Because oseltamivir crosses the BBB, Ro 64-0802 can be produced during the penetration of oseltamivir across the BBB and then be subjected to active efflux by MRP4. Although Northern blotting did detect OAT3 mRNA expression in the human brain (Cha et al., 2001), its distribution and membrane localization remain to be determined. Therefore, whether Ro 64-0802 can penetrate into the brain and is eliminated from the brain by OAT3 in humans, as well as in mice, also remains in question. Fluctuations in their activities will cause interindividual variations in their exposure to the central nervous system. For instance, genetic variations have been reported for P-gp and MRP4, which may alter their transport activities and expression. The silent mutation 3435C>T is associated with reduced P-gp expression (Hoffmeyer et al., 2000) and affects protein folding, resulting in a substrate-dependent functional change (Kimchi-Sarfaty et al., 2007). In MRP4, the nonsynonymous mutations 559G>T, 1460G>A, and 2269G>A are associated with altered transport activity and expression (Abla et al., 2008; Krishnamurthy et al., 2008). It is possible that these polymorphisms are associated with the adverse effects of Ro 64-0802 on the central nervous system.
In conclusion, Mrp4 and Oat3 are responsible for the elimination of Ro 64-0802 from the brain across the BBB although, at steady-state, Oat3 may not affect its brain distribution, probably because of its possible contribution also to the brain uptake of Ro 64-0802. This is the first demonstration of the cooperation of uptake and efflux transporters in the directional (brain-to-blood) transport of anionic drugs across the BBB.
Acknowledgments
We thank Dr. Junko Iida and Futoshi Kurotobi (Shimadzu) for technical support with the LC-MS system.
Footnotes
-
This work supported in part by a grant-in-aid for Scientific Research (A) [Grant 20249008] and Scientific Research (B) [Grant 20390046] from the Ministry of Education, Culture, Sports, Science and Technology.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.108.024018.
-
ABBREVIATIONS: Ro 64-0802, [3R,4R,5S]-4-acetamido-5-amino-3-(1-ethylpropoxy)-1-cyclohexene-1-carboxylate phosphate; BBB, blood-brain barrier; CES1A1, carboxylesterase 1A1; P-gp, P-glycoprotein; Oat/OAT, organic anion transporter; Mrp/MRP, multidrug resistance-associated protein; HEK, human embryonic kidney; MDCK, Madin-Darby canine kidney; LC, liquid chromatography; MS, mass spectrometry; GFP, green fluorescent protein; ANOVA, analysis of variance; PCR, polymerase chain reaction; Mdr, multidrug resistance; Bcrp, breast cancer resistance protein; Oatp, organic anion transporter peptide.
- Received August 18, 2008.
- Accepted November 20, 2008.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}