


Abstract
The recently introduced Clonetics human corneal epithelium (cHCE) cell line is considered a promising in vitro permeability model, replacing excised animal cornea to predict corneal permeability of topically administered compounds. The purpose of this study was to further characterize cHCE as a corneal permeability model from both drug metabolism and transport aspects. First, good correlation was found in the permeability values (Papp) obtained from cHCE and rabbit corneas for various ophthalmic drugs and permeability markers. Second, a previously established real-time quantitative polymerase chain reaction method was used to profile mRNA expression of drug-metabolizing enzymes (major cytochromes P450 and UDP glucuronosyltransferase 1A1) and transporters in cHCE in comparison with human cornea. Findings indicated that 1) the mRNA expression of most metabolizing enzymes tested was lower in cHCE than in excised human cornea, 2) the mRNA expression of efflux transporters [multidrug resistant-associated protein (MRP) 1, MRP2, MRP3, and breast cancer resistance protein], peptide transporters (PEPT1 and PEPT2), and organic cation transporters (OCTN1, OCTN2, OCT1, and OCT3) could be detected in cHCE as in human cornea. However, multidrug resistance (MDR) 1 and organic anion transporting polypeptide 2B1 was not detected in cHCE; 3) cHCE was demonstrated to possess both esterase and ketone reductase activities known to be present in human cornea; and 4) transport studies using probe substrates suggested that both active efflux and uptake transport may be limited in cHCE. As the first detailed report to delineate drug metabolism and transport characteristics of cHCE, this work shed light on the usefulness and potential limitations of cHCE in predicting the corneal permeability of ophthalmic drugs, including ester prodrugs, and transporter substrates.
Topical instillation is the desired route of administration for ophthalmic drugs to treat diseases in the anterior segment of the eye including glaucoma, inflammations, infections, and dry eye. The primary pathway of drug permeation from the tear fluid to the anterior chamber of the eye is via the transcorneal route. The cornea has a multilayered structure constituted primarily of corneal epithelium, stroma, and endothelium. However, passage through the corneal epithelium is considered to be the rate-limiting step in the transcorneal penetration of most ophthalmic drugs (Maurice and Mishima, 1984).
In the past decade, several in vitro corneal permeability models that can substitute for the isolated cornea to predict ocular absorption and facilitate ophthalmic drug discovery have been developed (Reichl and Becker, 2008). These models evolved from primary cultures using rabbit corneas and immortalized rabbit cell lines to the immortalized human corneal epithelial (HCE) cell lines, and even include complex systems such as human corneal constructs (summarized in Table 1). Given the pros and cons of each model, HCE cell lines seem to be most appealing owing to their human origin, the relatively short preparation time, and the good reproducibility often ensured by commercially available products. A recent comparative study suggested that the Clonetics cHCE could be more suitable than other HCE cell lines (i.e., HCE-T and sHCE) in predicting ocular drug permeability. This model not only closely resembles excised human corneas in the barrier function as measured by TEER but also seems to be able to adequately differentiate permeability differences observed between poorly and highly permeable compounds (Becker et al., 2008).
Summary of in vitro models used for drug corneal permeability assessment
Not only does the cornea serve as a diffusion barrier to limit drug penetration, it also expresses certain drug-metabolizing enzymes and active transport systems (Duvvuri et al., 2004; Mannermaa et al., 2006; Zhang et al., 2008). These enzymes and transporters, when functionally expressed, could affect ocular drug absorption. For example, the corneal epithelium has robust esterase activity, which underlies the significance of the ester prodrug approach in ophthalmic drug design (Shirasaki, 2008). For the efflux and uptake transporters potentially present in the cornea, the ocular absorption of their drug substrates could be either hindered or facilitated. Because of the restricted tissue availability of fresh human cornea, mechanistic understanding about the effect of metabolic conversion and drug-transporter interaction on ocular drug absorption is lacking. Encouragingly, the newly introduced cHCE model could be an appropriate in vitro system to study these mechanisms, allowing for more flexible and in-depth experimental designs that are otherwise prohibitive with fresh human cornea.
As described here, studies were designed to further characterize cHCE as a corneal permeability model, particularly from drug metabolism and transport aspects. In brief, a previously established real-time quantitative PCR approach (Zhang et al., 2008) was adopted to profile mRNA expression of key metabolizing enzymes and transporters in cHCE in comparison with human cornea. Moreover, functional studies using probe substrates were conducted to assess drug metabolism (i.e., esterase and ketone reductase) and active transport in cHCE.
Materials and Methods
Materials. Acetazolamide, amantadine, ampicillin, atenolol, brimonidine, buspirone (internal standard), cimetidine, desipramine, dexamethasone, dexamethasone 21-acetate, digoxin, diclofenac, etoposide, latanoprost, latanoprost acid, methotrexate, metoprolol, nadolol, NADPH, pilocarpine, quinidine, and timolol were purchased from Sigma-Aldrich (St. Louis, MO). The cHCE on Transwell filter inserts was purchased from Lonza Walkersville, Inc. (Walkersville, MD). Fresh rabbit corneas (albino) were purchased from Pel-Freez Biologicals (Rogers, AR), kept in Optisol-GS (Bausch & Lomb, Rochester, NY), and used within 36 h after harvesting. BSS and BSS PLUS solution was purchased from Alcon (Dallas, TX).
RNA Extraction, cDNA Synthesis, and Real-Time Quantitative PCR. Human eye tissue collection, RNA extraction, cDNA synthesis, real-time PCR, and quantitative analysis were performed as reported previously (Zhang et al., 2008). Total RNA from cHCE cells was extracted with the QIAGEN RNeasy kit after homogenization in a FastPrep F120 instrument (Qbiogene Inc., Irvine, CA). PCR reactions were run in a total volume of 20 μl, containing cDNA from 25 ng of total RNA. The geometric mean of human β-actin, glyceraldehyde-3-phosphate dehydrogenase, and peptidylpropyl isomerase A was used as the normalization factor for the relative quantification of each gene.
Permeability Studies with the Excised Rabbit Cornea and cHCE.Excised rabbit cornea. Fresh rabbit corneas were rinsed and equilibrated in BSS solution for 15 min. The corneas were mounted in a Ussing chamber perfusion system (Harvard Apparatus Inc., Holliston, MA), which was placed in the heating block (Harvard Apparatus Inc.). The exposed surface area of the cornea is approximately 0.2 cm2/well. One milliliter of BSS or BSS PLUS solution was added in the chamber bathing the endothelium side (the receiver chamber), and 1 ml of test compound (10 μM) in BSS or BSS PLUS was added in the chamber bathing the epithelium side (the donor chamber). Constant mixing of the donor and receiver solutions was achieved with a gentle gas flow of air-CO2 (95%:5%) through both chambers. The heating block was maintained at 37°C by a circulating water bath. Serial samples of 100 μl were removed from the receiver chamber every hour during a period of 4 h. Each aliquot was replaced immediately with an equal volume of BSS. The samples, mixed with 10 μl of internal standard solution, were analyzed directly by LC-MS/MS.
cHCE. Upon the receipt of each cHCE plate, the cells were cultured in the medium overnight and used the next day. The surface area of the membrane is 0.33 cm2 per insert. The permeability experiments were performed under a humidified atmosphere of air-CO2 (95%:5%) at 37°C. In the A to B transport study, 1 ml of cell medium was added in the basolateral side (the receiver side). The study was initiated by dosing the apical compartment with 200 μl of cell medium containing the test compound and permeability markers (i.e., nadolol and metoprolol). In the B to A transport study, 200 μl of cell medium was added in the apical side (the receiver side), and the study was initiated by dosing the basolateral compartment with 1000 μl of cell medium containing the test compound and permeability markers. The buffer pH in both donor and receiver chambers was 7.4 for all of the compounds tested except for ampicillin, which was dosed in the cell medium with pH adjusted to 6.5. Samples of 100 μl were collected from the receiver side every 60 min for up to 4 h. Each 100-μl aliquot removed was immediately replenished with an equal volume of fresh cell medium. The samples, mixed with 10 μl of internal standard solution, were analyzed directly by LC-MS/MS.

Permeability values (Papp) obtained in excised rabbit corneas and cHCE. Data are presented as the mean of triplicates.
Permeability Data and Statistical Analysis. Apparent permeability coefficients (Papp) were calculated using the following equation: Papp = (1/A × C0) (dM/dt), where dM/dt is the flux (nanomoles per second) across the cell layers or cornea, A (square centimeters) is the exposed surface area of the insert membrane of cHCE or rabbit cornea, and C0 is the initial drug concentration (micromolar) in the donor compartment at t = 0. Flux (dM/dt) across the cell layer or cornea was determined from the slope of the linear portion of the cumulative amount permeated versus time plot. The results are expressed as the mean ± S.D. The statistical significance (p < 0.05) was determined by Student's t test.
Esterase Activity and Prodrug Permeability in cHCE. Esterase activity of cHCE cells was monitored by the conversion of ester prodrug, latanoprost (10 μM), to the acid form. The permeability value of latanoprost was calculated according to the concentrations of either latanoprost or its acid in the receiver chamber of the cHCE system. In addition, the Papp value of latanoprost acid was calculated when the acid itself was dosed in the donor chamber.
Ketone Reductase Activity in cHCE. Ketone reductase activity was monitored by LC-mass spectrometry detection of dihydrolevobunolol in the receiver side after levobunolol was dosed at 10 μM in the donor chamber.
LC-MS/MS. The analytical system comprised a Sciex API 4000 mass spectrometer (Applied Biosystems, Foster City, CA), a CTC HTS PAL autosampler (LEAP Technologies, Carrboro, NC), and Shimadzu LC-10ADvp pumps (Shimadzu, Kyoto, Japan). The samples were analyzed using reverse-phase chromatography (Zorbax SB-Phenyl, 50 mm × 2.1 mm, 5 μm; Agilent Technologies, Santa Clara, CA). A 4-min linear gradient from 5 to 90% of mobile phase I (acetonitrile-water 98:2, containing 0.1% formic acid) was used at the flow of 0.6 ml/min, and mobile phase II was water-acetonitrile (98:2) containing 0.1% formic acid. Multiple reaction monitoring was used to monitor drug levels in the various experiments. The parent/fragment information is as follows: acetazolamide (223/181), amantadine (152/135), ampicillin (350/106), atenolol (267/145), brimonidine (292/212), buspirone (386/122), cimetidine (253/159), desipramine (267/208), dexamethasone (393/373), dexamethasone 21-acetate(435/415), digoxin (781/651), diclofenac (296/215), dihydrolevobunolol (294/220), dorzolamide (325/199), etoposide (589/229), ethoxyzolamide (259/178), indinavir (614/421), latanoprost (433/397), latanoprost acid (391/355), levobunolol (292/236), methotrexate (455/308), metoprolol (268/116), nadolol (310/254), pilocarpine (209/163), quinidine (325/307), timolol (317/261), and verapamil (455/165).
Results
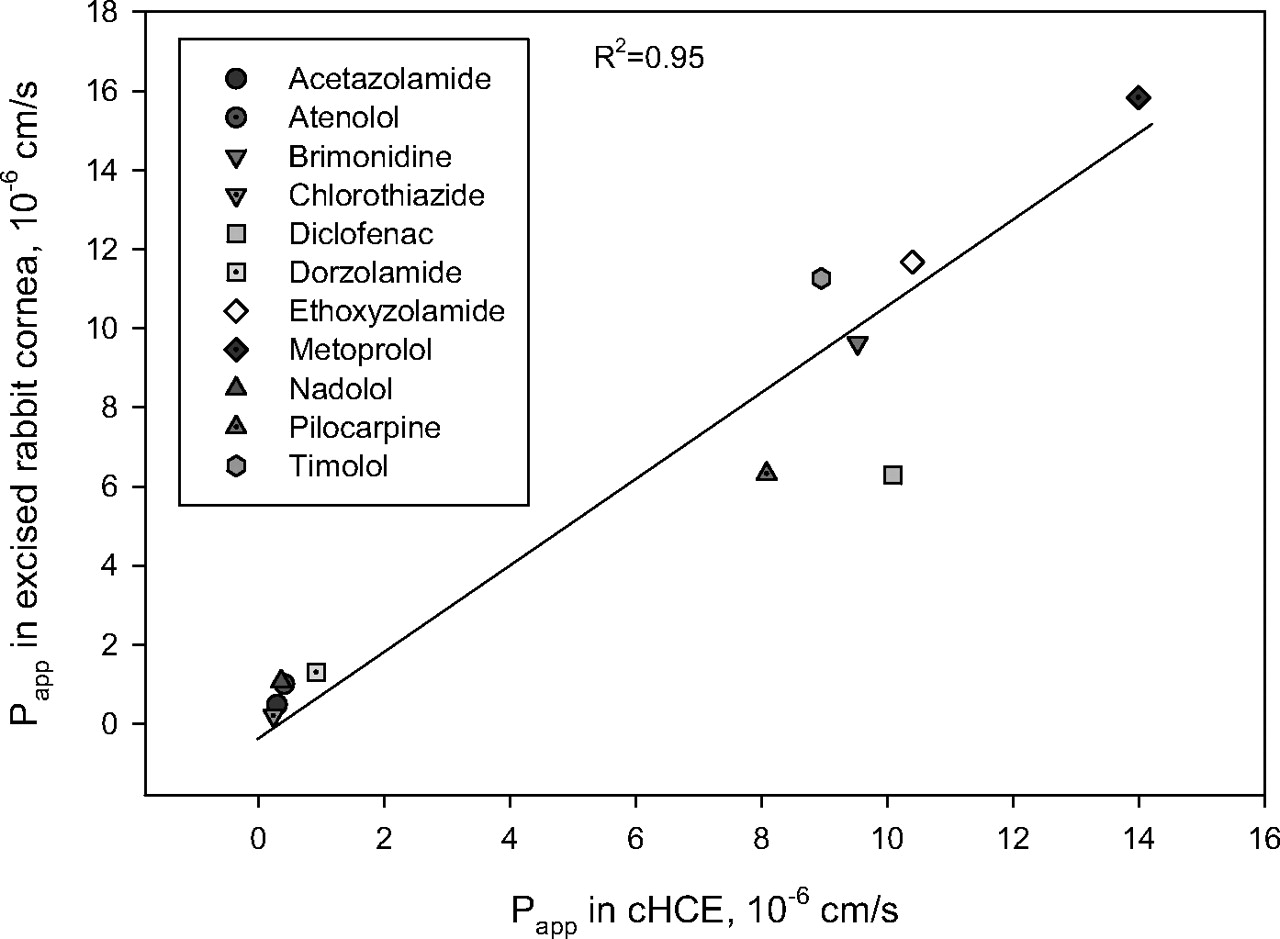
Transcorneal Permeability in Excised Rabbits Corneas and cHCE. The permeability of eight ophthalmic drugs and three β-blockers (atenolol, metoprolol, and nadolol) was assessed using either excised rabbit corneas or cHCE. The Papp values for the 11 compounds track well between values for the two models with a correlation coefficient (R2) of 0.95 (Fig. 1). In addition, Spearman's rank correlation coefficient was calculated to be 0.92, suggesting the similar rank order of permeability data obtained from the two models.
mRNA Expression of Drug-Metabolizing Enzymes in Human Cornea and cHCE. Consistent with the previous report (Zhang et al., 2008), the mRNA level of each detectable metabolizing enzyme in human cornea was less than 1% of the corresponding hepatic level. CYP1A2 and CYP2B6 mRNAs were not detected in either cHCE or human cornea. All of the enzymes tested except CYP2A6 were lower in cHCE than in human cornea (Fig. 2).
mRNA Expression of Drug Transporter in Human Cornea and cHCE. Similar to that in human cornea, the mRNA expression of efflux transporters (MRP1, MRP2, MRP3, and BCRP), peptide transporters (PEPT1 and PEPT2), and organic cation transporters (OCTN1, OCTN2, OCT1, and OCT3) could be detected in cHCE. Their expression levels were more or less comparable between human cornea and cHCE. In addition, the mRNA expression of MDR1 and organic anion-transporting protein 2B1, which was detected in human cornea but quantified as very low (Zhang et al., 2008) was not detected in cHCE (Fig. 3).
Esterase Activity in cHCE and Rabbit Cornea. Esterase activity was evident in both cHCE and rabbit cornea as latanoprost acid was readily detected in the receiver chambers after latanoprost was dosed in the donor chamber. Permeability values of latanoprost (calculated based on the concentrations of either prodrug or acid in the receiver chambers) and latanoprost acid are shown in Table 2. In both models, latanoprost acid had low and similar permeability. Latanaprost exhibited poor permeability when measured as the parent ester itself but had a much higher permeability when measured as latanoprost acid.
Ester conversion and permeability of latanoprost in cHCE and excised rabbit cornea (dosed as either ester or acid)
Data are presented as the mean of triplicates ± S.D.
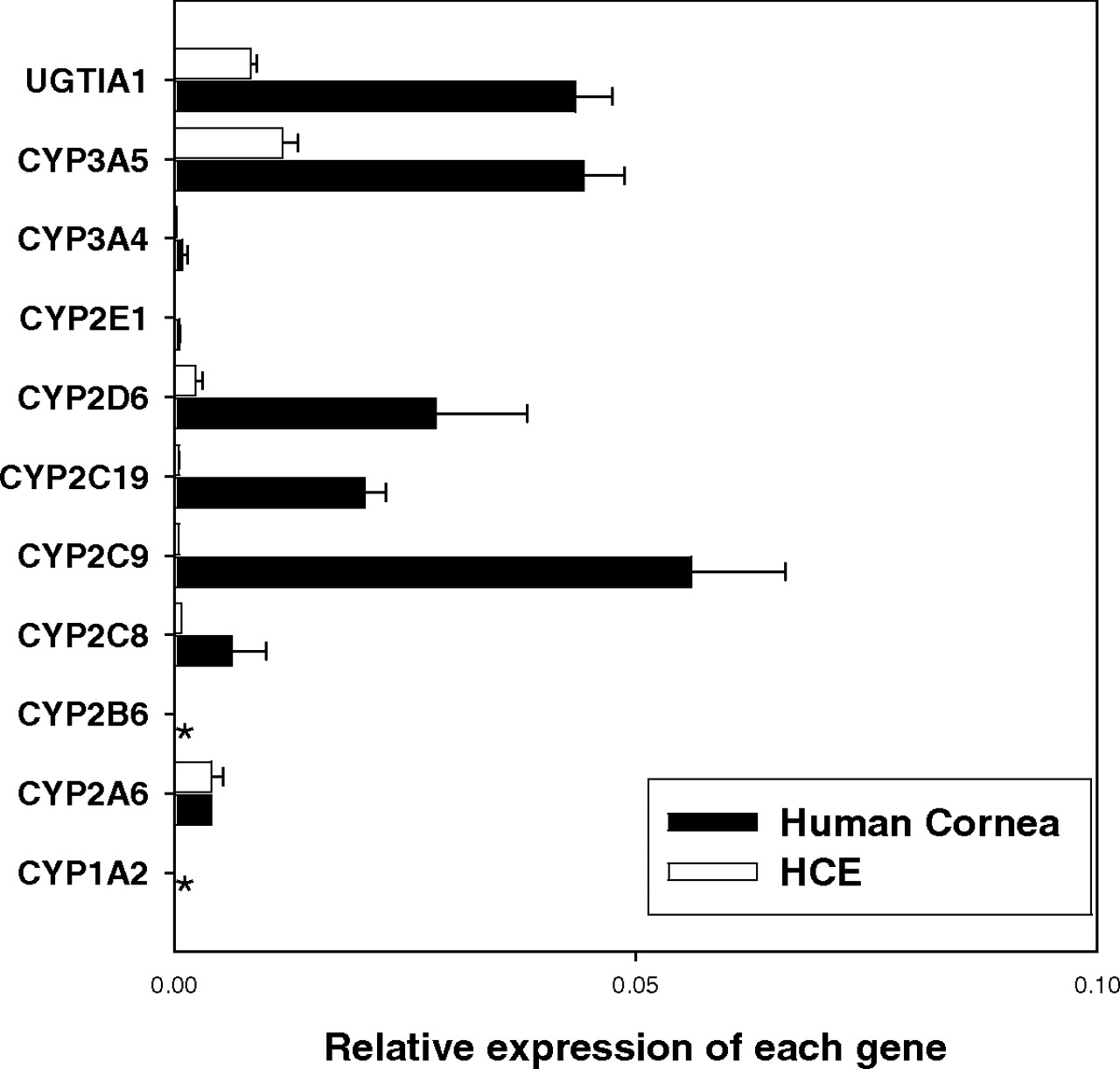
Relative mRNA expression level of major P450s and UGT1A1 in human cornea and cHCE. Data are presented as the mean of triplicates with the S.D. as the error bar. *, not detected.

Relative mRNA expression level of selected (A) drug efflux and (B) uptake transporter genes in human cornea and cHCE. Data are presented as the mean of triplicates with the S.D. as the error bar. *, not detected.
Ketone Reductase Activity in cHCE. Dihydrolevobunolol was readily detected in the receiver side after levobunolol was dosed in the donor side of the cHCE model (reaction shown in Fig. 4). As a nonselective β-adrenergic antagonist, levobunolol is used in the treatment of glaucoma. It has been shown that the metabolic reduction of the cyclohexanone functional group to form dihydrolevobunolol is mediated by NADPH-dependent ketone reductase in ocular tissues including cornea (Lee et al., 1988).
Permeability of Probe Substrates for Efflux Transporters in cHCE. The permeability of digoxin (P-gp substrate) and indinavir (substrate for P-gp, MRP1, and MRP2) in the cHCE model was measured at both A to B and B to A directions under various concentrations (Fig. 5). When measured in either direction, Papp values of either digoxin or indinavir did not seem to vary significantly with the increase in concentration. In addition, the flux ratios did not appear to significantly deviate from unity (i.e., in the range from 0.6 to 1.5) for either compound under different concentrations. The permeability of two MRP3 substrates, etoposide and methotrexate, was also tested but was found to be too low in both directions to derive a Papp ratio (data not shown).
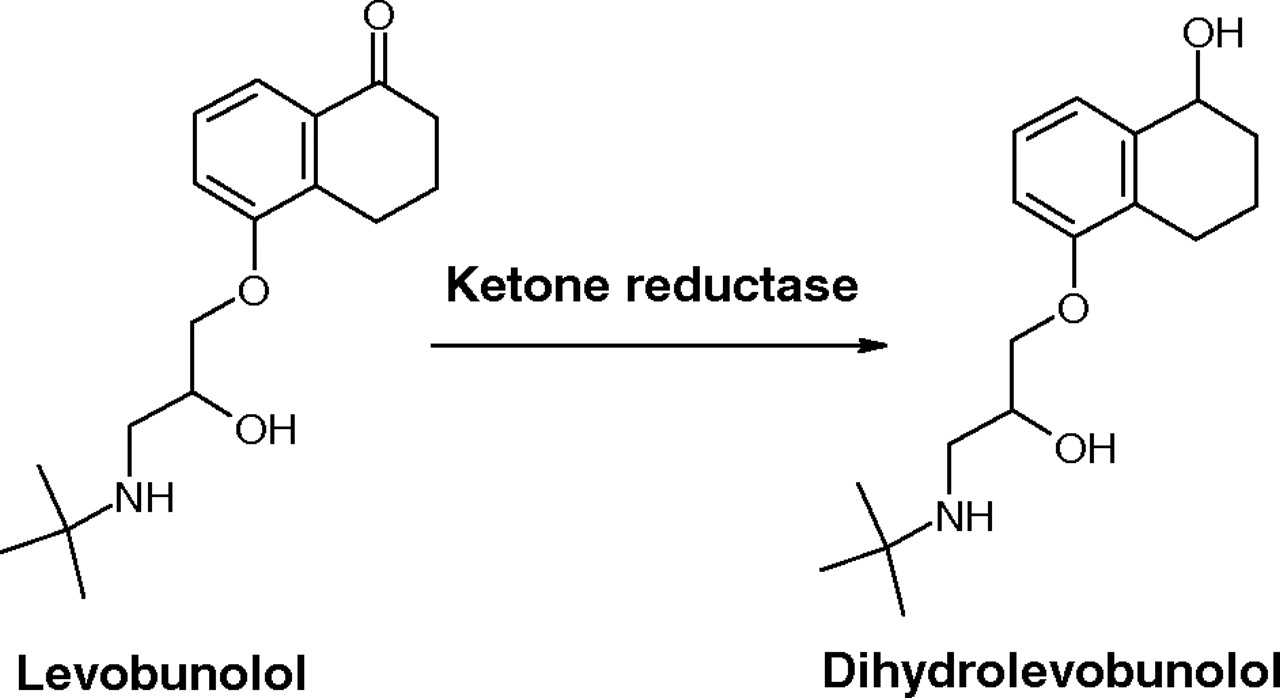
Ketone reductase activity in cHCE as evidenced by the formation of dihydrolevobunolol from levobunolol.
Permeability of Probe Substrates for Uptake Transporters in cHCE. The permeabilities of quinidine and verapamil (substrates for OCTN1 and OCTN2), amantadine and desipramine (substrates for OCT1), cimetidine (substrate for OCT3), and ampicillin (substrate for PEPT1 and PEPT2) in the cHCE model were measured in both A to B and B to A directions at 10 μM (Table 3). At this concentration, the Papp_A to B of quinidine, verapamil, amantadine, desipramine, and cimetidine appeared to be similar to the Papp_B to A, suggesting the lack of active uptake of these compounds in cHCE cells. Papp values of ampicillin were found to be poor in both directions with an apparent higher Papp_B to A (0.56 ± 0.1 × 10–6 cm/s) than Papp_A to B (0.12 ± 0.07 × 10–6 cm/s). It should be noted that the codosed low-permeability marker (i.e., nadolol) also appeared to be more permeable in the B to A direction (Papp_B to A 0.60 ± 0.15 × 10–6 cm/s versus Papp_A to B 0.36 ± 0.15 × 10–6 cm/s). Nevertheless, the data suggested that there may also be little active uptake of ampicillin in cHCE cells.
Papp values and Papp_A to B/Papp_B to A ratios of probe substrates (10 μM) of uptake transporters in cHCE
Data are presented as the mean of triplicates ± S.D.
Discussion
Assessing the transcorneal permeability of drug candidates is essential in ophthalmic drug discovery for front-of-eye delivery. This assessment is often achieved with excised animal (i.e., rabbit) cornea using a diffusion chamber setup, which offers reasonable predictability for human transcorneal permeability. However, such experiments are labor-intensive with low throughput and involve the sacrifice of animals. These disadvantages may be alleviated by cell-based models derived from corneal tissues (Table 1). In this work, we focused on the newly introduced Clonetics cHCE model, which is regarded as a promising in vitro model to minimize the use of animal corneas in transcorneal permeability testing (Beck et al., 2008). Permeability values for 11 compounds were obtained from excised rabbit cornea and cHCE models (Fig. 1). Good correlation between the two models suggests that the cHCE model could indeed be substituted for excised rabbit cornea as a surrogate permeability model in ocular drug discovery.
Human cornea is not simply a physical barrier that limits the diffusion of xenobiotics into the eye. It is equipped with sophisticated metabolic and potential active transport systems that should be taken into consideration for ophthalmic drug delivery and disposition (Duvvuri et al., 2004; Mannermaa et al., 2006). In previous work (Zhang et al., 2008), the mRNA expression of major P450s and drug transporters was profiled for human ocular tissues, including cornea. A similar real-time PCR approach was adopted in this present work to compare the mRNA expression of major metabolizing enzymes and drug transporters between human cornea and the cHCE. Furthermore, functional studies using probe substrates were explored to characterize cHCE as a permeability model from both metabolism and transport aspects.
Drug Metabolism in cHCE. The mRNA expression of most P450s and UGT1A1 tested was shown to be lower in cHCE than in human cornea, which itself possesses limited mRNA expression of these enzymes in comparison with the liver. There was little metabolic turnover when P450s and UGT1A1 probe substrates were incubated in rabbit corneal homogenates (data not shown). Taken together, the data suggest that 1) the human cornea probably possesses minimal metabolic capacity mediated by P450 enzymes and UGT1A1 and 2) although the cHCE cells may contain even lesser amounts of these enzymes than human cornea, the cHCE cells may still be a relevant model to evaluate the transcorneal permeability of substrates for these enzymes.
The cHCE cells were shown to have evident esterase activity to convert latanoprost (Xalatan) to its pharmacologically active metabolite (latanoprost acid), as well as ketone reductase activity to convert levobunolol (Betagan) to its equipotent ocular metabolite, dihydrolevobunolol. It should be noted that although these results confirmed that the esterases and ketone reductase possessed by human cornea are also functional in cHCE cells, it did not provide any quantitative comparison on the rate and extent of these enzyme activities between cHCE cells and human cornea.
Similar permeability data for latanoprost and latanoprost acid were obtained in both cHCE and excised rabbit cornea models (Table 2). Latanoprost acid itself had low permeability, whereas its ester prodrug (latanoprost, quantified in the form of the acid) showed much improved permeability in both cHCE (>40 fold) and excised rabbit corneas (>30 fold). As the prodrug design is a proven approach in ophthalmic drug design (Shirasaki, 2008), our data suggest that the cHCE model may be used to confirm metabolic conversion and evaluate transcorneal permeability of ester prodrug candidates. Further experiments are needed to quantitatively compare esterase activity between cHCE and human cornea in terms of esterase composition, specificity, and kinetics.
Drug Transport in cHCE. The mRNA expression and potential functional presence of drug transporters in human cornea have been documented in the literature (Duvvuri et al., 2004; Mannermaa et al., 2006; Karla et al., 2007; Zhang et al., 2008). In the present work, five efflux and seven uptake transporters, which were found to have quantifiable mRNA expression in human cornea (Zhang et al., 2008), were studied for their mRNA expression (Fig. 3) and functional relevance in cHCE cells (Table 3; Fig. 5). The different probe substrates for the different transporters were based on transporter substrate recommendations found in the U.S. Food and Drug Administration transporter database (http://www.fda.gov/cder/drug/drugInteractions/tableSubstrates.htm#major).
Except for MDR1 (not detectable in cHCE, but detected at a very low level in human cornea), the mRNA expression of other efflux transporters including MRP1, MRP2, MRP3, and BCRP was detected in cHCE cells. Consistent with the lack of MDR1 mRNA expression in cHCE cells, there was no significant unidirectional flux of digoxin (a substrate for P-gp) at different concentrations (Fig. 5). These data are consistent with a study that showed no significant difference in rhodamine 123 (a substrate for P-gp) flux in either direction in excised human cornea or cHCE (Becker et al., 2007, 2008). Although mRNAs of MRP1, MRP2, and BCRP could be detected in cHCE cells, their activities did not appear to manifest in these cells because no significant unidirectional flux of indinavir (a common substrate for these transporters) was observed under different concentrations (Fig. 5).

Permeability parameters of digoxin (A) and indinavir (B) obtained in cHCE. Data are presented as the mean of triplicates with the S.D. as the error bar.
Despite the detectable mRNA expression of uptake transporters (PEPT1, PEPT2, OCT1, OCT3, OCTN1, and OCTN2) in cHCE cells, the mRNA presence does not seem to translate to a net increase in the absorptive flux of their substrates (Table 3), suggesting that the active transport function of these uptake transporters also may be minimal in cHCE cells. It is important to note that the similarities or differences in mRNA expression (Fig. 3) between cHCE and human cornea of these tested efflux and uptake transporters do not necessarily correlate to protein expression and transport activity. Therefore, it is still difficult to conclude whether the cHCE model is predictive or not for human transcorneal permeability of these transporter substrates and whether further studies (i.e., permeability data obtained from freshly excised human cornea) would be needed.
In summary, this work further validated the fact that the cHCE model can be adopted as a surrogate permeability model to substitute for the excised rabbit cornea model in ophthalmic drug discovery and research. The detailed characterization of cHCE as a permeability model from both drug metabolism and transport aspects sheds light on the potential similarities and differences between cHCE and human cornea. These characteristics should be taken into account when cHCE is used to predict transcorneal permeability of drug candidates, including prodrug and transporter substrates.
Acknowledgments
We thank Ping Kang and Deepak Davie for insightful scientific discussions and critical reading of the manuscript.
Footnotes
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.108.026286.
-
ABBREVIATIONS: HCE, human cornea epithelial; TEER, transepithelial electrical resistance; PCR, polymerase chain reaction; BSS, balanced salt solution; LC, liquid chromatography; MS/MS, tandem mass spectrometry; A, apical; B, basolateral; MRP, multidrug resistant-associated protein; BCRP, breast cancer resistance protein; PEPT, peptide transporter; OCTN, organic cation/carnitine transporter; P-gp, P-glycoprotein; P450, cytochrome P450; UGT, UDP glucuronosyltransferase.
- Received December 18, 2008.
- Accepted February 11, 2009.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}