


Abstract
Drug-induced cholestasis can result from the inhibition of biliary efflux of bile acids in the liver. Drugs may inhibit the hepatic uptake and/or the biliary efflux of bile acids resulting in an increase in serum concentrations. However, it is the intracellular concentration of bile acids that results in hepatotoxicity, and thus serum concentrations may not necessarily be an appropriate indicator of hepatotoxicity. In this study, sandwich-cultured rat hepatocytes were used as an in vitro model to assess the cholestatic potential of drugs using deuterium-labeled sodium taurocholate (d8-TCA) as a probe for bile acid transport. Eight drugs were tested as putative inhibitors of d8-TCA uptake and efflux. The hepatobiliary disposition of d8-TCA in the absence and presence of drugs was measured by using liquid chromatography/tandem mass spectrometry, and the accumulation (hepatocytes and hepatocytes plus bile), biliary excretion index (BEI), and in vitro biliary clearance (Clbiliary) were reported. Compounds were classified based on inhibition of uptake, efflux, or a combination of both processes. Cyclosporine A and glyburide showed a decrease in total (hepatocytes plus bile) accumulation, an increase in intracellular (hepatocytes only) accumulation, and a decrease in BEI and Clbiliary of d8-TCA, suggesting that efflux was primarily affected. Erythromycin estolate, troglitazone, and bosentan resulted in a decrease in accumulation (total and intracellular), BEI, and Clbiliary of d8-TCA, suggesting that uptake was primarily affected. Determination of a compound's relative effect on bile acid uptake, efflux, and direct determination of alterations in intracellular amounts of bile acids may provide useful mechanistic information on compounds that cause increases in serum bile acids.
Drug-induced liver toxicity is the single most common reason for withdrawal of Food and Drug Administration-approved drugs from the market (Kaplowitz, 2001). Recent data suggest that hepatic transport proteins may be an important site of toxic interactions, and inhibition of the basolateral uptake and/or canalicular excretion of bile acids (cholestasis) by drugs or metabolites is becoming a well recognized cause of liver disease (Lewis and Zimmerman, 1999; Kosters and Karpen, 2008). Basolateral transporters are essential in the hepatic uptake of drugs from the blood, whereas canalicular transporters are involved in the elimination of drugs and bile acids across the canalicular membrane into the bile. Drugs or metabolites that affect these transporter processes can lead to the intracellular accumulation of bile acids resulting in the development of cholestatic liver damage (Fattinger et al., 2001; Funk et al., 2001b).
Transporters involved in the hepatic uptake of drugs and bile acids from the blood belong to the sodium-dependent and sodium-independent pathways. The sodium taurocholate cotransporting polypeptide (NTCP) accounts for the uptake of 80% of conjugated bile acids (i.e., taurocholic and glycocholic acids), and to a much lesser extent it accounts for unconjugated bile acids (Hagenbuch and Dawson, 2004). In addition to NTCP, members of the superfamily of organic anion-transporting polypeptides are involved in the sodium-independent transport of bile acids (Hagenbuch and Meier, 2004). Whereas multiple transporters are involved in the hepatic uptake of bile acids, the bile salt export protein (BSEP) is the primary transporter involved in the biliary efflux of conjugated bile acids across the canalicular membrane including taurocholate, glycocholate, chenodeoxycholate, and deoxycholate (Byrne et al., 2002).
In addition to their involvement in the transport of bile acids and other endogenous substrates, basolateral and canalicular transport systems are also involved in the transport of drugs. Compounds that compete for similar transport pathways may result in an interaction in which one compound inhibits the transport of another. For example, hepatotoxicity associated with troglitazone and bosentan has been attributed to alterations in hepatic bile acid transport through the inhibition of competing transport pathways (Fattinger et al., 2001; Funk et al., 2001a,b). In vivo, bosentan significantly increased serum bile salt levels (Fattinger et al., 2001). Furthermore, in vitro results showed that BSEP-mediated taurocholate transport was inhibited by bosentan, suggesting that bosentan-induced liver injury is mediated in part by inhibition of BSEP resulting in intracellular accumulation of bile salts and liver damage.
Most in vitro transporter assays using suspended hepatocytes, membrane vesicles, or transfected cell lines primarily assess either hepatic uptake or efflux; however, these assays cannot directly evaluate the relative effects of inhibition of hepatic uptake and/or biliary excretion. Because sandwich-cultured hepatocytes [(SCH) B-CLEAR; Qualyst, Inc., Durham, NC] maintain the expression and function of key uptake and efflux transporters relative to in vivo, it is the most relevant system to evaluate and predict the potential of a compound to cause transporter-based liver toxicity. Several reports have described the use of SCH to assess the effect of compounds on the inhibition of bile acid transport, albeit using different methodologies (Kostrubsky et al., 2003, 2006; Kemp et al., 2005; McRae et al., 2006; Marion et al., 2007). For example, Kostrubsky et al. (2006) used SCH to evaluate the potential of drugs to inhibit bile acid transport. However, inhibition of hepatic uptake or biliary efflux could not be distinguished based on the methodology used in their report because biliary efflux may be affected by inhibition of uptake, efflux, or a combination of both processes. In this report, we describe the development and application of an in vitro screen using sandwich-cultured rat hepatocytes (SCRH) to evaluate the potential of test compounds to inhibit the transport of deuterium-labeled taurocholic acid (d8-TCA) and define parameters that may be used to differentiate between effects on hepatic uptake and/or biliary efflux.
Materials and Methods
Materials.
Cyclosporine A, erythromycin estolate, glyburide, nefazodone, salicylate, troglitazone, and troleandomycin (TAO) were purchased from Sigma-Aldrich (St. Louis, MO). Bosentan was obtained from Toronto Research Chemicals (North York, Ontario, Canada). Stock solutions were prepared at 5 and 50 mM in 100% dimethyl sulfoxide (troglitazone was prepared at 1 and 10 mM) and stored at −20°C. The sodium salt of the stable isotope, d8-taurocholic acid (ethanesulfonic acid, 1,1,2,2-tetradeutero-2-[[(3α,5β,7α,12α)-2,2,4,4-tetradeutero-3,7,12-trihydroxy-24-oxocholan-24-yl]amino]-, monosodium salt) (d8-TCA), was synthesized by Martrex, Inc. (Minnetonka, MN). The internal standard, d4-taurocholic acid (d4-TCA), was purchased from Toronto Research Chemicals Inc. High-performance liquid chromatography grade methanol from Thermo Fisher Scientific (Waltham, MA) and fluka mass spectrometry grade ammonium acetate from Sigma-Aldrich were used for sample preparation and analysis.
Isolation, Plating, and Maintenance of Sandwich-Cultured Rat Hepatocytes.
Hepatocytes were isolated from male Wistar rats (250–300 g) by using a two-step, single path, recirculating collagenase perfusion as reported previously (LeCluyse et al., 1996). Cells were suspended at approximately 1 × 106 cells/ml in medium and subsequently added at a volume of approximately 1.5 ml per well to 6-well BIOCOAT plates (BD Biosciences, Bedford, MA). Postplating, (1–3 h) nonadherent cells were removed by aspiration and replaced with fresh plating medium. After 24 h of incubation, cells were overlaid with 2 ml of Matrigel (0.25 mg/ml) (BD Biosciences) solution prepared in culture medium. Culture medium was replaced every 24 h and uptake studies were conducted on day 4 of culture.
Hepatobiliary Disposition of d8-TCA in Sandwich-Cultured Rat Hepatocytes.
Hepatocytes were washed three times with 1 ml of either Hank's balanced salt solution containing calcium (standard buffer) or Hank's balanced salt solution without calcium containing 0.38 g/l EGTA (calcium-free buffer) and incubated with the third wash either in the presence or absence of test compound (5 and 50 μM, except for troglitazone, which was evaluated at 1 and 10 μM) for 10 min at 37°C. Incubation in standard buffer maintains the integrity of the tight junctions, whereas incubation in calcium-free buffer opens the tight junctions. After the initial incubation, the hepatocytes were washed, and d8-TCA (2.5 μM) and test compound were added to the hepatocytes and incubated. After a 10-min coincubation, the hepatocytes were washed and frozen at −80°C for later analysis of d8-TCA. d8-TCA, measured by liquid chromatography/tandem mass spectrometry (LC/MS/MS) analysis as described below, was used to distinguish between the probe and the endogenous taurocholate produced in sandwich-cultured hepatocytes. Average total protein per well was determined from separate plates from the same lot of hepatocytes using a BCA protein assay kit (Thermo Fisher Scientific), and d8-TCA mass (picomole) was normalized to protein content for each well. The amount of d8-TCA excreted into the bile pockets was determined by subtracting the amount of d8-TCA in the lysates from cells exposed to calcium-free buffer (hepatocytes) from the amount of d8-TCA in the lysates from cells exposed to standard buffer (hepatocytes + bile pockets).
Kinetic studies were performed in SCRH using the same protocol as described above in the absence of test compound. The hepatobiliary disposition of d8-TCA was evaluated over a concentration range of 0.1 to 50 μM and an incubation time of 1 to 20 min. Stock concentrations of d8-TCA were prepared such that the final dimethyl sulfoxide concentrations did not exceed 0.15%.
Sample Preparation for LC/MS/MS Analysis.
A volume of 750 μl of lysis solution [70:30 methanol/water (v:v) containing 25 nM d4-TCA (internal standard)] was added to each well of previously frozen 6-well plates containing study samples or standards. Plates were shaken for approximately 15 min, and the cell lysate solution was transferred to a Whatman 96-well Unifilter Plate (Whatman, Florham Park, NJ). Lysate was filtered into a deepwell plate by centrifugation (2000g for 5 min). The sample filtrate was evaporated to dryness, and the samples were reconstituted in 300 μl of sample diluent containing 60% methanol and 40% ammonium acetate (10 mM, native pH) and mixed for 15 min on a plate shaker. The reconstituted samples were transferred to a Whatman 96-well Unifilter Plate and filtered into a Costar 3956 plate (Corning, Corning, NY) by centrifugation (2000g for 5 min) and sealed with a silicone CapMat (Whatman, Florham Park, NJ) before LC/MS/MS analysis.
LC/MS/MS Analysis.
A Shimadzu binary high-performance liquid chromatography system (Columbia, MD) composed of LC-10ADvp pumps, a CTO-10Avp oven, and an SIL-HTc 96-well autosampler were used. The chromatographic column was either a Thermo Fisher Scientific BetaBasic-18 (100 × 1.0 mm, 3 μm) or a Hypersil Gold C18 (100 × 1.0 mm, 3 μm). Column temperature was maintained at 35°C. A mobile phase gradient composed of 0.5 mM ammonium acetate (native pH) and methanol was used at a flow rate of 50 μl/min and a total run time of 10 min. The d8-TCA retention time was approximately 5 min. An injection volume of 10 μl was used. The kinetic studies used an injection volume of 1 μl for the analysis of samples containing 100 to 1000 pmol/well analyte. Tandem mass spectrometry with negative ion electrospray ionization was conducted with a Thermo Electron TSQ Quantum Discovery MAX (Thermo Fisher Scientific) with an Ion Max ESI source. The transitions monitored at unit resolution for d8-TCA and d4-TCA (internal standard) were (precursor m/z > product m/z) 522 > 128 and 518 > 124, respectively.
Data Analysis.
Accumulation was calculated in hepatocytes plus bile (standard buffer) and hepatocytes (calcium-free buffer) by subtracting the amount of d8-TCA (expressed as picomoles per well) from control plates (nonspecific binding) from the amount of d8-TCA (expressed as picomoles per well) and dividing by protein content (expressed as milligrams of protein per well). The biliary excretion index (BEI) was calculated according to the following equation:

The in vitro biliary clearance (Clbiliary) was determined using the following equation and was scaled to body weight using 0.2 g protein/g liver weight and 40 g liver/kg body weight (Seglen, 1976):

The accumulation (standard and calcium-free buffer), BEI, and in vitro biliary clearance for d8-TCA were expressed as a percentage of the control value (no test compound). Statistical analysis was performed using one-way analysis of variance and Dunnett's multiple comparison test. A P value ≤0.05 was considered significant.
Results
Kinetics of d8-TCA Uptake and Efflux in SCRH.
The effect of substrate concentration and incubation time on the hepatobiliary disposition of d8-TCA was determined in SCRH, and the results are presented in Fig. 1, A and B. The results from time-dependent accumulation of d8-TCA in hepatocytes or hepatocytes plus bile pockets are presented in Fig. 1A. A concentration of 2.5 μM d8-TCA was chosen because it represented a concentration within a linear range of accumulation. The accumulation of d8-TCA in hepatocytes plus bile pockets was linear over the 20-min time period evaluated.

Time (A) (d8-TCA at 2.5 μM) and concentration-dependent (B) (10-min incubation time) accumulation of d8-TCA in SCRH in incubations treated with standard buffer (hepatocytes plus bile) and calcium-free buffer (hepatocytes only).
An increase in d8-TCA concentration resulted in an increase in intracellular accumulation (Fig. 1B), i.e., accumulation in calcium-free buffer representing the mass of d8-TCA accumulated in hepatocytes only. Saturable uptake of d8-TCA was observed within the concentration range evaluated. A concentration-dependent increase in d8-TCA accumulation was observed in standard buffer incubations representing the mass of d8-TCA accumulated in hepatocytes plus bile pockets (Fig. 1B). Accumulation of d8-TCA in hepatocytes plus bile pockets reached a maximum value at 25 μM d8-TCA. Kinetic constants (Km and Vmax) were not determined because accumulation in hepatocytes or hepatocytes plus bile represents several processes including the following: uptake into hepatocytes, efflux across the basolateral membrane, and efflux across the canalicular membrane into bile.
The Effect of Test Compounds on the Hepatobiliary Disposition of d8-TCA in SCRH.
The effects of test compounds (all 50 μM, with the exception of troglitazone at 10 μM) on the accumulation, BEI, and Clbiliary of d8-TCA were evaluated in SCRH, and the results are presented as percentage of control (no test compound) in Figs. 2 and 3.

The effect of test compounds (50 μM) on the accumulation of d8-TCA (2.5 μM) in SCRH in incubations treated with standard buffer (hepatocytes plus bile) and calcium-free buffer (hepatocytes only). Data are expressed as mean ± S.E. (n = 3) of percentage of control (no test compound). ∗, indicates a statistically significant difference from control, P ≤ 0.05.
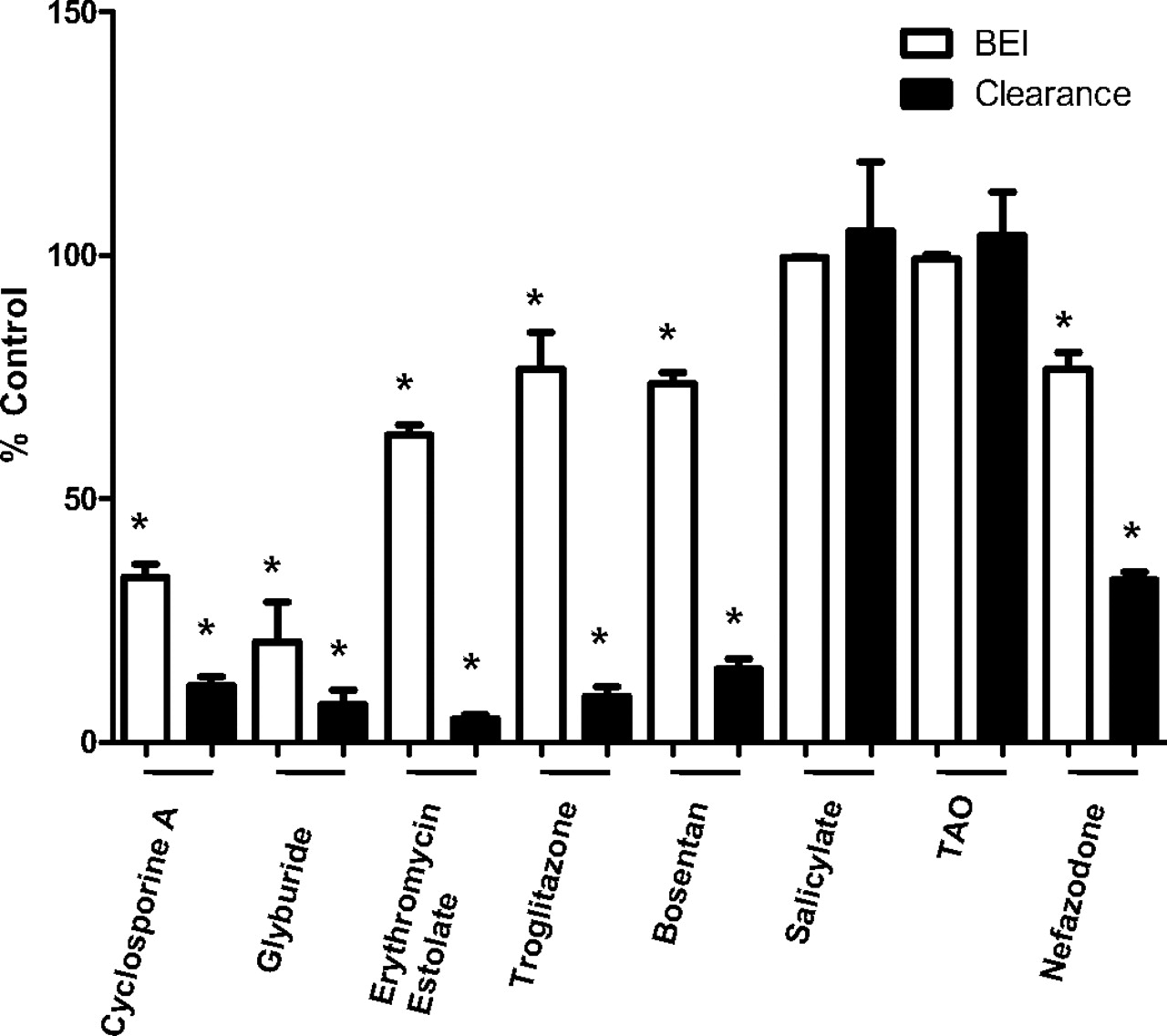
The effect of test compounds (50 μM) on the BEI and Clbiliary of d8-TCA (2.5 μM) in SCRH. Data are expressed as mean ± S.E. (n = 3) of control (no Test Compound). ∗, statistically significant difference from control, P ≤ 0.05.
In the absence of test compound, the accumulation of d8-TCA in incubations with standard buffer (hepatocytes and bile pockets) and calcium-free buffer (hepatocytes only) was 131 ± 31.4 and 15.1 ± 2.47 pmol/mg protein (mean ± S.E.), respectively. Cyclosporine A and glyburide significantly decreased the accumulation of d8-TCA in standard buffer incubations; however, both compounds showed a significant increase in the accumulation of d8-TCA in calcium-free buffer incubations as presented in Fig. 2. Erythromycin estolate, troglitazone, and bosentan significantly decreased the accumulation of d8-TCA in both standard and calcium-free buffer incubations. Nefazodone significantly decreased the accumulation of d8-TCA in standard buffer incubations and had no effect on the accumulation of d8-TCA in calcium-free buffer incubations. Salicylate and troleandomycin had no effect on the accumulation of d8-TCA in either incubation.
The effects of test compounds on the biliary efflux of d8-TCA were measured and presented as BEI and Clbiliary (Fig. 3). In the absence of test compound, the BEI for d8-TCA was 88.3 ± 1.32%, indicating that 88% of the d8-TCA taken up by the hepatocytes was effluxed into the bile. Of the eight compounds evaluated, glyburide and cyclosporine A had the greatest inhibitory effect on the BEI (Fig. 3), reducing the BEI of d8-TCA to 21 and 34% of control, respectively. Erythromycin estolate, nefazodone, bosentan, and troglitazone showed an approximately 20 to 35% inhibitory effect on the BEI of d8-TCA. The decreases in BEI were all statistically significant, except for salicylate and TAO, which had no effect on the BEI of d8-TCA.
The Clbiliary of d8-TCA in the absence of inhibitor was 37.1 ± 9.54 ml/min/kg. All test compounds evaluated, with the exception of salicylate and TAO, showed a statistically significant decrease in the in vitro biliary clearance of d8-TCA.
Discussion
Cholestasis can be defined as any condition in which substances normally excreted into bile are retained. The most common method for the clinical determination of cholestasis is to measure serum concentrations of bile acids or conjugated bilirubin. Bile acids are strong detergents that cause cell membrane injury and impairment of membrane function. However, an increase in serum bile acid concentrations may not necessarily reflect an increase in intracellular hepatocyte concentrations. It is generally assumed that it is the concentration of bile acids inside the hepatocyte that is the primary determinant of hepatotoxicity. Thus, it is important to differentiate between a compound's effect on the uptake or efflux of bile acids, because inhibition of uptake will result in decreased hepatocellular concentrations of bile acids that would be less likely to cause hepatotoxicity, whereas, inhibition of efflux (either basolateral and/or canalicular) would result in increased hepatocyte concentrations of bile acids, increasing the potential for hepatotoxicity. Inhibition at either site will result in increased serum concentration of bile acids, and therefore increased serum levels may not necessarily reflect the hepatotoxic potential of a drug.
Transporter-based drug interactions that inhibit basolateral uptake or efflux (basolateral or canalicular) in the liver may lead to the alteration of the in vivo hepatobiliary disposition of bile acids (Fattinger et al., 2001; Funk et al., 2001a,b; Pauli-Magnus and Meier, 2006). Knowledge of the site of inhibition is important in understanding the relationship between elevated serum concentrations of bile acids and the intracellular concentration of bile acids in hepatocytes leading to hepatotoxicity. Unlike other in vitro transporter models, SCH can simultaneously determine the potential for a compound to alter the basolateral uptake and/or the basolateral or canalicular efflux of bile acids and therefore predict the overall effect a drug may have on bile acid disposition in vivo (Kemp et al., 2005). Inhibition of basolateral efflux of bile acids could result in an increase in the intracellular concentration of bile acids. If the increased levels of bile acids are within the linear range of the canalicular efflux transporters, the BEI should not change; however, the in vitro biliary clearance could increase under this condition.
Treatment with cyclosporine A, glyburide, erythromycin estolate, troglitazone, and bosentan decreased the in vitro biliary clearance of d8-TCA to less than 20% of the control value. The in vitro biliary clearance is an indicator of the overall effect of the compound on bile acid excretion. A decrease in the in vitro biliary clearance reflects a decrease in the amount of d8-TCA excreted into the bile, which can result from inhibition of either of the following: 1) basolateral uptake transporters or 2) canalicular efflux transporters. The BEI represents the fraction of the total mass of d8-TCA taken up that is excreted into the bile. A decrease in the BEI represents inhibition of d8-TCA efflux into the bile. The BEI, in conjunction with the accumulation and in vitro biliary clearance, can be used to determine the site of action (basolateral/uptake versus canalicular/efflux) of a particular compound on the hepatobiliary disposition of bile acids in SCH.
Cyclosporine A and glyburide resulted in a decrease in the accumulation of d8-TCA in samples treated with standard buffer, representing a decrease in the total mass of d8-TCA accumulated in hepatocytes plus bile pockets. Both compounds also decreased the BEI of taurocholate to 34 and 21% of control, suggesting an inhibition of bile acid efflux out of the hepatocyte. Both compounds increased the mass of d8-TCA in calcium-free treated samples (representing an increase in hepatocellular concentration). This activity suggests that canalicular (and/or basolateral) efflux processes were inhibited to a greater extent than uptake processes, resulting in an increase in the hepatocellular concentration of bile acids. The potential for cyclosporine A and glyburide to increase the levels of bile acids inside of the hepatocytes may be related to their potential for in vivo hepatotoxicity. These observations are consistent with previous reports showing increased bile acid levels in the livers of rats treated with cyclosporine A and glyburide resulting in cholestatic hepatotoxicity (Chan and Shaffer, 1997; Mizuta et al., 1999; Kostrubsky et al., 2003). Hepatotoxicity was also reported in human subjects treated with cyclosporine A, resulting in a 2- to 3-fold increase in total serum bile acids (Kassianides et al., 1990; Tripodi et al., 2002), which is consistent with the in vitro results obtained in this study using SCRH. However, cyclosporine A and glyburide are not associated with high incidences of hepatotoxicity in vivo, and only a few cases of cholestatic hepatotoxicity were reported with the use of these drugs. The low incidences of hepatotoxicity associated with these drugs could be explained by the low in vivo plasma concentrations of these drugs (0.2 μM glyburide and 8.3–332 nM cyclosporine A) or differential intracellular accumulation in human hepatocytes due to differences in kinetic properties of uptake and efflux transporters between rat and human hepatocytes.
Erythromycin estolate, an erythromycin analog used in the treatment of bacterial infections, has been associated with cholestatic liver injury. In SCRH, erythromycin estolate decreased d8-TCA accumulation by approximately 70 and 90% in hepatocytes and hepatocytes plus bile, respectively. This effect resulted in a decrease in the Clbiliary of d8-TCA to less than 5% of control. However, the BEI decreased to only 63% of control, suggesting that erythromycin estolate has more of an effect on the uptake of d8-TCA into the hepatocyte than on the efflux of d8-TCA into the bile. Bosentan and troglitazone demonstrated similar effects on the hepatobiliary disposition of d8-TCA in SCRH as observed for erythromycin estolate. The Clbiliary of d8-TCA decreased to approximately 15% of control for bosentan and troglitazone. d8-TCA accumulation decreased by approximately 50 and 70% in hepatocytes and hepatocytes plus bile, respectively, whereas the BEI decreased by less than 25%, suggesting that troglitazone and bosentan have a potent effect on uptake of d8-TCA in addition to their inhibition of the efflux of d8-TCA into bile. These results are consistent with the findings of Leslie et al. (2007) in which bosentan was identified as a potent inhibitor of rat Ntcp. Furthermore, Kemp et al. (2005) also observed inhibition of hepatic uptake and biliary efflux of TCA by troglitazone coadministration in SCRH. At the concentrations evaluated in our experiments, troglitazone had a greater effect on hepatic uptake than on biliary efflux because accumulation was inhibited to a greater extent than the BEI. In vivo studies in rats demonstrated that troglitazone and its metabolites inhibited the efflux of bile acids by interfering with the canalicular efflux transporter, Bsep (Funk et al., 2001a,b).
Nefazodone had no effect on the accumulation of d8-TCA in hepatocytes; however, coadministration with nefazodone resulted in an approximately 50% decrease in accumulation in hepatocytes plus bile (consistent with an effect on uptake). The lack of an effect on the accumulation of d8-TCA in hepatocytes along with the decrease in the accumulation in hepatocytes plus bile and the decrease in the BEI indicates that nefazodone is inhibiting both uptake and efflux of d8-TCA. If inhibition were strictly on the efflux processes, the accumulation in hepatocytes would be greater than that of control as seen for cyclosporine A and glyburide. However, accumulation in hepatocytes was similar to control values; therefore, to decrease the mass excreted in bile, both hepatic uptake and efflux must be inhibited to some extent. Nefazodone has been reported to increase serum bile acids in rat (Kostrubsky et al., 2006) and was withdrawn from the market due to hepatotoxicity (Spigset et al., 2003). Salicylate and troleandomycin were used as negative controls to demonstrate that compounds with no reported in vivo cholestatic potential have no effect on uptake and/or efflux of d8-TCA in SCRH.
The observations of increased hepatocellular amounts of bile acids in the presence of cyclosporine A and glyburide indicate that it is important to use methodology that can simultaneously evaluate the effect of inhibitor compounds on the uptake and efflux of bile acids. Determination of the hepatocellular amount of bile acids allows for the differentiation between compounds that cause a decrease in the in vitro biliary clearance, resulting from inhibition of uptake and/or efflux of bile acids, and those that cause a decrease in the in vitro biliary clearance by strongly inhibiting the efflux of bile acids into the bile. Methodologies that evaluate only efflux cannot quantitate the role of uptake transporter inhibition relative to the inhibition of bile acid efflux (Kostrubsky et al., 2006). B-CLEAR sandwich-cultured hepatocytes have also demonstrated the capacity to synthesize endogenous bile acids. After 4 days of culture, taurocholic acid, glycocholic acid, and the taurine and glycine conjugates of chenodeoxycholic acid, and muricholic acid have been detected inside the hepatocytes by using liquid chromatography/mass spectrometry analysis. The effect of drugs on the production and hepatobiliary disposition of endogenous bile acids may offer additional insights to the hepatotoxic effects of drugs, and this experimentation is an area of ongoing research in our laboratories.
In summary, SCH may serve as an in vitro model to assess the cholestatic potential of drug candidates. The methodology used in this study allows for the simultaneous assessment of both hepatic uptake and biliary efflux and assessment of alterations in the hepatocellular amounts of bile acids. The site of inhibition may be an important parameter in understanding whether increased serum bile acids in vivo may lead to cholestatic hepatotoxicity (inhibition of efflux leading to an increase in intracellular bile acids). Furthermore, a deuterium-labeled taurocholate analog (d8-TCA) may serve as a useful probe for assessing hepatobiliary disposition of bile acids in SCH, eliminating the need for use of radiolabeled probes.
Footnotes
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
doi:10.1124/dmd.109.028407.
-
- NTCP
- sodium taurocholate cotransporting polypeptide
- BSEP
- bile salt export protein
- SCH
- sandwich-cultured hepatocytes
- TAO
- troleandomycin
- d8-TCA
- deuterium-labeled taurocholic acid
- LC/MS/MS
- liquid chromatography/tandem mass spectrometry
- BEI
- biliary excretion index
- Clbiliary
- in vitro biliary clearance
- SCRH
- sandwich-cultured rat hepatocytes.
- Received June 17, 2009.
- Accepted November 4, 2009.
- Copyright © 2010 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}