


Abstract
Newborn rats are more sensitive to the toxic effects of cardiac glycosides than are adult rats. This is associated with a decreased ability to remove cardiac glycosides from blood into the liver. Pregnenolone-16α-carbonitrile (PCN), a prototypical rodent CYP3A inducer and pregnane-X-receptor (PXR) ligand, stimulates the hepatic clearance of cardiac glycosides in newborn rats, which results in decreased toxicity of the cardiac glycosides. The mechanism responsible for this phenomenon is not clear; however, if elucidated, it would help in understanding and preventing potential drug-drug interactions. The recently cloned rat organic anion-transporting polypeptide 2 (oatp2) (Slc21a5) is a sinusoidal hepatic uptake transporter, with very high affinities for cardiac glycosides, and thus it was hypothesized that rat oatp2 increases during postnatal development and is inducible by PCN. In the present study, livers were removed from Sprague-Dawley rats from postnatal days (pnd) 0 to 45, in 5-day increments; as well as from pnd 10 to 90, in 10-day increments, after PCN (75 mg/kg i.p., for 4 days) or corn oil (vehicle for PCN) treatment. The protein and mRNA levels of rat oatp2 were determined by Western blot analysis and branched DNA signal amplification technique, respectively. Expression of rat oatp2 protein and mRNA increased gradually during postnatal development. PCN treatment increased liver to body weight ratio in both genders, and dramatically accelerated the maturation of hepatic oatp2 protein and mRNA levels. In summary, rat oatp2 undergoes age-dependent and chemical regulation during postnatal development, and is a potential target for drug-drug and age-drug interactions.
Compared with adult rats, newborn rats are more sensitive to the toxicity elicited by cardiac glycosides, such as ouabain and digoxin (Klaassen, 1972, 1973). This is associated with the decreased ability of the liver in newborn rats to remove cardiac glycosides from the blood, thereby causing greater toxicity in newborn rats (Klaassen, 1972). The mechanism of toxicity has been further characterized by showing that the rodent CYP3A inducer pregnenolone-16α-carbonitrile (PCN2) increases hepatic clearance of cardiac glycosides in newborns, thus reducing cardiac glycoside toxicity (Klaassen, 1974a,b). The kinetics of ouabain uptake in isolated hepatocytes from rats at various ages demonstrates thatVmax increases with age, suggesting an increase in uptake transporter(s). Although there is no statistical difference in the Km of ouabain uptake in hepatocytes during development, theKm tended to be higher in the young rats, suggesting the involvement of two transporting systems (Stacey and Klaassen, 1979).
Further understanding of the hepatic uptake of cardiac glycosides was made possible by the recent advances in the cloning of the rat organic anion-transporting polypeptide 2 (oatp2) (Slc21a5) (Noe et al., 1997). Rat oatp2 is expressed in the hepatic sinusoidal membrane (Noe et al., 1997; Abe et al., 1998, 1999; Gao et al., 1999), in the choroid plexus (Reichel et al., 1999), and in the retina (Noe et al., 1997; Abe et al., 1998, 1999; Gao et al., 1999; Reichel et al., 1999). Oatp2 transports structurally unrelated compounds, such as anions, cations, and neutral compounds, in a sodium- and ATP-independent manner (Meier et al., 1997; Noe et al., 1997; Abe et al., 1998; Li et al., 1998;Eckhardt et al., 1999; Reichel et al., 1999). Furthermore, oatp2 transports cardiac glycosides such as ouabain and digoxin with very high affinity (Noe et al., 1997). Although the first cloned oatp family member, organic anion-transporting polypeptide 1 (oatp1), also transports ouabain with low affinity, it does not appear to account for the hepatic uptake of ouabain (Sasabe et al., 1997; Kouzuki et al., 1999). Therefore, rat oatp2 appears to be the main transporter responsible for cardiac glycoside uptake into adult liver. Although rat oatp2 appears to be important for cardiac glycoside uptake into liver, its regulation during development and after chemical treatment during postnatal development has not been studied.
In the present study, the expression of rat oatp2 protein and mRNA was determined in rat liver during postnatal development in both male and female rats. The effect of PCN treatment on the regulation of hepatic oatp2 was also determined during postnatal development in both genders. The results demonstrated that rat oatp2 increases steadily at both the protein and mRNA levels during postnatal development. PCN treatment greatly enhances the rat oatp2 protein and mRNA expression at all postnatal ages. The data explain, in part, the reason why newborn rats are more sensitive to cardiac glycoside toxicity and how PCN protects newborn rats from cardiac glycoside toxicity. Furthermore, the results indicate potential drug-drug and age-drug interactions due to differences in expression of liver sinusoidal transporters responsible for hepatic uptake of xenobiotics.
Experimental Procedures
Materials.
Pregnenolone-16α-carbonitrile was purchased from Sigma Chemical (St. Louis, MO). Bicinchoninic acid protein assay kit was obtained from Pierce Chemical (Rockford, IL). Primary polyclonal antibody for rat oatp2 was developed in our laboratory against a synthetic peptide designed from a unique region of rat oatp2 protein (Rausch-Derra et al., 2001). Horseradish peroxidase-conjugated secondary antibody, protein molecular weight marker, and chemiluminescence reagents for Western blot analysis were from Amersham Biosciences, Inc. (Piscataway, NJ). SDS-polyacrylamide gel electrophoresis reagents were from Novex (San Diego, CA). QuantiGene high volume kit for the direct quantification of cellular mRNA was purchased from Bayer Diagnostics (East Walpole, MA). Other chemicals were from Sigma Chemical, unless otherwise indicated.
Animals and Treatments.
Female and male Sprague-Dawley (SD) rats were obtained from Sasco Laboratories, Inc. (Kingston, NY). For age groups equal to or younger than 50 days old, the offspring of pregnant female rats bred at the University of Kansas Medical Center facilities were used. The day of birth of the newborn rats was considered postnatal day (pnd) 0. For age groups older than 50 days, SD rats were ordered from the supplier. The animals were housed according to American Association for the Accreditation of Laboratory Animal Care guidelines and were fed with Harlan Teklad rat chow (Harlan Bioproducts for Science, Indianapolis, IN) and water ad libitum. Newborn rats were weaned at pnd 21. PCN was administered i.p. in corn oil at 75 mg/kg/day for four consecutive days. Control rats were treated with the same volume of corn oil (5 ml/kg) and the same schedule as the PCN-treated rats. Twenty-four hours after the last administration, livers were removed and snap-frozen in liquid nitrogen, and stored at −80°C until used.
Preparation of Membrane Protein Samples.
Liver membrane protein samples were prepared according to Trauner et al. (1997). Briefly, liver membrane protein samples were prepared by homogenizing liver samples (0.1–0.5 g) in 10 ml of buffer A (0.25 M sucrose; 10 mM Tris-HCl, pH 7.5; 25 μg/ml leupeptin; 50 μg/ml aprotinin and antipain; 0.5 μg/ml pepstatin; and 40 μg/ml phenylmethylsulfonyl fluoride), by using a Teflon pestle and a 15-ml glass homogenizing vessel (Wheaton, Millville, NJ). The homogenate was centrifuged at 4°C at 100,000g for 1 h. The pellet was then resuspended in buffer B (0.3 mM sucrose and 20 mM HEPES, pH 7.5, with 40 μg/ml phenylmethylsulfonyl fluoride). The protein concentration of each sample was determined by the bicinchoninic acid procedure by using a bicinchoninic acid kit (Pierce Chemical).
Western Blot Analysis.
Polyacrylamide gel electrophoresis (running buffer: 25 mM Tris, 192 mM glycine, 0.1% SDS, pH 8.4) was performed to separate membrane proteins and molecular markers (Bio-Rad, Hercules, CA) by using 12% Tris-glycine gels from Novex. Two universal internal standards (two samples) were used in each gel to correct for gel-to-gel differences. Proteins in the gels were transferred onto polyvinylidene difluoride membranes (Novex) for 3 h at 20 V (transfer buffer: 12 mM Tris base, 96 mM glycine, 20% methanol). Membranes were placed in a blocking solution [5% nonfat dry milk in Tris-buffered saline/Tween 20 (TBST)] overnight at 4°C. Blots were then incubated with primary oatp2 antibody (1:1000 in TBST with 2.5% nonfat dry milk) for 3 h at room temperature. After thorough washing, blots were incubated with secondary antibody (1:5000 in TBST) (donkey anti-rabbit IgG, Amersham Biosciences, Inc.) for 1 h at room temperature. Blots were then washed thoroughly, and antibody was detected by enhanced chemiluminescence (Amersham Biosciences, Inc.). Blots were exposed to radiographic film (X-MOAT AR; Eastman Kodak, Rochester, NY) for 1 min. The film was developed and examined by densitometric analysis by using a densitometer (Molecular Dynamics, Sunnyvale, CA), followed by quantification with ImageQuant software, version 4.2a (Molecular Dynamics).
Total RNA Isolation.
Total RNA was isolated using RNAzol B reagent (Tel Test B, Friendswood, TX) according to the instructions of the manufacturer, and the integrity of the total RNA was assessed by agarose gel electrophoresis. RNA concentration was determined by UV absorbance at 260 nm.
Rat oatp2 Probe Set.
The specific oligonucleotide probe sets (consisting of capture extenders, label extenders, and blockers) were designed using the Bayer probe Designer software, version 1.0 (Emeryville, CA), where each oligonucleotide probe has a Tm of 63°C. Every probe was submitted to the National Center for Biotechnological Information for nucleotide comparison by the basic logarithmic alignment tool (BLASTn). All probes were synthesized by Operon Technologies (Palo Alto, CA). The compete list of rat oatp2 oligonucleotide probes has been previously published (Rausch-Derra et al., 2001).
mRNA Assay.
Rat oatp2 mRNA levels were assessed using the Quantigene expression kit as described in the manufacturer's protocol, and validated in this laboratory (Hartley and Klaassen, 2000). Briefly, the oligonucleotide probes (capture extenders, label extenders, and blockers) were combined and diluted to 50 fmol/ml in lysis buffer (100 mM HEPES buffer, pH 7.6; 0.65 mg/ml proteinase K; 1% lithium lauryl sulfate; 800 mM lithium chloride; 8 mM EDTA; and 0.5% Micro-O-protect). Total RNA (1 μg/μl, 10 μl) was added to each well containing 50 μl of capture hybridization buffer (100 mM HEPES buffer, pH 7.6; 3 mg/ml Boehringer-Mannheim blocking reagent; 1% lithium lauryl sulfate; 8 mM EDTA; and 0.5% Micro-O-protect) and 50 μl of each probe set in lysis buffer. RNA was allowed to hybridize to each probe set overnight at 53°C. Subsequently, the plate was allowed to cool to 46°C, and washed with washing buffer [2× with 0.1× standard saline citrate (1× standard saline citrate contains 0.15 M sodium chloride, 0.015 M sodium citrate), 1% sodium dodecyl sulfate, and 0.5 mg/ml sodium azide]. Samples were hybridized with the branched DNA (bDNA) amplification molecule (50 μl/well at 0.2 fmol/μl bDNA) in the amplifier/label probe buffer (100 mM HEPES buffer, pH 7.6; 1.5 mg/ml Boehringer-Mannheim blocking reagent; 1% lithium lauryl sulfate; 10 μM ZnCl2; 1 mM MgCl2; 1% Brij 35) for 30 min at 53°C. The plate was allowed to cool to room temperature, and the wells were washed with washing buffer (two times). The alkaline phosphatase conjugated label probe was diluted in amplifier/label probe buffer, added to each well (50 ml/well at 0.4 fmol/μl alkaline phosphatase), and allowed to hybridize to the bDNA-RNA complex for 15 min at 53°C. The plate was cooled to room temperature and washed with washing buffer (2 times). The reaction was triggered by the addition of a dioxetane substrate solution (50 μl/well) of Lumi-Phos Plus (Lumigen, Inc., Southfield, MI) containing 0.3% SDS. The enzymatic reaction was allowed to proceed for 1 h at 37°C, and the luminescence was measured with the Quantiplex 320 Luminometer (Chiron Corp., Emeryville, CA) equipped to read 96-well plates.
Statistical Analysis.
The data were expressed as mean ± S.E. Statistical significance was determined by one-way analysis of variance, followed by Duncan's post hoc test. The level of significance was set at p< 0.05.
Results
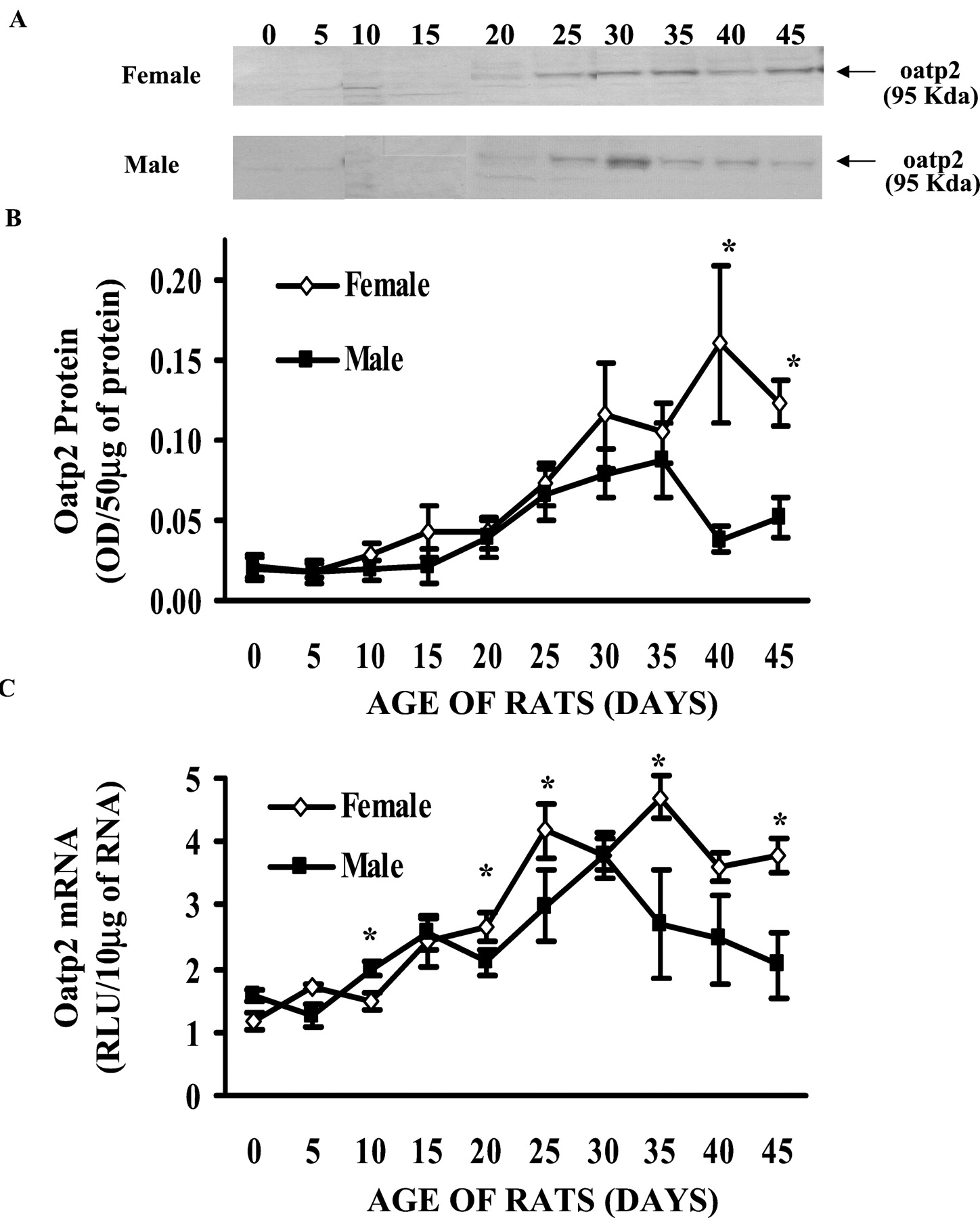
Normal Postnatal Development of oatp2 Protein in Male and Female Rats.
Figure 1, A and B, illustrates the normal postnatal development of rat oatp2 protein, determined by Western blot analysis, in male and female SD rats, from pnd 0 to 45, in 5-day increments. The expression level of oatp2 protein was relatively low at birth in both genders. In males, pnd 0 rats had approximately 35% of the concentration of oatp2 protein in liver as in adult liver. There was no apparent increase of oatp2 protein from pnd 0 to 15, but levels increased gradually from pnd 20, reaching peak levels around pnd 35, and then declined to adult levels by pnd 40 to 45. In females, oatp2 protein at pnd 0 was about 20% of that in adult females. The oatp2 protein in females started to increase gradually from pnd 0 to 40, when it reached maximal expression. Although oatp2 protein in females appeared to be slightly higher than that in males, it was only statistically significant at pnd 40 and 45.

Postnatal development of oatp2 protein and mRNA in male and female rat.
Livers were removed from 0- to 45-day-old male (▪) and female (⋄) rats, at 5-day intervals. Each group was composed of four to five animals. For protein determination (A), membrane proteins were isolated and separated on 12% SDS-polyacrylamide gel electrophoresis (50 μg of protein/lane). Oatp2 protein was detected by a polyclonal antibody designed against a unique peptide region of oatp2 protein. B, quantitative expression of oatp2 protein. For mRNA determination (C), total RNA was used to detect oatp2 mRNA levels by the bDNA signal amplification technique, with oatp2 gene-specific probe sets. The data are expressed as mean ± S.E. (n = 4–5). Asterisks (∗) denote statistical significance between male and female rat values (p < 0.05).
Normal Postnatal Development of oatp2 mRNA in Male and Female Rats.
The mRNA levels of rat oatp2 during postnatal development were determined by the bDNA technique. Figure 1C illustrates the normal postnatal ontogeny of rat oatp2 mRNA expression in male and female rats from pnd 0 to 45, in 5-day increments. Overall, the mRNA expression of oatp2 in both genders was similar to that of protein, starting with lower expression at birth, followed by a gradual increase from pnd 0 to 35, when maximal expression of oatp2 mRNA was reached in both genders. Oatp2 mRNA expression then decreased slightly after pnd 35, reaching a plateau at pnd 40 and 45. The expression of oatp2 mRNA also appeared to be higher in female than male rats, with the differences being statistically significant at pnd 20, 25, 35, and 45.
Effects of PCN Treatment on Liver to Body Weight Ratio during Postnatal Development.
This result is illustrated in Fig. 2. In corn oil-treated control groups, the liver to body weight (L/B WT) ratio in both genders tended to increase slowly from pnd 10 to 40, and then declined slowly over time. At pnd 90, the L/B WT ratio was similar to that at pnd 10. Although the L/B WT ratio in PCN-treated rats generally followed the same trend as in the corn oil control groups, PCN treatment significantly increased L/B WT ratio in both genders during postnatal development. In male rats, PCN significantly increased L/B WT in all age groups, except pnd 40 when the increase was not statistically significant (Fig. 2A). In female rats, although there was no apparent increase of L/B WT ratio by PCN at pnd 10 and 20, PCN significantly increased L/B WT ratio from pnd 30 to 90 (Fig. 2B).

Liver to body weight ratio in rats treated with PCN during postnatal development.
Male and female rats were treated with PCN (75 mg/kg) or vehicle (corn oil) for 4 days by i.p. injection. Each corn oil-treated group had four animals, and each PCN-treated group had five animals. Livers were removed, corresponding to 10, 20, 30, 40, 50, 60, and 90 days of age. Comparison of liver to body weight between control and PCN-treated male rats is shown in A, whereas comparison of liver to body weight between control and PCN-treated female rats is shown in B. Corn oil-treated rats are represented with open diamonds, and PCN-treated rats are represented with filled squares. All data are expressed as mean ± S.E. for four to five animals in each group. Asterisks (∗) denote statistical significance from control values (p < 0.05).
Effect of PCN Treatment on Rat oatp2 Protein Expression during Postnatal Development.
Treatment with PCN, a prototypical rodent CYP3A inducer and pregnane-X-receptor (PXR) ligand, dramatically increased the expression of oatp2 protein in both male (Fig. 3A) and female rats (Fig. 3B) throughout postnatal development. The induction of oatp2 protein in both male and female rats was so dramatic with PCN treatment that the concentration of oatp2 protein in 10-day-old rats, in which oatp2 protein is undetectable in control rats, was even higher than that in 90-day-old control adult rats. PCN induced oatp2 significantly in both genders; however, female rats tended to respond to PCN more than male rats. In addition, induction of oatp2 protein by PCN in both genders at pnd 20 seemed to be higher compared with any other age group.

Effects of PCN on oatp2 protein expression during postnatal ontogeny.
Male and female rats were treated i.p. for 4 days with PCN (75 mg/kg) or vehicle (corn oil). Livers were removed at 10, 20, 30, 40, 50, 60, and 90 days of age. A and C, representative Western blots of oatp2 protein after PCN treatment in male and female rats, respectively, during postnatal development. Oatp2 protein was detected by the polyclonal antibody designed against a unique peptide region of oatp2 protein. The gray columns represent the corn oil-treated control rats, and the filled columns represent the PCN-treated rats. B and D, summarized quantitative results from the Western blots (n = 4–5). The data are expressed as mean ± S.E. Asterisks (∗) denote statistical significance between corn oil- and PCN-treated rats (p < 0.05).
Effect of PCN Treatment on Rat oatp2 mRNA Expression during Postnatal Development.
Because oatp2 protein was dramatically induced by PCN treatment, it was of interest to determine whether increased levels of oatp2 mRNA were responsible for the increase in protein. Therefore, oatp2 mRNA levels were determined after PCN treatment in rats from pnd 10 to 90, in 10-day increments. PCN increased oatp2 mRNA significantly at all age groups in both genders (Fig. 4, A and B), except in 30-day-old male rats in which PCN did not induce oatp2 mRNA. The induction was very dramatic in that the oatp2 mRNA concentrations in 10-day old animals were close to adult concentrations. Furthermore, female rats also appeared to respond more to PCN induction than male rats, similar with what was seen at the protein level. In addition, pnd 20 male rats tended to be induced by PCN more than any other age group. However, pnd 20 female rats did not show the marked increase in oatp2 mRNA as demonstrated in male rats.

Effects of PCN on oatp2 mRNA expression during postnatal ontogeny.
Male and female rats were treated i.p. for 4 days with PCN (75 mg/kg) or vehicle (corn oil). Livers were removed at 10, 20, 30, 40, 50, 60, and 90 days of age. A and B, effects of PCN on male and female rat oatp2 mRNA expression, respectively, during postnatal ontogeny. Oatp2 mRNA was determined by the bDNA signal amplification technique, with gene-specific probe sets for oatp2. Each group was composed of four to five animals. The gray columns represent the corn oil-treated control rats, and the filled columns represent the PCN-treated rats. The data was expressed as mean ± S.E. Asterisks (∗) denote statistical significance between corn oil- and PCN-treated rats (p < 0.05).
Discussion
Oatp2 belongs to the multispecific organic anion-transporting polypeptide family, and was the second member cloned in rats (Noe et al., 1997). Although there are overlaps with regard to tissue distribution and substrates among the oatp family members, oatp2 is unique, due to its high-affinity and specific transport of cardiac glycosides, such as digoxin and ouabain (Noe et al., 1997; Abe et al., 1998). This laboratory previously showed that newborn rats are much more sensitive to cardiac glycoside toxicity compared with adult rats, and this appears to be due to insufficient hepatic uptake of cardiac glycosides in the younger animals (Klaassen, 1972). Some microsomal enzyme inducers, such as PCN and phenobarbital, were able to decrease the cardiac glycoside toxicity in younger animals by increasing the hepatic uptake and excretion of cardiac glycosides (Klaassen, 1972,1973; Stacey and Klaassen, 1979). The mechanism underlying this phenomenon has not been elucidated; however, determining the mechanism by which this protection occurs would aid in the understanding and possible prevention of drug-drug interactions. The recent molecular cloning and functional characterizations of rat oatp2 provide a potential candidate for the aforementioned phenomenon, making an in-depth investigation possible at the molecular level.
The present study demonstrated that the expression of rat oatp2 is regulated during postnatal development. The expression of oatp2 protein and mRNA was very low at birth, it increased gradually with age, reaching peak levels at about pnd 30 to 35, then slowly declined to adult levels. In addition, livers of female rats seemed to have a slightly higher expression of oatp2 than male rats. It appears that the oatp family members follow a similar temporal pattern with regard to the developmental regulation of their mRNAs (Dubuisson et al., 1996). The ontogenic study with rat oatp1 mRNA has demonstrated that there is also a time-related expression of oatp1 in liver (Dubuisson et al., 1996). Oatp1 mRNA expression is first detected on day 16 of gestation, remaining stable during the perinatal period and increasing dramatically after weaning (Dubuisson et al., 1996). The increase of both oatp1 and oatp2 expression after weaning might be attributed to the dramatic increase in bile acid concentrations in serum at this age (Belknap et al., 1981; Suchy et al., 1981; Shneider et al., 1993,1995). The postnatal development of Na+-dependent taurocholate-cotransporting polypeptide, the Na+-dependent hepatic bile acid uptake transporter, seems to occur earlier than the development of oatp1 and oatp2. The Na+-dependent taurocholate-cotransporting polypeptide protein and mRNA were also low at birth in newborn rats compared with adult animals; however, by 1 week of age, they have reached adult levels (Shneider et al., 1995).
The present data indicate that female rats tend to express more oatp2 protein and mRNA than male rats, especially when the animals were older than 30 to 35 days of age. Because it is known that rats become sexually mature around pnd 30 to 35, increased oatp2 expression in female rats at this age might be attributed to the increased estrogen levels in the circulation. However, whether estrogen is a positive regulator for oatp2 needs further investigation.
Throughout postnatal development, oatp2 protein and mRNA were both induced dramatically by PCN, the prototypical inducer for rodent CYP3A (Pichard et al., 1991; Michalets, 1998) and ligand for PXR (Bertilsson et al., 1998; Kliewer et al., 1998; Lehmann et al., 1998; Jones et al., 2000). The PXR has recently been shown to be essential for the induction of CYP3A and the human P-glycoprotein transporter that is encoded by the gene MDR1 (Geick et al., 2001). In the paradigm in which PCN induces rodent CYP3A, PXR, which resides in the nucleus, heterodimerizes with the retinoid-X-receptor α. Activation of gene transcription by the PXR-retinoid-X-receptor α heterodimer is prevented by the interaction with one or more inhibitory transcriptional factors unless a PXR ligand, such as PCN, binds to PXR, which results in the release of nuclear corepressors and recruitment of nuclear coactivators, such as SRC-1, to initiate PXR-regulated gene expression (Sueyoshi and Negishi, 2001). The strong induction of oatp2 by PCN indicates that oatp2 probably is a PXR target gene. One could speculate that different phases of drug metabolism are under coordinate regulation, i.e., chemicals that induce drug biotransformation enzymes (such as CYP3A) would also induce the drug uptake transporters (such as oatp2) and/or the drug export transporters (such as P-glycoprotein).
The degree of induction of oatp2 protein by PCN in both male and female pnd 20 rats seems higher than any other age groups. Rats are weaned at pnd 21 at which time the body undergoes dramatic physiological changes due to the switch from high-lipid, low-carbohydrate-containing milk to high-carbohydrate, low-lipid-containing animal chow. As a result of this switch, the insulin-to-glucagon ratio increases (Mayor and Cuezva, 1985; Ferre et al., 1986; Girard et al., 1991). In addition, thyroxin and corticosterone levels are higher than those of suckling rats (Henning, 1981). These or other endogenous factors might interact with PCN to generate the marked induction of oatp2 in 20-day-old rats. As mentioned earlier, oatp1 mRNA is also dramatically increased after weaning, which might be induced by increased bile acid concentrations after weaning (Belknap et al., 1981; Suchy et al., 1981; Shneider et al., 1993, 1995). Interestingly, at pnd 20, only oatp2 mRNA in male rats was induced by PCN to the comparable levels of protein; oatp2 mRNA in female rats did not respond as robustly as in male rats, indicating probably a different regulation of oatp2 protein in females around weaning.
In summary, the present studies characterized the postnatal development of oatp2 protein and mRNA in rat liver. Oatp2 undergoes a time-dependent increase during postnatal ontogeny. PCN, a prototypical rodent CYP3A inducer and PXR ligand, dramatically increases oatp2 protein and mRNA during postnatal development. These results suggest that rat neonates may be more sensitive to cardiac glycoside toxicity because they have low levels of oatp2 to transport cardiac glycosides into the liver. The present study also indicates that PCN may prevent cardiac glycoside toxicity in newborn rats by dramatically increasing the amount of oatp2. Furthermore, the study indicates that hepatic uptake is a potential target for drug-drug and age-drug interactions.
Acknowledgments
We acknowledge Dr. Nathan J. Cherrington for excellent guidance on the bDNA signal amplification technique.
Footnotes
-
↵1 Present address: Department of Biochemistry and Molecular Pharmacology, University of Massachusetts Medical School, 55 Lake Ave. North, Worcester, MA 01655-0126.
-
This work was supported by National Institute of Environmental Health Sciences Grants ES-09649 and ES-03192. D.R.J. was supported by the National Institute of Environmental Health Sciences Training Grant ES-07079.
- Abbreviations used are::
- PCN
- pregnenolone-16α-carbonitrile
- oatp2
- organic anion-transporting polypeptide 2
- oatp1
- organic anion-transporting polypeptide 1
- SD
- Sprague-Dawley
- pnd
- postnatal day
- TBST
- Tris-buffered saline/Tween 20
- bDNA
- branched DNA signal amplification technique
- L/B WT
- liver to body weight
- PXR
- pregnane-X-receptor
- Received August 9, 2001.
- Accepted December 6, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}