


Abstract
The transepithelial intestinal elimination of ivermectin was studied using the intestinal closed-loop model in the rat. The common bile duct was cannulated, and duodenum, jejunum, and ileum were isolated in situ with their intact blood supplies. Following administration of 100, 200, or 400 μg/kg b.wt. ivermectin via the carotid artery, the elimination of parent ivermectin into the small intestinal lumen over 90 min was approximately 5-fold higher than in bile. The major amount of secreted ivermectin was recovered in the jejunum, but the duodenum showed a higher intestinal elimination capacity than the other intestinal segments with respect to the intestinal length. Systemic coadministration of the P-glycoprotein blocker verapamil significantly reduced the elimination capacity of jejunum by 50%, which resulted in a 30% decrease of ivermectin overall elimination by the small intestine. In contrast, verapamil did not significantly affect ivermectin secretion in duodenum, ileum, or bile in the same animals. Ivermectin small intestinal and biliary clearances were estimated to account for 27 and 5.5% of the total drug clearance, which was evaluated from a parallel in vivo experiment in which rats were given 200 μg/kg b.wt. ivermectin intra-arterially. In conclusion, intestinal secretion plays a greater role than biliary secretion in the overall elimination of ivermectin in the rat, providing major amounts of active drug to the intestinal lumen and to feces. This is discussed in terms of therapeutic efficacy against intestinal parasites in humans and animals and of ecotoxicity resulting from the contamination of livestock dung with parent drug.
Ivermectin is an anthelmintic of unprecedented efficacy, currently used worldwide to treat and control various parasitic diseases in human and veterinary medicine. It has become the drug of choice in the treatment of a major public disease, human onchocerciasis, and is administered to more than 18 million people each year (Burkhart, 2000). Ivermectin is extensively eliminated by the fecal route as parent drug and metabolites, regardless of the species and of the route of administration, with less than 2% excreted in the urine (Campbell, 1985). As the biliary concentration of ivermectin is substantially higher than that in plasma (Bogan and McKellar, 1988; Lifschitz et al., 2000), it has been assumed that biliary secretion was the major pathway of elimination of the parent drug. The study of Hennessy et al. (2000), performed in sheep with the structural analog doramectin, supported this hypothesis. However, absolute amounts of biliary secreted ivermectin have never been determined in human or animal species. In addition, recent findings have shown that ivermectin was a substrate for some intestinal efflux transporters, which points toward the possible existence of another elimination pathway, e.g., the intestinal secretion of drug from blood into the intestinal lumen.
P-gp1 is a plasma membrane protein, which is able to pump a broad range of structurally and functionally unrelated compounds out of the cell in an energy-dependent manner (reviewed bySharom, 1997). First identified as a factor for multidrug resistance in mammalian tumor cells (Juliano and Ling, 1976), P-gp has been discovered to be physiologically expressed in a number of tissues including the liver, intestines, and blood-brain barrier (Thiebaut et al., 1987; Cordon-Cardo et al., 1989). The strategic distribution of P-gp on the biliary canalicular membrane of hepatocytes and on the apical side of enterocytes provides a mechanistic support for both the biliary and the intestinal secretion of xenobiotics (Hunter and Hirst, 1997; Smit et al., 1998; Van Asperen et al., 1998).
Ivermectin was shown to be actively excreted in vitro by multidrug-resistant tumor cells (Pouliot et al., 1997) and by cells transfected by the gene coding for P-gp in human or in mouse,MDR1 and mdr1a, respectively (Schinkel et al., 1995; Smith et al., 2000). In vivo, the kinetic disposition of ivermectin was modified in mice lacking their P-gp I function (Schinkel et al., 1994; Lankas et al., 1997; Kwei et al., 1999), with a markedly increased accumulation in brain tissue and signs of neurotoxicity. In the intestines, P-gps constitute a barrier against the absorption of orally administered ivermectin (Kwei et al., 1999), but their role in ivermectin elimination was never documented.
The purpose of the current study was to investigate in vivo the existence, and the extent, of a nonbiliary intestinal elimination pathway with direct secretion of ivermectin from systemic blood through the gut wall. The involvement of P-gp in ivermectin elimination was assessed by coadministration of verapamil, a prototypic P-gp substrate and inhibitor (Ford and Hait, 1990), which was already shown to affect the in vivo disposition of ivermectin in the rat following topical application (Alvinerie et al., 1999).
Experimental Procedures
Materials.
Ivermectin (IVOMEC; injectable ovin) was purchased from Merial (Lyon, France). Verapamil hydrochloride was obtained from Sigma-Aldrich (St. Quentin Fallavier, France), and [14C]inulin (62.5 μCi/ml) was obtained from PerkinElmer Life Sciences (Boston, MA). All other chemicals used were of the highest grade available.
Animals.
Male Wistar rats (Iffa Credo, L'Arbresle, France) weighing 323 ± 30 g were used in the experiments. All the investigations were performed in accordance with the European regulations for the use of laboratory animals.
Pilot Experiments (Open-Perfusion Model).
Two rats were prepared as described below for the closed-loop model (Trial 1), but the intestinal elimination was investigated from a 20-cm jejunal loop only, following systemic administration with 200 or 400 μg/kg b.wt. ivermectin. The jejunal segment was isolated 2 cm distal to the ligament of Treitz and perfused continuously with thermostated saline at the constant flow rate of 0.6 ml/min determined as optimal bySavina et al. (1981). A catheter was placed at the distal end of the segment, allowing serial recuperation of intestinal perfusates into tubes at 8-min intervals, up to 184 min. The rats were sacrificed at the end of the experiments. The 20-cm segment was excised, opened lengthwise, and the residual content was scraped. Acetonitrile was added to the perfusate samples to achieve an acetonitrile/water 4:5 v/v mixture, and all samples were stored at −20°C until analysis.
Trial 1 (Intestinal Closed-Loop Model).
After an overnight fast with free access to water, animals were anesthetized by i.p. administration of 1.5 g/kg b.wt. urethane and 40 mg/kg b.wt. α-chloralose. Rats were tracheotomized for a facilitated respiration under anesthesia and placed under a heating lamp. The left carotid artery was cannulated with a polyethylene tube (i.d., 0.58 mm) and perfused using an electric syringe with thermostated (37°C) saline at the constant flow rate of 1 ml/h. A midline abdominal incision was made, and a polyethylene tube (i.d., 0.3 mm) was inserted into the common bile duct, near the hilum of the liver, for bile collection. The segments of the small intestine were exposed in situ with their blood supply intact. A polyethylene catheter (25 × 0.9 mm; BD Biosciences, Meylan, France) was placed at the proximal end of the duodeno-jejunum (starting from the pylorus and ending 30 cm above the ileocecal junction), and the intestinal contents were washed out by perfusing thermostated (37°C) saline for 20 min at the constant rate of 0.6 ml/min. A small incision was made at the distal end of the duodeno-jejunum to allow collection of intestinal contents and effluents. Five minutes before the end of intestinal perfusion, the segment was gently ligated at its distal end and filled with perfusion liquid. The proximal end of the segment was ligated, and one additional ligature was placed 1 cm distal to the ligament of Treitz to separate the duodenum from the jejunum. The two segments were carefully returned to the abdominal cavity. The ileal segment (30 cm) was prepared following the same procedure. Care was taken not to interrupt the normal blood supply of the segments during their preparation. The abdominal cavity was then covered with a compress soaked with saline to prevent heat loss and evaporation. The perfusion of the carotid artery with saline was interrupted, and ivermectin was injected rapidly (1 min) via the catheter at the dose of 100, 200, or 400 μg/kg b.wt. (ivermectin was diluted in polyethylene glycol 400 to fit low dose rates). Saline was perfused again during 90 min.
In the competition experiments, the i.a. perfusion of saline was stopped 5 min prior to ivermectin administration, and verapamil was given at a loading rate of 24 mg/kg b.wt. per hour during 5 min via the carotid artery. Ivermectin was then injected i.a. at the standard dose of 200 μg/kg b.wt., and verapamil was perfused again at 4.8 mg/kg b.wt. per hour and during 90 min.
Bile was collected throughout the experiment. Animals were sacrificed at 90 min after ivermectin administration. Blood was previously collected by cardiac puncture in some treated and nontreated rats. The intestinal segments were excised, their length measured, and they were opened lengthwise. The whole intestinal content was carefully collected using a cell scraper, and 3 ml of acetonitrile was added to the samples. The scraping procedure was very gentle to remove the mucus layer with minimal disruption of the intestinal cells. Blood samples were centrifuged, and plasma was collected. Biliary, plasma and intestinal samples were stored at −20°C until analysis. Five to six rats were used to test each dose level of ivermectin in the noncompetition experiments, and seven rats were used in the competition experiments.
Another group of animals (n = 8) was taken for an accurate estimation of the area under the plasma concentration-time curve between 0 and 90 min postadministration. Rats were anesthetized as described above, but no other surgical procedure was performed. Blood was sampled at 2, 5, 15, 30, 60, and 90 min after i.a. administration of 200 μg/kg b.wt. ivermectin.
Validation of the Study Model of Intestinal Elimination.
To determine the extent of nonspecific paracellular transport, three rats were prepared as described for the closed-loop model. They were injected with 9 μCi of [14C]inulin via the carotid artery. At 90 min postadministration, the intestinal content was collected from duodenum, jejunum, and ileum, and radioactivity was measured in a beta scintillation counter (Kontron Beta V, Montigny Le Bretonneux, France).
Trial 2: Determination of Ivermectin Total (Plasma) Clearance.
Forty-two rats were used to evaluate ivermectin total clearance. Rapid anesthesia was achieved by i.p. injection of 87 mg/kg b.wt. xylazine and 13 mg/kg b.wt. ketamine. The left carotid artery was cannulated, and ivermectin was injected via the catheter at the dose of 200 μg/kg b.wt.. The rats began to recover 30 min after the beginning of anesthesia. At 2, 5, 15, 30, 60, and 90 min, 4, 8, and 24 h, and 2, 3, 4, 5, 7, 9, 12, 15, and 18 days postadministration, two to three rats were taken for terminal blood sampling performed by cardiac puncture under anesthesia (xylazine and ketamine, i.p.).
Analytical Assay.
Ivermectin was analyzed using a high-performance liquid chromatography method with automated solid phase extraction and fluorescence detection, as previously described by Alvinerie et al. (1987). Minor modifications were made to fit analysis of low plasma and bile volumes (extraction of 100 μl of plasma or bile).
The intestinal samples were sonicated twice for 20 min. After 2 min of centrifugation at 2000 g, the clean supernatant was transferred into a first tube, and the pellet was resuspended in 3 ml of acetonitrile, sonicated for 20 min, and centrifuged (2000g, 2 min). The resulting clean supernatant was removed into a second tube. Deionized water was added to the tubes to achieve an acetonitrile/water 4:5 v/v mixture. Solid phase extraction was then performed as described by Alvinerie et al. (1987).
Ivermectin H2B1a (22,23-dihydroavermectin B1a) was designated as analyte. The limit of quantification of the high-performance liquid chromatography method was 0.05 ng/ml. Accuracy and precision (intra-assay variation) expressed as relative standard deviation were less than 8 and 6%, respectively.
Pharmacokinetic and Statistical Analysis.
The plasma concentrations were fitted using nonlinear regression analysis (SYSTAT 8.0; SPSS Inc., Chicago, IL). The areas under the mean plasma concentration-time curve, AUC (0-tlast) (from 0 to the last quantifiable sample) and AUC (0–90) (from 0 to 90 min), were calculated using the trapezoidal rule. Ivermectin total clearance was obtained by dividing the administered dose (200 μg/kg) by AUC (0-tlast). Biliary and intestinal clearances were computed as the ratio of the total amount of drug (H2B1a) eliminated within 90 min into bile and in the small intestinal lumen, respectively, divided by the corresponding plasma AUC (0–90) obtained in Trial 1. The mean residence time (MRT), the plasma terminal half-life (t1/2λz), and the steady-state volume of distribution (Vss) were calculated according to the classical pharmacokinetic equations associated with noncompartmental analysis (Gibaldi and Perrier, 1982).
The results were expressed as means ± S.D. Analysis of variance was used to test the significance of differences between the three intestinal segments and the three dose levels (SYSTAT 8.0). Post hoc comparisons were performed using the Bonferroni test. A ttest was done to compare verapamil-treated and nontreated rats. Ap < 0.05 was regarded as significant.
Results
Validation of the Study Model.
Less than 0.1% of the administered radioactivity was detected in any of the intestinal segments of rats receiving labeled inulin, indicating that the integrity of intestinal epithelium was preserved under the described experimental conditions.
Pilot Experiments (Open-Perfusion Model).
The perfusion of the 20-cm jejunal segment with saline resulted in low basal concentrations of parent ivermectin in most perfusate samples. However, very high concentrations of parent ivermectin were observed in intestinal effluents each time some intestinal mucus had leaked accidentally via the distal catheter into the sample. These results indicate that ivermectin was largely associated with mucus. Following systemic administration of 200 and 400 μg/kg ivermectin, 111 and 163 ng of ivermectin were eliminated unchanged from the 20-cm loop, respectively. About 65 and 74% of these amounts were obtained from perfusate samples, and 35 and 26% by collection of residual mucus in the lumen of the loop.
Trial 1 (Closed-Loop Model).
At the end of the 90-min experiments, the contents of the closed intestinal loops consisted almost exclusively of mucus containing high amounts of ivermectin as parent drug. The total amount of ivermectin recovered in the small intestinal lumen exceeded systematically (from 3.2 to 8.8 times) that eliminated in bile over the same 90-min period in the same animal (Table 1). The ratio of intestinal to biliary elimination did not show any significant difference between the tested doses (Table 1). The cumulative amount of ivermectin excreted into bile and in the small intestinal lumen within 90 min following i.a. administration of ivermectin accounted for approximately 1% of the administered dose, independently of the dose level. The bile flow rate was homogenous among the rats and equal to 0.84 ± 0.13 ml/h.
Total amounts (ng/kg b.wt.) of ivermectin eliminated into bile and in the small intestinal lumen over 90 min postadministration in rat, and biliary and intestinal clearances
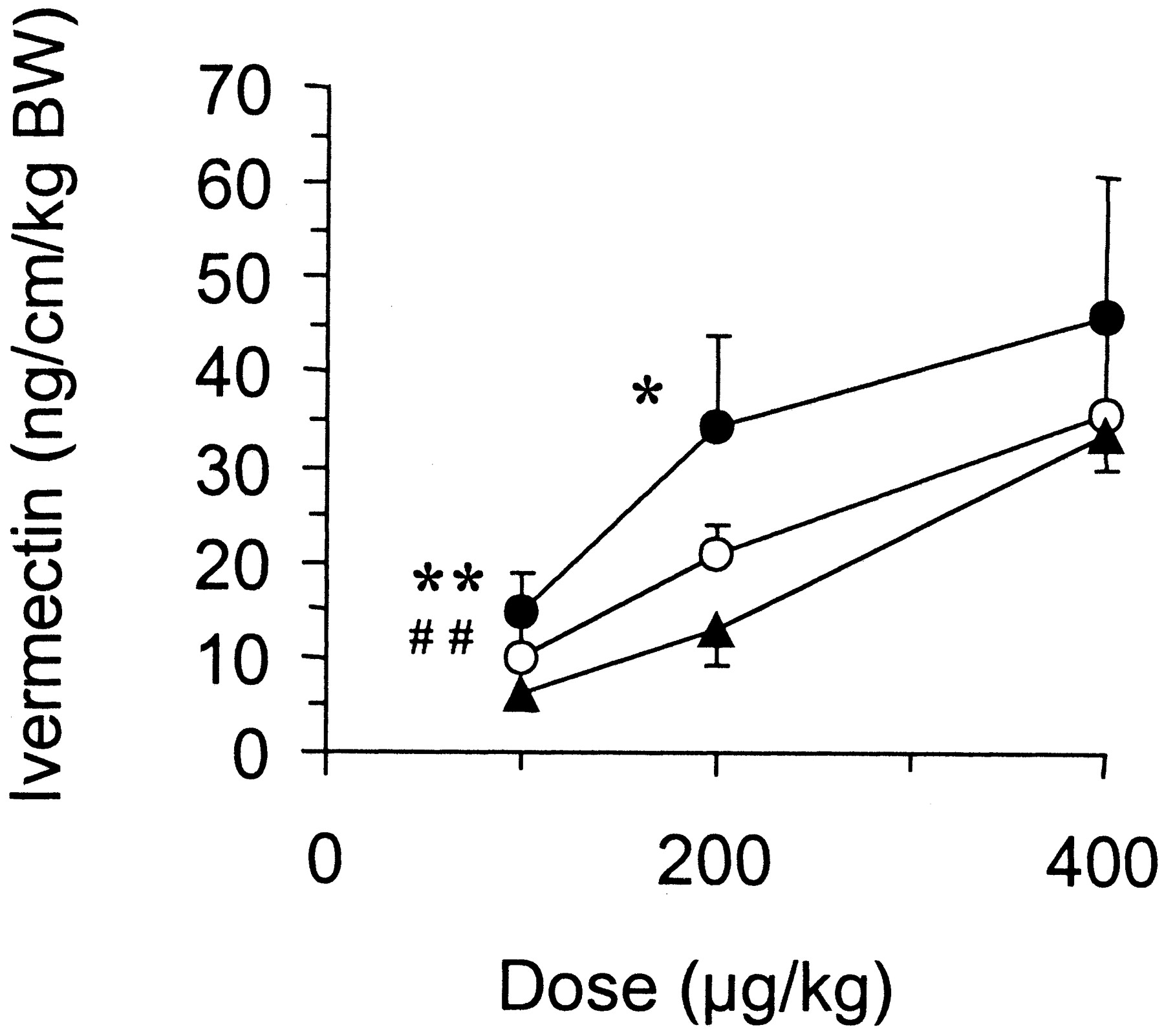
Over the dose range tested, the intestinal elimination of ivermectin was greater in the jejunum than in the other intestinal segments (Table1). However, when normalized by the length of the segment, ivermectin elimination was statistically higher in the duodenum at the doses of 100 and 200 μg/kg and statistically lower in the ileum at the dose of 100 μg/kg, in the same animal (Fig. 1).

Segmental elimination of ivermectin through the gut wall following intra-arterial administration to rats at the doses of 100 (n = 6), 200 (n = 5), and 400 μg/kg b.wt. ivermectin (n = 5).
The data (mean ± S.D.) correspond to the elimination capacities of the intestinal segments, which were obtained by dividing the total amount of parent drug eliminated in duodenum, jejunum, or ileum over 90 min by the length of the corresponding segment. ★ (p < 0.05) and ★★ (p < 0.01) indicate a significant difference between duodenum and jejunum in the same animal, whereas ## (p < 0.01) indicates a significant difference between jejunum and ileum. ●, duodenum; ○, jejunum; ▴, ileum.
Coadministration of the P-gp inhibitor, verapamil, significantly reduced the overall small intestinal elimination of ivermectin by 28% (1397 ± 378 versus 1936 ± 204 ng/kg in controls,p < 0.05) but did not statistically affect ivermectin secretion into bile (346 ± 129 versus 389 ± 111 ng/kg in controls). The elimination capacity of the jejunum was markedly reduced by 50% in the treated rats compared with the controls, whereas no statistical difference could be observed with respect to the rate of elimination in the duodenum or in the ileum (Fig.2). Plasma concentrations of ivermectin at 90 min postadministration were not significantly different between treated rats (82 ± 9.6 ng/ml) and nontreated rats (71 ± 9.6 ng/ml). No signs of toxicity were detected in any of the seven rats treated with verapamil under our experimental conditions.

Influence of verapamil on the elimination capacities of the intestinal segments in rats given 200 μg/kg i.a. ivermectin.
The data (mean ± S.D.) correspond to the total amounts of parent drug eliminated in each intestinal segment over 90 min postadministration in treated (n = 7) and nontreated rats (n = 5), divided by the length of the segment. ★★, p < 0.01.
The second group of rats taken for an accurate estimation of AUC (0–90) showed plasma levels of ivermectin very similar to those obtained in the rats given 200 μg/kg ivermectin in the closed-loop study. The AUC (0–90) was equal to 168 ng · h/ml. Small intestinal and biliary clearances in situ were 0.277 and 0.056 l/day/kg, respectively.
Trial 2.
The plasma data were best fitted with a triexponential concentration-time curve (Fig. 3). The values of AUC (0-tlast) and AUC (0–90) were 2233 and 355 ng · h/ml, respectively. The MRT,t1/2 λz, andVss were 1.09 days, 2.51 days, and 2.34 l/kg, respectively. Ivermectin total clearance was found to be equal to 2.15 l/day/kg. Trial 2 was referred to as an in vivo situation, considering that xylazine and ketamine produced a short-duration surgical anesthesia with moderate side effects.

Ivermectin plasma concentration-time profile following intra-arterial administration of rats (n = 42) with 200 μg/kg ivermectin.
Each point corresponds to one rat.
The difference between the AUCs (0–90) in Trial 2 and that in Trial 1 (twice lower) shows that anesthesia with urethane and α-chloralose, albeit required by the model, most likely influenced the plasma disposition of ivermectin. This was taken into account for the assessment of ivermectin total (plasma) clearance in the rats used in Trial 1. Assuming the homogeneity of the rats used in Trials 1 and 2, the total plasma clearance of ivermectin in Trial 1 (in situ) was estimated from that obtained in Trial 2 (in vivo) corrected by the ratio of the AUCs (0–90), e.g., 1.02 l/day/kg. It results that the small intestinal clearance of ivermectin accounted for 27% of the corrected total (plasma) clearance versus 5.5% for the biliary clearance.
Discussion
At all three different ivermectin dose rates, the amount of parent ivermectin eliminated in the small intestinal lumen was 5 times higher than in bile. Since conjugates of parent ivermectin have never been detected in bile, this study clearly demonstrates that the major route for the elimination of parent ivermectin in the rat intestine is not biliary but intestinal secretion. Such a difference between intestinal and biliary elimination has been already observed for other drugs such as roxithromycin (Arimori et al., 1998), ciprofloxacin (Dautrey et al., 1999), and vinblastine (Van Asperen et al., 2000).
The open-perfusion model is commonly used to study the intestinal secretion of drugs as it enables measurement of an overall intestinal elimination by keeping the reabsorption phenomenon negligible (Dautrey et al., 1999). The use of this model was, however, not desirable in the present study given the extensive binding of ivermectin to intestinal mucus. Indeed, the random leakage of mucus in perfusate samples observed in pilot experiments impairs a reliable and reproducible quantification of the drug intestinal elimination. Consequently, we used the alternative closed-loop model, of which the advantage was to extend the investigation from 20-cm loops (open-perfusion model) to the totality of the small intestine, allowing thereby a more reliable prediction of the in vivo state. The resulting drawback was, however, a possible reabsorption into blood of the ivermectin secreted in the intestinal lumen and thus the measurement of the net intestinal elimination of drug compared with the overall biliary secretion. It is therefore possible that the intestine-to-bile elimination ratio might be slightly underestimated.
On the other hand, it cannot be excluded that the gentle scraping procedure for collection of intestinal contents resulted in a slight overestimation of ivermectin intestinal elimination. This procedure was necessary to ensure collection of the totality of the mucus present in the lumen as it is known to be partly bound to the glycocalyx layer of enterocytes. However, this technique implies that some intestinal mucosa tissue might be scraped as well. In pilot experiments (open-perfusion model), up to 74% of the drug eliminated by a 20-cm loop was already obtained from perfusate samples and not by scraping. Thus, even if the contribution of intestinal mucosa tissue to the total collection of ivermectin cannot be excluded, this would not affect the conclusions of the study. The experiments were terminated 90 min after ivermectin administration, before a deterioration of animal condition and depletion of bile salts could have been able to affect the bile flow and thus the extent of ivermectin elimination in bile.
Compared with the other intestinal segments, the jejunum provided the major source of parent ivermectin regardless of the administered dose. However, the intestinal elimination capacity (expressed per unit of intestinal length) appeared to decrease gradually from proximal-to-distal small intestine, with maximal values in the duodenum. Regional differences could possibly be attributed to differences in passive diffusion, expression levels of efflux transporters, or carrier affinity (Makhey et al., 1998), but other factors should be considered as well in interpretation of data such as a different proportion of villi in the intestinal segments (Olivier et al., 1998). Data were expressed per unit of intestinal length and not of intestinal area, since the loop diameter along the segments was too variable to be properly assessed.
In duodenum and jejunum, increasing the dose from 200 to 400 μg/kg did not result in a significantly higher absolute elimination of ivermectin, which is consistent with the involvement of an active (saturable) transport mechanism. As the P-gp substrate and inhibitor verapamil significantly inhibited the elimination of ivermectin into the jejunum by 50%, the involvement of a P-gp carrier-mediated process in the jejunum can be assumed. In contrast, neither the i.a. perfusion of verapamil nor an i.a. bolus administration of 5 mg/kg verapamil (data not shown) could significantly affect ivermectin elimination in bile, duodenum, or ileum in the same animals. It is likely that, at the applied low (nontoxic) dosages, verapamil did not completely block the P-gp function. However, these results also suggest that other efflux mechanisms might play a significant role in the secretion process. This would be in agreement with the observations of Kwei et al. (1999) who systemically administered 200 μg/kg b.wt. ivermectin to mdr1a-P-gp-deficient mice with a noncannulated gallbladder and reported only a partial 30% reduction (from 3.4 to 2.4% of the administered dose) of the cumulative ivermectin excreted in bile plus intestinal contents compared with wild-type mice. Additional in vitro or ex vivo studies should clarify the potential role of P-gp and/or other carriers without the drawbacks related to the use of P-gp inhibitors in vivo and highlight the differences between intestinal segments.
Finally, qualitative analysis of chromatograms indicated the ability of the intestinal epithelium to excrete ivermectin-derived metabolites (data not shown). As for the parent drug, this should be further investigated in vitro.
Biliary and small intestinal clearances of ivermectin were found to account for 5.5 and 27% of ivermectin total (plasma) clearance, respectively. Intestinal clearance was calculated solely from the small intestine, but other regions of the digestive tract may participate as well in the elimination process. Ivermectin elimination by distal parts of the digestive tract has never been investigated thus far, but colon has been reported to display high levels of P-gp expression (Thiebaut et al., 1987) and therefore could provide a significant contribution to the absolute intestinal elimination of ivermectin as for other xenobiotics (Mayer et al., 1996; Ramon et al., 1996; Makhey et al., 1998; Van Asperen et al., 2000). Besides these considerations, our findings suggest an important metabolism of ivermectin in rat. The proportion of metabolites in fecal drug residues has never been documented in rat following systemic administration but is known to represent approximately 60 and 70% in cattle and swine, respectively (Halley et al., 1989), which would be consistent with our results.
Our study suggests the existence of an intestinal route of elimination of ivermectin in human and in target animal species. The observations reported in cattle by Bogan and McKellar (1988) support this hypothesis. Following subcutaneous administration, ivermectin was found at rather high concentrations in small intestinal mucus with no significant difference between mucus distal and proximal to the bile duct opening. Furthermore, ileal fluid showed three times higher concentrations of ivermectin than in intestinal fluids sampled proximal to the bile duct. In contrast, following systemic administration with the structural analog doramectin to sheep, Hennessy et al. (2000)recovered 130% of the administered dose in bile. This would imply that bile was the major elimination pathway, which is difficult to conciliate with our results apart from interspecies differences.
The characterization of an intestinal elimination pathway in human and target animal species would be of therapeutic significance. Not all parasites feed on plasma, and the compartmental distribution of drugs out of the plasma in secondary compartments (bile, intestinal secretions) needs to be taken into consideration to discuss and optimize the efficacy of antiparasitic drugs. It has been suggested that ivermectin could be available to parasites in intestinal mucus (Bogan and McKellar, 1988). Consequently, an extensive elimination of ivermectin along the digestive tract following systemic absorption or parenteral administration would provide high concentrations of drug at the site of action. It is also possible that the mucus functions as a reservoir for exchange of drug into intestinal fluid.
Besides clinical considerations, it is noteworthy that a large part of the ivermectin excreted in cattle dung (38% of the dose after systemic administration; Laffont et al., 2001) may arise from intestinal secretion. The presence of parent (active) ivermectin in feces can have deleterious effects on nontarget organisms such as some dung-degrading and dung-breeding insects (Wall and Strong, 1987; Sommer et al., 1993). Although the issue of environmental impact of ivermectin used in large scale in cattle is still under debate, our study provides new directions into the pharmacokinetic behavior of endectocides to reduce the fecal excretion of parent drug. The therapeutically desirable intestinal secretion should be then evaluated against the undesirable exposure of the environment with feces containing ivermectin.
In conclusion, we have provided evidence that the major route for the elimination of parent ivermectin in the rat intestine is not biliary but intestinal secretion, which enforces the role of the intestines in the elimination of xenobiotics. In vitro studies are now underway to clarify the mechanisms involved in ivermectin intestinal elimination. A better understanding of these mechanisms is of interest for the further development of endectocides to optimize their safety and efficacy.
Acknowledgments
We thank Pr. Johanna Fink-Gremmels for carefully reading the manuscript and fruitful discussion.
Footnotes
- Abbreviations used are::
- P-gp(s)
- P-glycoprotein(s)
- AUC
- area under the curve
- Received October 25, 2001.
- Accepted February 13, 2002.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}