


Abstract
Understanding the role of transporters in placental handling of xenobiotics across the maternal-fetal interface is essential to evaluate the pharmacological and toxicological potential of therapeutic agents, drugs of abuse, and other xenobiotics to which the mother is exposed during pregnancy. Therefore, the purpose of this study was to assess mRNA levels of various transporters in placenta and to compare these to levels in maternal liver and kidney, predominant organs of excretion, to determine which transporters are likely to have a role in xenobiotic transfer within the placenta. During late stage pregnancy, relative amounts of mRNA levels of 40 genes representing 11 families/group of transporters were assessed in placenta with respect to relative maternal liver and kidney mRNA levels. Members of the following transporter families were assessed: three multidrug resistance (Mdr), six multidrug resistance-associated protein (Mrp), eight organic anion-transporting polypeptide (Oatp), three organic anion transporters (Oat), five organic cation transporters (Oct), two bile acid transporters (Na+/taurocholate-cotransporting polypeptide and bile salt export protein), four metal (ZnT1, divalent metal transporter 1, Menkes and Wilsons), a prostaglandin, two peptide, two sterolin, and four nucleoside transporters. Of the 40 genes evaluated, 16 [Mdr1a and 1b, Mrp1 and 5, Oct3 and Octn1, Oatp3 and 12, four metal, a prostaglandin, AbcG8, equilibrative nucleoside transporter 1 (ENT1), and ENT2] were expressed in placenta at concentrations similar to or higher than in maternal liver and kidney. The abundance of these mRNA transcripts in placenta suggests a role for these transporters in placental transport of xenobiotics and supports their role in the transport of endogenous substances.
Transporters within the placenta play a crucial role in the distribution of nutrients across the maternal-fetal interface. There are transporters that perform vital physiological functions in facilitating the transfer of nutrients and other normal metabolites across the placenta, such as amino acids, nucleosides, monocarboxylates, dicarboxylates, vitamins, and folate. Many of these transporters recognize xenobiotics as substrates, due to structural resemblance to physiological substrates, and mediate the transfer of these xenobiotics across the placenta. There are also transporters in the placenta that appear to function exclusively as xenobiotic transporters (Ganapathy et al., 2000).
In general, xenobiotic transporters mediate the passage of water-soluble xenobiotics into the cell and the excretion of these xenobiotics and/or their metabolites out of cells. Thus, the presence or absence of these transporters is important in determining whether a xenobiotic will accumulate in a tissue. Furthermore, localization of each transporter may aid in the prediction of toxicity due to xenobiotic exposures. In embryo/fetal development, this concept is of extreme importance. The placenta is the sole link between mother and the developing fetus and performs a multitude of functions that are essential for the maintenance of pregnancy and normal development of the fetus. Moreover, the placenta was once thought to serve as a physical barrier that provided absolute protection to the developing fetus. However, research over the years following the thalidomide debacle has proven the theory of placenta as a physical barrier to be a falsehood. In fact, the placenta, which is derived from both fetal and maternal tissue, is considered the first fetal organ to be exposed to exogenous substances and is now viewed as a metabolic barrier (Simone et al., 1994; Gupta and Sastry, 2000) rather than a physical barrier.
The placenta plays the role of several organs and organ systems for the fetus throughout the course of development. The placenta takes on the role of lung, gut, kidney, and exocrine/endocrine glands (Juchau, 1985). In addition, the placenta can perform biotransformation and detoxication reactions. In fact, the placenta is capable of carrying out most of the biotransformation reactions that occur in liver (Juchau, 1980). Xenobiotic transporters are present and functional in placenta and may be a missing link between maternal exposure and embryo/fetal toxicity.
A thorough understanding of the role of various transporters in the placenta in the handling of xenobiotics across the maternal/fetal interface is essential to evaluate the pharmacological and toxicological potential of therapeutic agents, drugs of abuse, and other xenobiotics used by the mother during pregnancy. The goal of this study was to measure the gene expression of transporters that carry xenobiotics across membranes and to compare placental levels to levels in maternal liver and kidney, the major organs of excretion, to determine which transporters are likely to have a role in placental handling of xenobiotics.
Materials and Methods
Animals.
Sasco Sprague-Dawley female rats were obtained from Charles River Laboratories, Inc. (Wilmington, MA) at 60 days of age. Animals were housed in the Association for Assessment and Accreditation of Laboratory Animal Care-accredited facility at the University of Kansas Medical Center on a 12-h light/dark cycle at 70 to 72°F and provided rodent chow (Teklad; Harlan Sprague-Dawley, Inc., Indianapolis, IN) and water ad libitum. Animals were bred in house, and gestation day (gd1) 0 was established upon detection of the copulatory plug or the presence of sperm after overnight mating. Liver, kidney, and placenta were removed from five animals on gd 18 and 21, snap frozen in liquid nitrogen and stored at −80°C.
RNA Extraction.
Total RNA was extracted with RNAzol B reagent (Tel-Test, Inc., Friendswood, TX) according to the protocol of the manufacturer. RNA integrity was confirmed by formaldehyde agarose gel electrophoresis, and concentration was determined by ultraviolet absorbance at 260 nm. RNA was diluted and stored in sterile diethyl pyrocarbonate-treated dH2O at −80°C before analysis by the branched DNA (bDNA) signal amplification assay (High Volume QuantiGene bDNA Signal Amplification kit; Bayer Corp.-Diagnostics Div., Tarrytown, NY).
Branched DNA Signal Amplification Assay.
Forty transporter mRNA transcripts were analyzed with the bDNA assay (High Volume QuantiGene bDNA Signal Amplification kit) with modifications (Hartley and Klaassen, 2000). Rat gene sequences were accessed from GenBank (Table 1). Multiple oligonucleotide probes (containing capture, label, and blocker probes) specific to each of the 40 mRNA transcripts were designed using Probe Designer software v1.0 (Bayer Diagnostics). Probe sets were designed with a Tm of approximately 63°C, enabling hybridization conditions to be held constant (i.e., 53°C) during each hybridization step and for each probe set. Every probe developed in Probe Designer was submitted to the National Center for Biotechnological Information (NCBI, Bethesda, MD) for nucleotide comparison by the basic logarithmic alignment search tool (BLASTn) to ensure minimal cross-reactivity with other known rat sequences and expressed sequence tags. Oligonucleotides with a high degree of similarity (≥80%) to other rat gene transcripts were eliminated from the probe set design. The nucleotide sequences and function for the probes are reported in the following references: Mdr1a and Mdr1b (Brady et al., 2002); Mrp1, Mrp2, and Mrp3 (Cherrington et al., 2002); organic anion transporters (Oat 1, 2, and 3; Buist et al., 2002); Oct1, Oct2, Oct3, OctN1, and OctN2 (Slitt et al., 2002); organic anion-transporting polypeptide (Oatp 1, 2; Rausch-Derra et al., 2001) (Oatp 3, 4, 5; Li et al., 2002); and divalent metal transporter 1 (DMT1; Park et al., 2002). The remaining probe sets are listed in Fig.1 as follows: Mdr2; Mrp4, Mrp5, and Mrp6; sterolins 1 and 2 (AbcG5 and Na+/taurocholate-cotransporting polypeptide (Ntcp); bile salt export pump (Bsep); Oatp 9 and 12; organic anion transporter-K (Oat-K); prostaglandin (PGT); peptide (PEPT1 and 2); zinc transporter 1 (ZnT-1); Menkes (ATP7A); Wilson's (ATP7B); concentrative nucleoside (Cnt1 and 2); and equilibrative nucleoside (Ent1 and 2) transporters.
Gene names and accession numbers of transporters examined in placenta





Oligonucleotide probes generated for analysis of expression by bDNA signal amplification.
Capture extenders, label extenders, and blockers represent the function of each of the oligonucleotide probes in the bDNA assay.
Total RNA (1 μg/μl; 10 μl/well) was allowed to hybridize to each individual oligonucleotide probe set cocktail overnight at 53°C. Following the incubation of the probe set cocktail and the target RNA hybridization reaction, the amplifier was hybridized to the label extenders during a 1-h incubation at 46°C. Label probes conjugated to alkaline phosphatase were then hybridized to the amplifier molecules for 1 h at 46°C. Following each incubation, the plates were washed stringently, and excess buffer was aspirated from the plate to remove excess reactants from previous reactions. Finally, the labeled amplifier complexes were detected as chemiluminescence following 60- to 90-min incubation with the substrate dioxetane. Relative light units (RLU) are directly proportional to the amount of target RNA in each sample, and were measured by the Quantiplex 320bDNA luminometer interfaced with Quantiplex data management software version 5.02 (Bayer Diagnostics) for analysis of luminescence from 96-well microtiter plates. These data are reported as RLU/10 μg total RNA.
Statistical Analysis.
Means and standard error were calculated for 5 animals per gestation day (gd 18 and 21). The Student's t test was conducted to determine differences between gestation days within tissues. Few differences were observed; therefore, RLU from gd 18 and 21 were averaged and graphed as a combined sample. When differences between expression on gestation days 18 and 21 in placenta were apparent, they were noted in the results.
Results
All three members of the P-glycoprotein gene family, Mdr1a, Mdr1b, and Mdr2, were examined (Fig. 2). Placental Mdr1a mRNA levels were approximately twice those in kidney and 4 times those in liver in late stage pregnancy. Mdr1b mRNA levels were also highest in placenta, compared with liver and kidney in the pregnant rat. Placental Mdr1b mRNA levels were 4 times higher than in kidney and 15 times higher than in liver. Placental Mdr1b mRNA levels were nearly 2.5 times higher at gd 21 than at gd 18 (data not shown). Mdr2 mRNA expression was highest in liver. Placental Mdr2 mRNA levels were 3% of those in liver and about one-third those in kidney.

Placenta P-glycoprotein mRNA levels compared with maternal liver and kidney levels.
Total RNA was isolated from placenta, liver, and kidney of pregnant (gd 18 and 21) rats and analyzed with the branched DNA amplification assay for Mdrla, Mdr1b, and Mdr2 mRNA content. The data are presented as mean RLU ± S.E.M. per 10-μg total RNA.
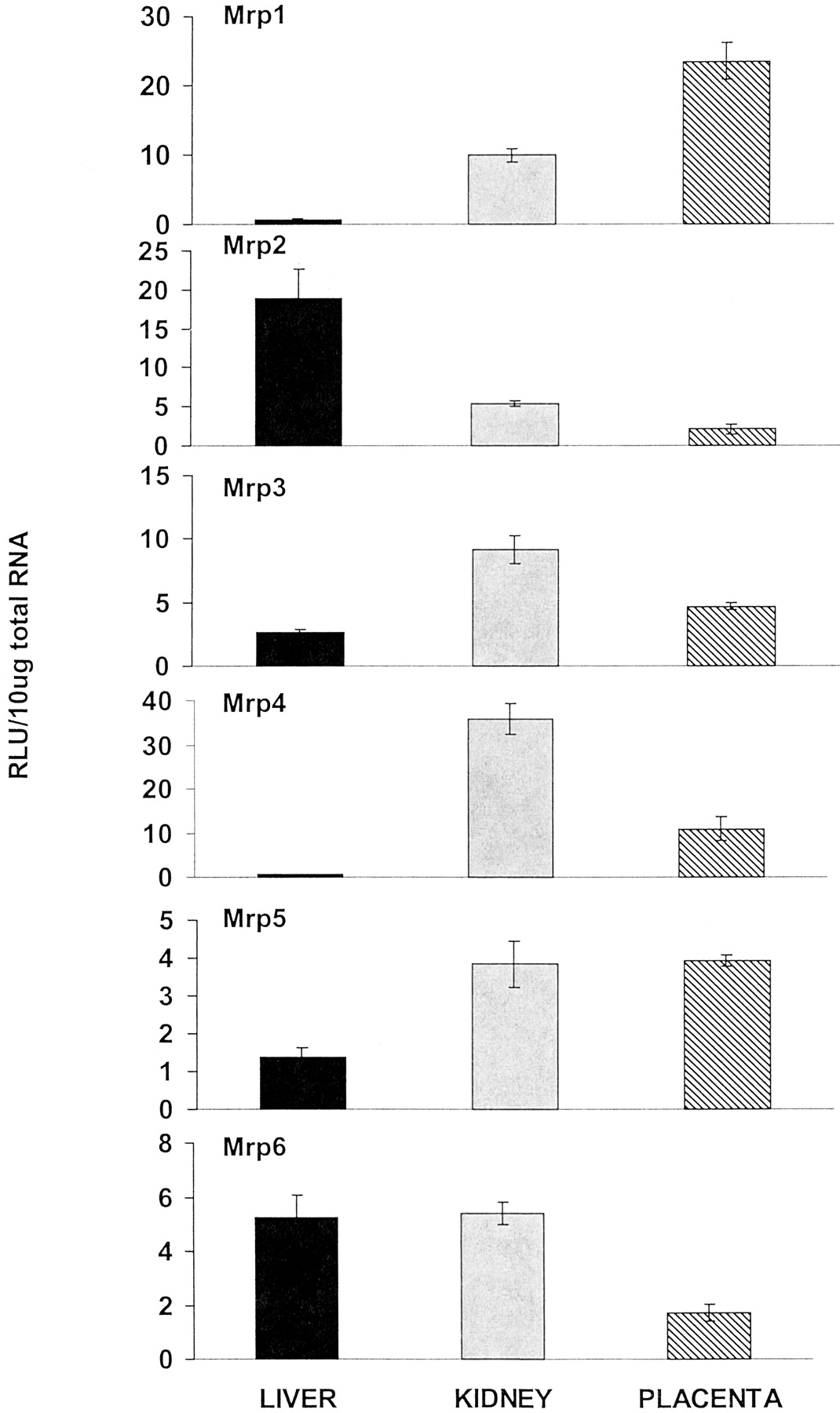
Six members of the multidrug resistance-associated protein drug transporter family were examined: Mrp1, Mrp2, Mrp3, Mrp4, Mrp5, and Mrp6 (Fig. 3). Mrp1 mRNA levels were expressed most abundantly in placenta compared with liver and kidney. Placental Mrp1 mRNA levels were more than twice those present in kidney and more than 40 times that present in liver. The apical membrane transporter Mrp2 mRNA levels were highest in liver. Placental Mrp2 was less than one-half that present in kidney and about one-tenth that in liver. Mrp3 mRNA levels were most abundant in kidney. Placental Mrp3 mRNA levels were about one-half that in kidney but nearly 2 times higher than liver. Mrp4 mRNA levels were highest in kidney. Placental Mrp4 levels were approximately 20 times higher than liver but were about one-third of those in kidney. Mrp5 mRNA levels were highest in placenta and kidney. Placental Mrp5 levels were approximately 3 times higher than liver and approximately equal to that in kidney. Mrp6 mRNA levels were highest in liver and kidney and lowest in placenta. Placenta Mrp6 mRNA levels were about one-third of those in liver and kidney. However, placental Mrp6 mRNA levels were 2.4 times higher at gd 18 than 21 (data not shown).

Placenta multidrug resistance-associated protein transporter mRNA levels compared with maternal liver and kidney levels.
Total RNA was isolated from placenta, liver, and kidney of pregnant (gd 18 and 21) rats and analyzed with the branched DNA amplification assay for Mrp1, Mrp2, Mrp3, Mrp5, and Mrp6 mRNA content. The data are presented as mean RLU ± S.E.M. per 10-μg total RNA.
AbcG5 and AbcG8 were also assessed (Fig.4). AbcG5 mRNA levels were highest in liver. Placental levels were about three-fourths of those in kidney and about one-eighth of those in liver. AbcG8 levels were highest in placenta, and placental levels were more than 1.5 times that in both liver and kidney.

Placenta sterolin transporter mRNA levels compared with maternal liver and kidney levels.
Total RNA was isolated from placenta, liver, and kidney of pregnant (gd 18 and 21) rats and analyzed with the branched DNA amplification assay for AbcG5 and AbcG8 mRNA content. The data are presented as mean RLU ± S.E.M. per 10-μg total RNA.
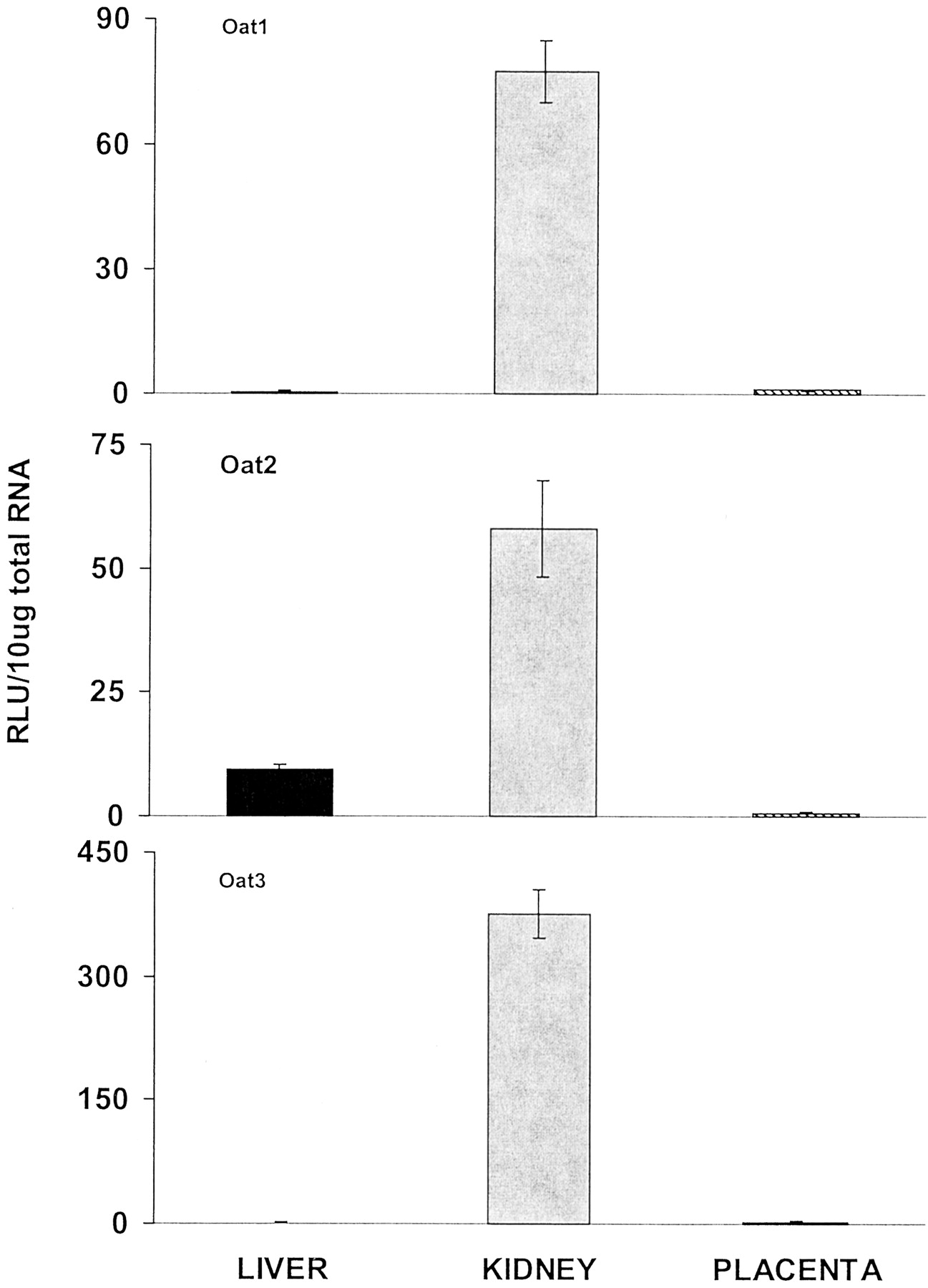
The three organic anion transporters, Oat1, Oat2, and Oat3, were determined (Fig. 5). The mRNA levels of each of these transporters were predominant in kidney, whereas placental levels were extremely low.

Placenta organic anion transporter mRNA levels compared with maternal liver and kidney levels.
Total RNA was isolated from placenta, liver, and kidney of pregnant (gd 18 and 21) rats and analyzed with the branched DNA amplification assay for Oat1, Oat2, and Oat3 mRNA content. The data are presented as mean RLU ± S.E.M. per 10-μg total RNA.
The bile acid transporters, Ntcp and sister of P-glycoprotein or bile salt export protein (Bsep) were also examined (Fig.6). Ntcp mRNA levels were highest in liver. Placental levels were less than 1% of those in the liver. Bsep mRNA levels were also highest in liver. Placental Bsep mRNA was similar to that of kidney, which was about five percent of that in liver.

Placenta bile acid transporter mRNA levels compared with maternal liver and kidney levels.
Total RNA was isolated from placenta, liver, and kidney of pregnant (gd 18 and 21) rats and analyzed with the branched DNA amplification assay for Ntcp and Bsep mRNA content. The data are presented as mean RLU ± S.E.M. per 10-μg total RNA.
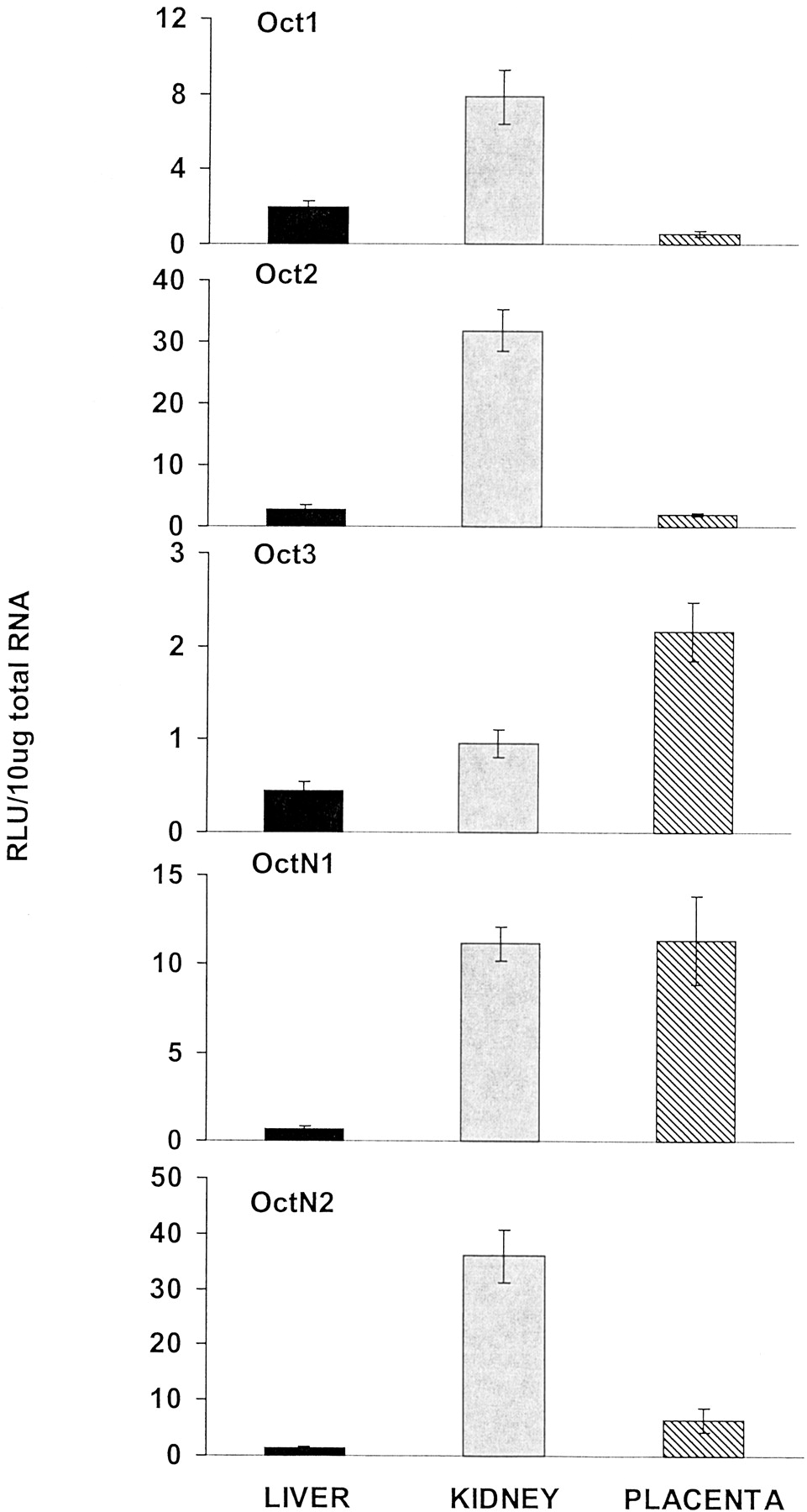
Five members of the rat Oct family, namely Oct1, Oct2, Oct3, OctN1, and OctN2, were examined in the current study (Fig.7). Oct1 mRNA levels were highest in kidney. Placental Oct1 mRNA levels were approximately one-third of those present in liver and less than one-tenth of those found in kidney. Oct2 mRNA levels were also highest in kidney. Placental and liver Oct2 mRNA levels were low. Oct3, originally cloned from placenta (Kekuda et al., 1998), had 2.5 times higher levels in placenta than liver and 5 times higher mRNA levels in placenta than kidney, however the levels of this gene were low in all tissues. OctN1 mRNA levels were highest in placenta and kidney, which was about 18 times higher than those in liver. Placental OctN1 mRNA levels on gd 21 were nearly 3 times higher than gd 18 (data not shown). Like Oct1 and Oct2, OctN2 mRNA levels were highest in kidney. Placental OctN2 levels were about 6 times higher than those in liver but about 18% of those in kidney.

Placenta organic cation transporter mRNA levels compared with maternal liver and kidney levels.
Total RNA was isolated from placenta, liver, and kidney of pregnant (gd 18 and 21) rats and analyzed with the branched DNA amplification assay for Oct1, Oct2, Oct3, OctN1, and OctN2 mRNA content. The data are presented as mean RLU ± S.E.M. per 10-μg total RNA.
Eight members of the Oatp family were examined in this study: Oatp1, Oatp2, Oatp3, Oatp4, Oatp5, Oatp9, Oatp12, and Oat-K (Fig.8). Oatp1 mRNA levels were most prominent in liver, whereas kidney and placenta were similarly low. Placenta Oatp1 mRNA levels were about one-fifth those determined in liver but nearly equal to those in kidney. Oatp2 mRNA levels were also highest in liver, with the levels in kidney and placenta less than 50 and 20%, respectively. Oatp3 mRNA expression was most abundant in placenta and kidney but lowest in liver. Placental Oatp3 mRNA levels were about 2 times those in liver but similar to the level in kidney. Oatp4 (liver-specific transporter) mRNA levels were predominant in liver, with extremely low levels in kidney and placenta. Oatp5 mRNA levels were highest in kidney and extremely low in liver and placenta. Oatp9 mRNA levels were highest in liver. Placental Oatp9 mRNA levels were about one-third of those in liver and about one-fourth of those in kidney. However, placental Oatp9 levels were nearly 3 times higher on gd 21 than those on gd 18 (data not shown). Oatp12 mRNA levels were highest in placenta. Placental levels were more than 45 times higher than kidney, and liver levels were negligible compared with placenta. Placental Oatp2 mRNA levels were nearly 5 times higher on gd 21 than those in gd 18 (data not shown). Oat-K mRNA levels were highest in kidney. Placental Oat-K mRNA levels were similar to liver but were about one-tenth of those in kidney.

Placenta organic anion transporter protein mRNA levels compared with maternal liver and kidney levels.
Total RNA was isolated from placenta, liver, and kidney of pregnant (gd 18 and 21) rats and analyzed with the branched DNA amplification assay for Oatp1, Oatp2, Oatp3, Oatp4, Oatp5, Oatp9, Oatp12, and Oat-K mRNA content. The data are presented as mean RLU ± S.E.M. per 10-μg total RNA.
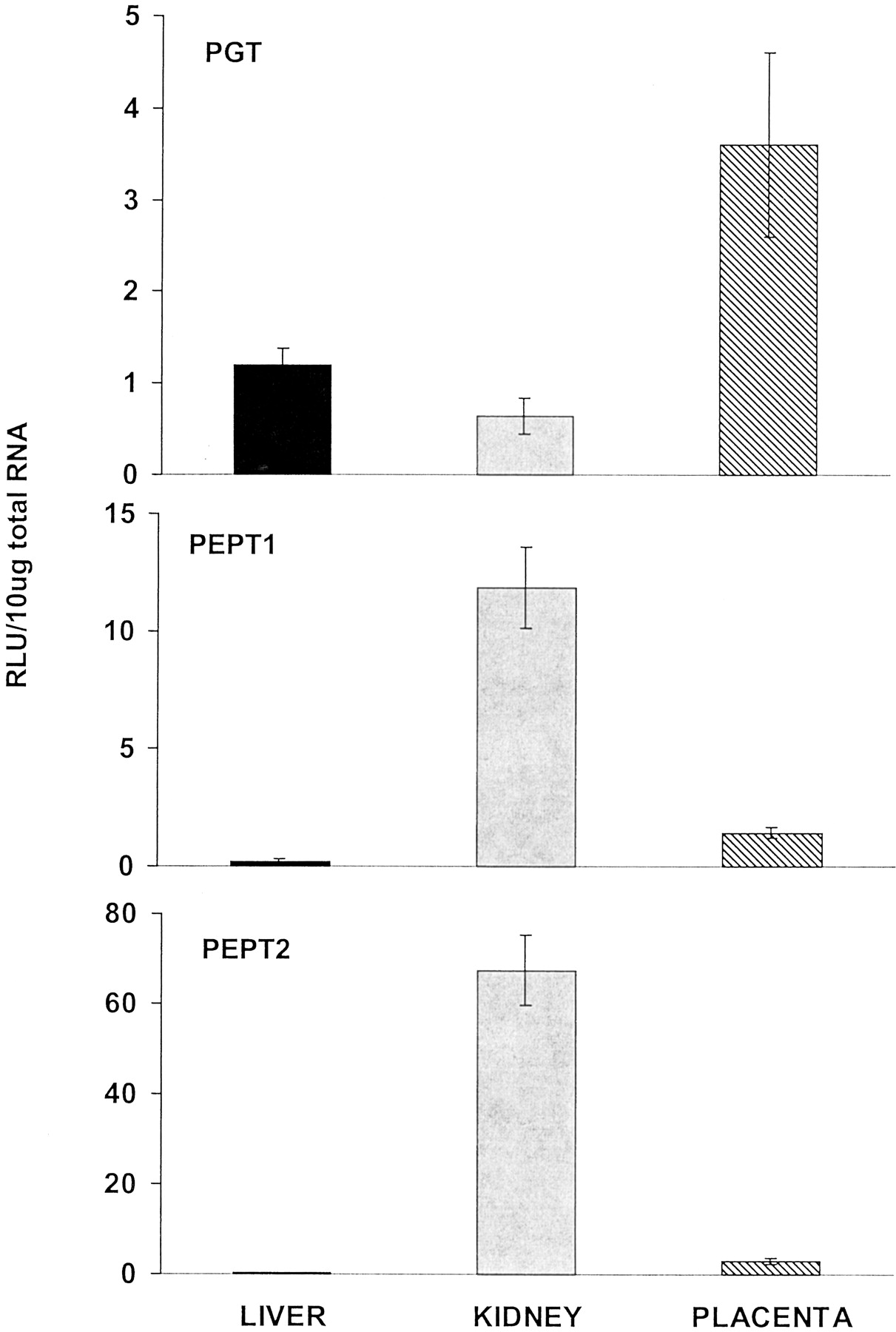
The PGT and proton-coupled PEPT1 and PEPT2 were also examined (Fig.9). PGT mRNA levels were predominant in placenta. Placental levels of PGT mRNA were 6 times higher than in kidney and more than 2 times higher than in liver. Placental PGT mRNA levels on gd 21 were 5 times higher than gd 18 mRNA levels (data not shown). PEPT1 and PEPT2 were both high in kidney. Placental PEPT1 levels were about one-eighth of those in kidney but were 6 times higher than those in liver. Placental PEPT2 mRNA levels were about 20% of those in kidney but about 8 times higher than liver.

Placenta prostaglandin and peptide transporter mRNA levels compared with maternal liver and kidney levels.
Total RNA was isolated from placenta, liver, and kidney of pregnant (gd 18 and 21) rats and analyzed with the branched DNA amplification assay for PGT, PEPT1, and PEPT2 content. The data are presented as mean RLU ± S.E.M. per 10-μg total RNA.
The following metal transporter mRNA levels were also assessed: ZnT-1, DMT1, Menkes, and Wilsons (Fig. 10). ZnT-1 mRNA levels were highest in kidney. Placental ZnT-1 mRNA levels were 5 times higher than those in liver and about 75% of those in kidney. Placental ZnT-1 mRNA levels were about 42% higher at gd 21 than 18 (data not shown). DMT1 mRNA levels were highest in placenta. Placental levels were about 25 times higher than liver and about 1.5 times higher than kidney. DMT1 mRNA levels in placenta were 2 times higher on gd 21 than on gd 18 (data not shown). Menkes mRNA levels were most abundant in placenta. Menkes mRNA levels in placenta were more than 1.5 times higher than kidney and about 14 times higher than liver. Menkes mRNA levels in placenta were 42% higher on gd 21 than 18 (data not shown). Wilsons mRNA was highest in liver. Placental levels were about 75% of those in kidney and about 60% of those in liver.

Placenta metal transporter mRNA levels compared with maternal liver and kidney levels.
Total RNA was isolated from placenta, liver, and kidney of pregnant (gd 18 and 21) rats and analyzed with the branched DNA amplification assay for ZnT-1, DMT-1, and Menkes and Wilson mRNA content. The data are presented as mean RLU ± S.E.M. per 10-μg total RNA.
Sodium-linked Cnt1 and Cnt2 and Ent1 and Ent2 were assessed in placenta (Fig. 11). Cnt1 mRNA levels were most abundant in kidney. Placental Cnt1 mRNA levels were one-fourth of those in liver and were very low (1:25) compared with those in kidney. Cnt2 levels were highest in liver. Placental Cnt2 mRNA levels were about half of those in liver and about two-thirds of those in kidney. Placental Cnt2 mRNA levels were 2.5 times higher at gd 21 than 18 (data not shown). Ent1 mRNA levels were most abundant in placenta. Placental Ent1 levels were more than 1.5 times those in kidney and 5 times those in liver. Ent2 mRNA levels were highest in placenta and kidney. Placental levels, while very similar to kidney, were 5 times higher than those in liver.

Placenta sodium-linked nucleoside transporters and equilabrative nucleoside transporter mRNA levels compared with maternal liver and kidney levels.
Total RNA was isolated from placenta, liver, and kidney of pregnant (gd 18 and 21) rats and analyzed with the branched DNA amplification assay for Cnt1, Cnt2, Ent1, and Ent2 mRNA content. The data are presented as mean RLU ± S.E.M. per 10-μg total RNA.
Discussion
Placenta, the sole link between mother and fetus, performs a multitude of functions required for normal fetal development. Among the many functions are the transfer of nutrients from mother to fetus, the exchange of metabolic waste from fetus to mother, and the biotransformation of xenobiotics. The placenta uses transporters to provide nutrients and other essential chemicals to the fetus and to protect the fetus from some xenobiotics to which the pregnant animal is exposed. Liver and kidney are the two major organs that use transporters in the excretion of xenobiotics. Therefore, both liver and kidney were compared with the level of xenobiotic transporter mRNA in term placenta. The current work was designed to quantitatively determine the level of mRNA of 40 transporter genes in placenta and compare this expression with the expression in tissues predominantly involved in elimination of xenobiotics.
The P-glycoproteins (P-gp) (members of the ATP binding cassette gene family) are encoded by two groups of the Mdr gene family. Group I Mdr genes encode P-gps that have been shown to mediate drug transport and clinical tumor drug resistance. Rodents contain two group I genes (denoted Mdr1a and 1b), both encoding proteins that are capable of drug transport. Rodent P-gps are associated with chemotherapeutic drug resistance and intestinal excretion. Additionally, they are present in the blood-brain barrier (Fromm, 2000). Thus, the function of P-gp may be to protect cells from naturally occurring toxins. The group II gene product, Mdr2, which contains extensive sequence identity to group I, is associated with phospholipid transport and excretion of lipid into bile, rather than with drug resistance (Smit et al., 1999). The mRNA levels of Mdr1a in rats are highest in the gastrointestinal tract (Brady et al., 2002). The present data indicate that Mdr1a is 2 times higher in placenta than in kidney and 4 times higher in placenta than in liver. Mdr1b is about 15 times higher in placenta than in kidney and liver (Fig. 2). In contrast, very little Mdr2 mRNA was detected in the placenta in comparison to that found in liver. These data indicate that Mdr1a and 1b levels are relatively high in placenta and may protect the fetus from xenobiotics. In fact, Lankas et al. (1998) provided evidence to support a role for P-gp in protection of the fetus from the teratogenic pesticide, avermectin. They showed that Mdr1a is present in the fetal-derived epithelial cells that make up the exchange border between the fetal and maternal blood circulation, with Mdr1a facing the maternal blood side. In naturally occurring Mdr1a mutant mice of the CF-1 outbred mouse stock that lack Mdr1a (Umbenhauer et al., 1997), P-gp is associated with enhanced sensitivity of the fetus to an isomer of the pesticide avermectin. Specifically, fetuses deficient in P-gp were 100% susceptible to cleft palate, whereas the heterozygote (±) littermates were less sensitive. Moreover, the homozygous fetuses (+/+) with abundant P-gp were totally insensitive to the avermectin isomer.Umbenhauer et al. (1997) further showed that enhanced fetal drug penetration paralleled the increased avermectin sensitivity in Mdr1a mutant fetuses. Furthermore, with the Mdr1a/Mdr1b knockout mouse model,Smit et al. (1999) determined that the drug transporting P-gp at the fetoplacental unit serves to limit placental transfer of P-gp substrates such as digoxin, saquinavir, and paclitaxel from the maternal circulation to the fetus. These knockout mice accumulated higher levels of the P-gp substrates than either the homozygous or heterozygous animals. They further showed that inhibitors of P-gp could enhance the accumulation of the P-gp substrates in all genetic variations of this mouse model. These data suggest that the presence of P-gp in placenta is of great importance in limiting fetal penetration of various compounds.
Mrp are members of the ATP-binding cassette transporter superfamily and are important in ATP-dependent transport of many organic anions, including many phase II metabolites (Jedlitschky et al., 1997; Kawabe et al., 1999). Mrp1, 2, and 3 have all been shown to export and confer resistance to cytotoxic drugs (Stockel et al., 2000). In liver, mrp1 and 3 are located on the sinusoidal membrane (Konig et al., 1999; Kool et al., 1999), whereas Mrp2 is localized to the apical membrane. This implies that in liver, Mrp2 mediates excretion into bile, whereas Mrp1 and 3 mediate transport into blood, which leads to excretion into urine. Cherrington et al. (2002) indicated that Mrp1 mRNA was expressed at relatively similar levels in all tissues examined, with the exception of low levels in liver, however placenta was not examined in that study. The current data (Fig. 3) indicate that Mrp1 is more abundantly expressed in placenta than in any other tissue in that study. In contrast, Mrp2 levels were most abundant in liver, and its expression in the placenta was about one-fourth that in liver. In the present study, placental Mrp3 mRNA levels were twice as much as liver, but about one-half that in kidney. Recent studies have localized Mrp3 primarily to the fetal blood endothelia of term placenta, with some additional evidence for expression in the syncytiotrophoblast layer (St-Pierre et al., 2000). Mrp4 mRNA levels were approximately 20 times higher in placenta than liver, but were about one-third of those in kidney. Mrp5 was most abundantly expressed in placenta and kidney, which was more than twice that in liver. In contrast, Mrp6 mRNA in placenta was less than one-half that in kidney and liver. These data suggest that of the Mrp family of transporters, Mrp1 and 5 are most likely to protect the fetus from xenobiotics at the placental level.
Sterolin half-transporters are encoded from the AbcG5 and AbcG8 ATP binding cassette genes. Sterolin transporters are thought to function by limiting the intestinal absorption of sterols (plant sterols) and enhancing the excretion of sterols from liver into bile (Goldstein and Brown, 2001; Lee et al., 2001; Lu et al., 2001). Mutations in AbcG5 and AbcG8 result in the human disease sitosterolemia (Heimer et al., 2002), which is a rare autosomal recessive disorder characterized by highly elevated plasma levels of plant sterols and cholesterol as a consequence of hyperabsorption and impaired biliary secretion of sterols. The present data indicate that AbcG8 mRNA levels were highest in placenta, and placental levels were more than 1.5 times those in both liver and kidney (Fig. 4). Although transcript levels are generally low, their presence indicates a role for the sterolin transporters in the elimination of sterols, particularly plant sterols, within the placenta. Based on unpublished tissue distribution data from our lab, small intestine has higher expression of AbcG5 and AbcG8 than liver or kidney in rat.
In general, the organic anion transporter (Oats) mRNA levels were predominant in kidney of pregnant rats (Fig. 5), which corresponds to the tissue distribution of Oat1, 2, and 3 reported in male and nonpregnant female rats (Buist et al., 2002). Very little Oat1, 2, or 3 mRNA was present in placenta, therefore it is not likely that these transporters play an important role in protecting the fetus from xenobiotics. In humans, Oat4 has been cloned from placenta but has not been detected in rats (Cha et al., 2000).
Bile acid transporters are thought to exist in the placenta to protect the fetus from exposure to bile acids in the maternal blood and to facilitate transport of fetal-derived bile acids by vectorial translocation from fetal to maternal circulation before elimination (Marin et al., 1990). Ntcp is located in the sinusoidal membrane of the hepatocyte to transport bile acids into the liver from the blood. Physiological and pathophysiological stimuli, such as pregnancy and cholestasis, modulate Ntcp. Bile salt export protein (Bsep or sister of P-glycoprotein) is considered the fourth member of the P-gp family. Bsep is associated with hepatic bile salt excretion and is implicated in the disease progressive familial intrahepatic cholestasis 2 (Gerloff et al., 1998; Strautnieks et al., 1998). In the present study, Ntcp and Bsep were present most abundantly in the maternal liver, with low mRNA levels present in placenta (Fig. 6). Thus, it is not clear which transporter is responsible for protecting the fetus from bile acids.
Organic cation transporters are responsible for the uptake of prototypical organic cations, such as tetraethylammonium; secretion of endogenous amines, such as dopamine; as well as secretion of cationic drugs, such as antihistamines and antiarrhythmics (Zhang et al., 1998;Burckhardt and Wolff, 2000). Slitt et al. (2002) reported the tissue distribution of the organic cation transporter family and determined that the mRNA levels were highest in kidney for Oct1, Oct2, OctN1, and OctN2, whereas Oct3 was highest in blood vessel. The present study indicates the placenta expresses relatively high levels of Oct3 and OctN1, and relatively low levels of Oct1, Oct2, and OctN2 (Fig. 7). In fact, Oct3 mRNA levels in placenta are more than twice those in the kidney and five times those in liver. Octn1 is expressed at the same level in the placenta as in the kidney, whereas its expression in the liver is very low.
Oct3 is likely expressed in the fetal-facing basal membrane of the placenta, where it transports cationic drugs from the fetal circulation into the placental trophoblast (Kekuda et al., 1998). Once inside the trophoblast, these drugs can be eliminated into the maternal circulation across the maternal-facing brush border membrane, mediated by the organic cation H+-gradient-dependent system known to be present in this membrane. Hence, Oct3 may be a key player in the barrier function of the placenta protecting the developing fetus from possible deleterious effects of endobiotics produced by the fetus, as well as xenobiotics that may be present in the maternal circulation. Oct3 transcripts have also been found in the kidney and intestine, two other organs known to be involved in the elimination of drugs. Interestingly, Oct3 is not highly expressed in liver, the major organ responsible for metabolism and elimination of xenobiotics (Kekuda et al., 1998). The current study indicates that Oct3 and OctN1 were most abundant in placenta, whereas Oct1, Oct2, and OctN2 were highest in kidney (Fig. 7).
The Oatp family transports structurally unrelated compounds, such as anions, cations, and neutral compounds, in a sodium- and ATP-independent manner (Meier et al., 1997; Noe et al., 1997; Abe et al., 1998; Eckhardt et al., 1999). The tissue distribution of the Oatp was determined in rats, and the results indicated that Oatp1 mRNA levels were highest in male kidney (Lu et al., 1996b; Li et al., 2002) but were also high in liver of male and female rats. No gender difference was apparent in Oatp1 mRNA levels in liver, but much higher levels were detected in male kidney than female kidney. Female kidney levels of Oatp1 mRNA were relatively low. Oatp1 mRNA levels were also low in placenta. Oatp2 mRNA is mainly expressed in liver and brain (Noe et al., 1997; Li et al., 2002), with low expression in the placenta. Oatp3 was highest in lung (Walters et al., 2000; Li et al., 2002), with much lower levels in liver and kidney. Placental expression of Oatp3 is similar to that in kidney and liver. Oatp4, also called liver-specific transporter, is predominately expressed in liver (Kakyo et al., 1999;Choudhuri et al., 2000; Li et al., 2002), with minimal expression in placenta. Oatp5 mRNA was almost exclusively expressed in kidney, with minimal expression in other tissues (Li et al., 2002). Oatp9 mRNA, recently discovered as moat1 to transport taurocholate, prostaglandin D, and leukotrienes (Nishio et al., 2000), was present in placenta, however the levels were lower than in liver and kidney. Oatp12 mRNA levels, recently determined to transport thyroid hormone in various peripheral tissues (Fujiwara et al., 2001), were most abundant in placenta (Fig. 8). Oat-K mRNA levels were minimal in placenta as well. The current report determined with the exception of Oatp12, that expression of all Oatps in the placenta was minimal, thus, it is not likely that the Oatp family of transporters plays any major role in fetal protection or xenobiotic transfer by the placenta. However the levels of Oatp12 detected in placenta indicate a means for thyroid hormone transport within the placenta.
Functional analysis has demonstrated the transfer of prostaglandin across the placenta (Glance et al., 1986). In addition, the PGT mRNA has been detected in placenta (Lu et al., 1996a). However, membrane localization of PGT in placenta is not known. Prostaglandins and thromboxanes are known substrates for PGT, which have critical influences on placental circulation and function. These substrates and their synthetic analogs are particularly involved in the termination of pregnancy (Lu et al., 1996a). In addition, furosemide, a widely used diuretic, is also a substrate for PGT. PGT is an electrogenic obligatory anion exchanger (Chan et al., 1998), which suggests that this transporter functions in the uptake of prostaglandins and thromboxanes into the syncytiotrophoblast. In the present study, PGT mRNA levels were higher in placenta than either liver or kidney (Fig.9). The presence of PGT mRNA in placenta supports the role for this transporter in the placental handling of these compounds and also in the pregnancy-dependent alterations in their pharmacokinetics.
PEPT1 and 2 mediate the cellular uptake of small intact peptides consisting of two or three amino acids (Leibach and Ganapathy, 1996). PEPTs are critical to the absorption and reclamation of small peptides in the epithelial cells of the intestinal tract and kidney tubules (Leibach and Ganapathy, 1996). PEPT1 and 2 are members of the proton-coupled oligopeptide transporter family and are energized by a transmembrane electrochemical H+ gradient. PEPTs have similar substrate specificity but differ in their affinity for substrates. PEPT1 is a low-affinity, high-capacity transporter located primarily in intestine but also in kidney, whereas PEPT2 is a high-affinity, low-capacity transporter abundantly expressed in kidney (Leibach and Ganapathy, 1996; Fei et al., 1998). In addition to transporting the natural substrates, PEPTs are also capable of transporting pharmacologically active compounds including β-lactam antibiotics, antitumor agents, as well as inhibitors of renin and angiotensin converting enzymes (Leibach and Ganapathy, 1996). The current study assessed the peptide transporters in placenta and maternal liver and kidney and found that kidney mRNA levels were predominant. Placental levels were higher than liver but were considerably less than kidney (Fig. 9). These data do not support a prominent role for PEPT in fetal protection by the placenta.
Metal transporters were assessed in placenta, as trace metal homeostasis is vital for normal fetal development and maintenance of life. Trace levels of zinc, copper (Cu), and iron are required for various molecular processes including synthesis of genes, serving as cofactors for transcription factors, and enzyme function.
ZnT1, ubiquitously located in the plasma membrane, was proposed to function as an exporter to maintain zinc homeostasis (Palmiter and Findley, 1995). ZnT1 has been localized to the basolateral membrane of the renal tubule as well as duodenum and jejunum of the small intestine, suggesting it functions in zinc reabsorption by the renal tubules and zinc acquisition and/or retention by the small intestine (McMahon and Cousins 1998a,b). ZnT1 mRNA has been detected in mouse placenta and was not subject to regulation by dietary zinc (Langmade et al., 2000). Placental ZnT1 mRNA levels were 5 times higher than liver but about three-quarters of those in kidney (Fig. 10).
DMT1 is primarily responsible for dietary iron uptake in the duodenum and iron acquisition from transferrin in other tissues. However, DMT1 is not limited to iron transport, as it is known to transport other divalent metals such as zinc, cadmium, copper, and cobalt (Gunshin et al., 1997; Picard et al., 2000; Olivi et al., 2001). In addition to small intestine, DMT1 has been localized to hepatocyte plasma membrane (Trinder et al., 2000), kidney (Ferguson et al., 2001), and placenta (basal, fetal facing) (Georgieff et al., 2000). In the present study, DMT1 mRNA levels were highest in placenta, and placental levels were about 25 times higher than in liver and about 1.5 times higher than in kidney (Fig. 10).
Trace Cu is important for normal development and is necessary to sustain life, but excess can be toxic. Menkes (MNK; ATP7A) and Wilson's (WND; ATP7B) genes are involved in the transport of Cu and are the genes affected in Menkes syndrome and Wilson disease in humans. Both conditions result in Cu toxicity by different mechanisms. ATP7A and ATP7B genes are defective in Menkes and Wilson disease, respectively, and encode transmembrane P-type ATPase proteins (MNK and WND) that function to translocate Cu across cellular membranes. MNK and WND have a high degree of similarity at the protein level (Monaco and Chelly, 1995). At the level of the whole organism, these two proteins appear to have distinct roles in Cu transport and homeostasis.
Menkes disease is a fatal, X-linked Cu deficiency disorder that results from defective copper efflux from intestinal cells and inadequate Cu delivery to other tissues, leading to deficiencies of critical Cu-dependent enzymes. Failure to transport copper to the fetus during development results in reduced activity of Cu-dependent enzymes, leading to severe mental retardation; connective tissue abnormalities; steely, white, brittle hair; and ultimately death by the age of three years. Wilson disease is an autosomally inherited, Cu toxicosis disorder resulting from defective biliary excretion of Cu, resulting in Cu accumulation primarily in liver but also in the brain, kidney, cornea, and spleen (Royce et al., 1980; Bingham et al., 1998). In addition, serum plasma Cu concentrations are reduced because of a failure to incorporate Cu into the apo-form of ceruloplasmin before its release into serum. Symptoms of Wilson disease include hepatitis, cirrhosis, neurological damage, and psychiatric disorders, as well as renal abnormalities, hematologic disturbances, and endocrine dysfunction (Sternlieb, 1980; Brewer and Yuzbasiyan-Gurkan, 1992;Bingham et al., 1998).
Menkes (ATP7A) mRNA was previously determined to be expressed highest in muscle, kidney, lung, and brain, moderately in placenta and pancreas and lowest in liver (Chelly et al., 1993; Mercer et al., 1993; Vulpe et al., 1993). Wilson (ATP7B) mRNA levels were found to be predominant in liver and at lower levels in a number of other tissues (Bull et al., 1993; Tanzi et al., 1993). In the present study, Menkes mRNA levels were highest in placenta. Placental levels were more than 1.5 times the levels in kidney and about 14 times higher than liver (Fig. 10). Wilson mRNA levels were predominant in liver. Placental levels were about 60% of those in liver and about 75% of those in kidney. The placental mRNA levels of all the metal transporters were indicative of a possible role in placental function.
Cnt1 and 2 are Na+-dependent transporters that mediate active uptake of purine and pyrimidine nucleosides as well as nucleoside analogs by coupling to the inwardly directed Na+ gradient across the plasma membrane (Wang and Giacomini, 1997). Cnts have been found in heart, skeletal muscle, placenta, pancreas, and lung (Cass et al., 1999). Ent1 and 2 are two different Na+-independent transporters (Griffiths et al., 1997). Both mediate the transport of purine and pyrimidine nucleosides, such as adenosine and uridine, but are characterized by their difference in sensitivity to inhibition by nitrobenzylthioinosine. Ent1 is sensitive to inhibition, whereas Ent2 is relatively insensitive to inhibition. Immunolocalization studies have shown that Ent1 is expressed in the placental brush border (apical membrane) (Barros et al., 1995), however the location of Ent2 is unknown. Ent1 and Ent2 are energy-independent transporters and are capable of facilitating only equilibrative, not concentrative, transport of nucleosides across the membrane. These nucleoside transporters can also transport anticancer nucleoside analogs (Griffiths et al., 1997) as well as antiviral agents (Domin et al., 1993). The present study quantitates the levels of mRNA in placenta compared with the other major organs of excretion, liver and kidney, and shows that Cnt1 mRNA levels were most abundant in kidney. Placental Cnt1 levels were one-fourth of those in liver and were very low (1:25) compared with kidney. Cnt2 levels were highest in liver. Placental mRNA levels of Cnt2 were about half of those in liver and about two-thirds of those in kidney (Fig. 11). Ent1 was highest in placenta whereas Ent2 was highest in placenta and kidney. These transporters are expected to facilitate the transfer of these nucleoside analogs across the placenta from the mother to the fetus (Ganapathy et al., 2000).
Although the functions of the placenta are many, the importance of the placenta in preventing xenobiotics from reaching the fetus has not been well examined. Whereas it is known that the presence of Mdr transporters in the placenta can decrease exposure of the fetus to some xenobiotics such as avermectin, the importance of other xenobiotic transporters in protecting the fetus is not known. The present study indicates that Mdr1a and 1b are highly expressed in the placenta, which corroborates their demonstrated function of protecting the fetus from xenobiotics. However, not all families of xenobiotic transporters are highly expressed in the placenta. For example, none of the Oat family members were highly expressed in placenta, suggesting that they do not play an important role in protecting the fetus from xenobiotics. Although, it is possible that some transporters with low mRNA levels in placenta may have highly specific localized expression, which is diluted when the entire placenta or a sizable amount of the placenta is homogenized. However, some individual transporters of other gene families are relatively highly expressed in the placenta and may play a role in protecting the fetus from xenobiotic exposure. For example, two members of the Oatp family, Oatp3 and Oatp12, have relatively high mRNA levels in placenta, and some members of the Mrp family also have relatively high levels in the placenta, namely Mrp1 and 5. Because the Mrp family of transporters is known to excrete conjugates of endobiotics and xenobiotics, these transporters might be important in the transport of conjugates formed in the placenta into maternal blood. Two members of the Oct family of transporters, namely Oct3 and OctN1 are relatively highly expressed in the placenta and might protect the fetus from various cationic endobiotics and xenobiotics. The prostaglandin transporter, also highly expressed in placenta, is known to modulate prostaglandins and thromboxanes within the maternal-fetal interface. These physiological substrates are known to have an effect on the maintenance of pregnancy and the initiation of labor and delivery (Mitchell et al., 1995). Metals are essential for normal fetal development. The metal transporters were present at relatively high levels in the placenta. However, these transporters are not specific for essential metals as some are known to transport nonessential metals such as, the known rodent teratogen, cadmium (Gunshin et al., 1997). Ent1 and Ent2 mRNA levels were relatively high in placenta. These nucleoside transporters are thought to transport nucleosides from mother to fetus but are not specific for physiological substrates. Nucleoside transporters can also transfer other chemicals like antivirals and cancer chemotherapeutics across the placental membranes. Quantification of these transporter transcripts in placenta provides important information toward understanding chemical handling within the maternal-fetal interface and provides insight into the role the placenta might play in preventing xenobiotics from reaching the fetus.
Acknowledgments
We thank Dr. Dylan Hartley and Dr. Nathan Cherrington for designing the oligonucleotide probe sets. We also thank Susan Buist for her technical assistance.
Footnotes
-
Research supported by National Institute of Environmental Health Sciences Grants ES-03192, ES-09716, and ES-09649 and training Grant ES-07079.
- Abbreviations used are::
- gd
- gestation day
- bDNA
- branched DNA
- Ntcp
- Na+/taurocholate-cotransporting polypeptide
- Bsep
- bile salt export protein
- Oatp
- organic anion-transporting polypeptide
- Oat-K
- organic anion transporter K
- PGT
- prostaglandin transporter
- PEPT
- peptide transporter
- ZnT-1
- zinc transporter 1
- ATP7A
- Menkes transporter
- ATP7B
- Wilson's transporter
- Cnt
- concentrative nucleoside transporter
- Ent
- equilibrative nucleoside transporter
- RLU
- relative light units
- Mdr
- multiple drug resistance
- Oct
- organic cation transporter
- DMT1
- divalent metal transporter 1
- P-gp
- P-glycoproteins
- Mrp
- multidrug resistance-associated proteins
- AbcG5 and AbcG8
- sterolin transporters 1 and 2
- Received August 23, 2002.
- Accepted October 21, 2002.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}