


Abstract
Messenger RNA levels of rat organic anion transporter 1 (Oat1;Slc22a6) and Oat2 (Slc22a7) in kidney and Oat3 (Slc22a8) in liver are gender-predominant. Oat1 and Oat3 are male-predominant, whereas Oat2 is female-predominant. Gonadectomized and hypophysectomized (HX) rats were studied to determine whether sex steroids and/or growth hormone (GH) are responsible for these gender-divergent patterns. GH was administered to HX rats by two daily injections (simulating male secretion) or continuous infusion (simulating female secretion). Oat1 mRNA levels, normally higher in male than female kidney, were lowered by gonadectomy and HX in male rats, but not in females. Additionally, GH injections or infusion did not alter Oat1 levels in HX rats. Oat2 mRNA levels, typically much higher in female than in male kidney, were unaffected by gonadectomy. However, HX dramatically decreased Oat2 in female kidney without altering male levels. GH administered by continuous infusion increased Oat2 in kidneys of both HX male and female rats, whereas injections had no affect. Gonadectomy reduced Oat3 mRNA levels in male livers without affecting levels in female livers. In contrast, HX decreased male and elevated female Oat3 mRNA. GH injections did not significantly change Oat3 mRNA levels in HX rats, but infusion decreased Oat3 mRNA in liver. In conclusion, androgens, but not GH, are responsible for the Oat1 mRNA gender difference in kidney; the female GH secretion pattern is responsible for the Oat2 mRNA gender difference in kidney; and both androgens and the female GH secretion pattern are responsible for the Oat3 mRNA gender difference in liver.
Metabolism and excretion both determine elimination of endogenous and exogenous compounds. Liver is the primary site of metabolism, and liver and kidney are important organs in the process of excretion. Increasing or decreasing metabolism or excretion can lead to a respective decrease or increase of xenobiotic half-life, thus altering disposition. Expression of phase I [e.g., cytochromes P450 (P450s1)] and phase II enzymes [e.g., glucuronosyltransferases and sulfotransferases (STs)] is critical to metabolic processes, whereas transport proteins are vital for excretion. Interestingly, some enzymes and transporters are expressed in a gender-specific manner.
Metabolic enzymes CYP2C11 and CYP2C12 in rat liver are male- and female-specific, respectively (Gustafsson et al., 1983; Waxman et al., 1991). Furthermore, liver phase II STs 1A1 and 20/21 are male- and female-predominant, respectively (Liu and Klaassen, 1996a,b). Additionally, S-adenosylmethionine synthetase is more active in females than in males (Oscarsson et al., 2001). Select transport proteins in kidney, including organic anion transporting polypeptide 1 (oatp1), organic cation transporter 2, and organic anion transporters (Oats) also exhibit gender-divergent expression (Lu et al., 1996; Urakami et al., 1999; Kobayashi et al., 2002; Buist et al., 2002; Slitt et al., 2002). Moreover, male rat kidneys transportpara-aminohippurate (PAH), an anion commonly used to study active transport in kidney, at greater rates than female rat kidneys, both in vivo and in vitro (Kleinman et al., 1966).
Kleinman et al. (1966) and Bräunlich et al. (1993), respectively, determined that testosterone elevated PAH transport by male kidney, and estrogen had no role in gender-divergent PAH transport. Testosterone was also shown to increase expression of oatp1 in kidney (Lu et al., 1996). Conversely, growth hormone (GH) secretion patterns were responsible for the gender-divergent expression of CYP2C11, CYP2C12,S-adenosylmethionine synthetase, ST1A1, and ST20/21 (Waxman et al., 1991; Liu and Klaassen, 1996a,b; Oscarsson et al., 2001).
As mentioned above, gender differences have also been identified in the mRNA levels of three rat Oat family members. Specifically, Oat1 mRNA levels were higher in male compared with female kidney (Buist et al., 2002; Kudo et al., 2002). In addition, Oat2 mRNA was much higher in kidneys of females than those of males (Buist et al., 2002; Kudo et al., 2002), and castration led to a slight increase in Oat2 mRNA in male kidney (Kobayashi et al., 2002; Kudo et al., 2002). Furthermore, hypophysectomy (HX) decreased Oat2 mRNA levels in female rat kidney (Kobayashi et al., 2002). However, the experimental design did not consider HX males or GH replacement to HX rats. Finally, Oat3 mRNA in liver was considerably higher in males than in females (Buist et al., 2002; Kobayashi et al., 2002). After castration, Oat3 mRNA in liver of male rats decreased, and testosterone treatment restored normal male levels (Kobayashi et al., 2002). However, Oat3 mRNA levels were not determined in livers of ovariectomized (OX) females. Moreover, HX increased Oat3 mRNA in female liver, but effects of GH replacement or HX of male rats were not determined (Kobayashi et al., 2002).
Although there are some data addressing the cause of gender differences in Oat mRNA levels, the results are varied and incomplete. Therefore, the goal of this study was to determine whether sex steroids and/or GH secretion patterns are important factors in the gender-divergent expression of Oat1 and Oat2 mRNA in kidney, and Oat3 mRNA in liver. Data from this study complement the previous results of gonadectomy (Kobayashi et al., 2002; Kudo et al., 2002), support evidence of sex steroid regulation of Oat1, and suggest GH regulation of Oat2 and Oat3.
Materials and Methods
Materials.
Sodium chloride, HEPES sodium salt, HEPES free acid, lithium lauryl sulfate, EDTA, and d-(+)-glucose were obtained from Sigma-Aldrich (St. Louis, MO). Micr-O-protect and blocking agent were purchased from Roche Diagnostics (Indianapolis, IN). Formaldehyde, MOPS, sodium citrate, and NaHCO3 were purchased from Fischer Scientific Co. (Fair Lawn, NJ). Chloroform, agarose, and ethidium bromide were purchased from Amresco (Solon, OH). Recombinant rat GH was procured from the National Institute of Diabetes and Digestive and Kidney Diseases through the National Hormone and Pituitary Program (UCLA, Torrence, CA).
Animals.
Sasco Sprague-Dawley rats were purchased from Charles River Laboratories, Inc. (Wilmington, MA). Rats were gonadectomized at 25 days of age or HX at 30 days of age by Charles River Laboratories, Inc. Animals were maintained on automatically timed 12-h dark/light cycles in an American Animal Associations Laboratory Animal Care-accredited facility and allowed water and rat chow ad libitum (Teklad; Harlan, Indianapolis, IN); HX rats received 5% glucose water (w/v) ad libitum. HX rats that gained more than 10 g per week before the start of the study were excluded under the assumption that their surgery was incomplete. Gonadectomized rats were terminated at 62 days of age along with age-matched intact controls (n = 5/gender). HX rats were terminated at 45 to 47 days of age, also with age-matched intact controls. After termination, livers and kidneys were snap-frozen in liquid nitrogen and stored at −80°C.
Treatments.
HX male and female rats were separated into five treatment groups (n = 5–6/gender/treatment): 1) untreated control; 2) GH vehicle injected (0.15 M NaCl, 0.01 M NaHCO3; 10 ml/kg body weight) every 12 h; 3) GH injected (115 μg GH/kg body weight; 11.5 μg GH/ml) every 12 h; 4) GH vehicle continuously infused (1 μl/h); and 5) GH continuously infused (5 μg of GH/h). Intact, untreated, age-matched rats were used as controls. All treatments were administered subcutaneously beginning at 40 days of age and were sustained over 5 to 6 days. Continuous infusion (1.0 μl/h) of vehicle and GH was achieved via Alzet mini-osmotic pumps (model 2001; Durect Corp., Cupertino, CA). Pumps were filled and subcutaneously implanted, under nembutal anesthesia (40 mg/kg), according to protocols provided by the manufacturer. GH treatments were based on previously published protocols (Waxman et al., 1991; Liu and Klaassen 1996a,b). Waxman et al. (1991) demonstrated that although 12-h subcutaneous injections of GH did not produce a physiological GH plasma profile, they effectively stimulated male-specific CYP2C11-dependent steroid hydroxylase activity in HX males and females.
Total RNA Isolation.
Total RNA was isolated using RNAzol B reagent (Tel-Test Inc., Friendswood, TX) according to the manufacturer's protocol. RNA pellets were resuspended in diethyl pyrocarbonate-treated deionized water. Total RNA concentrations were quantified spectrophotometrically at 260 nm. Integrity of RNA samples was analyzed by formaldehyde-agarose gel electrophoresis (1.2% agarose, 2.1 M formaldehyde in 1× MOPS, 0.5 μg ethidium bromide/mg sample) with visualization under ultraviolet light by ethidium bromide fluorescence.
Development of Specific Oligonucleotide Probe Sets for Branched DNA (bDNA) Analysis.
Gene sequences of interest were accessed from GenBank (CYP2C11 and CYP2C12, Table 1; Oat1, Oat2, and Oat3,Buist et al., 2002). Target sequences were analyzed with ProbeDesigner software, version 1.0 (Bayer Corp., Emeryville, CA) for suitability as capture, label, or blocker probes. Multiple specific probes were developed to each P450 and Oat mRNA transcript (Table 1; Buist et al., 2002). All oligonucleotide probes were designed with a melting temperature of approximately 63°C. This enabled stringent hybridization conditions to be held constant (i.e., 53°C) during each hybridization step for each oligonucleotide probe set. Each probe designed in ProbeDesigner was submitted to the National Center for Biotechnological Information for nucleotide comparison by the basic local alignment search tool (BLASTn; Altschul et al., 1997) to ensure minimal cross-reactivity with other rat sequences. Oligonucleotides with a high degree of similarity to other rat gene transcripts were eliminated from the design. Together, the high melting temperature and sequence specificity to the gene of interest ensure detection of the specific mRNA of interest. Probes were synthesized by Operon Technologies (Almeda, CA).
List of oligonucleotide probes generated for analysis of CYP2C expression by bDNA signal amplification
bDNA Assay.
Reagents required for RNA analysis (i.e., lysis buffer, amplifier/label probe buffer, and substrate solution) were supplied in the HV-Quantigene bDNA signal amplification kit (Bayer Corp., Diagnostics Division, Tarrytown, NY). Oat1, Oat2, and Oat3 mRNA levels were analyzed according to the method of Hartley and Klaassen (2000). Briefly, specific rat cytochromes P450 (Table 1) or Oat oligonucleotide probes (previously described by Buist et al., 2002) were diluted in lysis buffer. Total RNA (1 μg/μl; 10 μl) was added to each well of a 96-well plate containing 50 μl of capture hybridization buffer [0.05 M HEPES sodium salt, 0.05 M HEPES free acid, 0.037 M lithium lauryl sulfate, 0.5% (v/v) Micr-O-protect, 8 mM EDTA, 0.3% (w/v) nucleic acid blocking agent] and 50 μl of diluted probe set. Total RNA was allowed to hybridize to probe sets overnight at 53°C. Subsequent hybridization steps were carried out according to the manufacturer's protocol, and luminescence was measured with a Quantiplex 320 bDNA luminometer interfaced with Quantiplex Data Management software, version 5.02, for analysis of luminescence from 96-well plates. Data are presented as relative light units (RLUs) per 10 μg of total RNA.
Statistical Analysis.
Data were analyzed between gender for HX and GH replacement groups by Student's t test (p ≤ 0.05). No significant differences were observed, so data were averaged together by treatment and represented as individual bars by treatment only. Averaged treatment values were then subjected to statistical analysis by analysis of variance followed by Duncan's multiple range post hoc test to identify differences between treatment groups. Asterisks (*) indicate a statistical difference (p ≤ 0.05) between intact female as compared with intact male; single daggers (†) indicate a statistical difference (p ≤ 0.05) compared with untreated HX control. Bars represent mean ± S.E.M.
Results
Kidney and liver samples from castrated and OX rats were analyzed for Oat mRNA levels and compared with age-matched, intact controls. The Oat1 mRNA level in control male kidney was higher than in control female kidney (Fig. 1, top). After castration, the level of Oat1 mRNA in male kidneys decreased to a level equivalent to control females. However, female kidney levels of Oat1 mRNA were unaffected by OX. In contrast, Oat2 mRNA in kidney, where female levels were notably higher than males, was unaffected by gonadectomy (Fig. 1, middle). Finally, the higher Oat3 mRNA level observed in male liver compared with female liver was reduced by castration of males (Fig. 1, bottom). Additionally, as with Oat1, OX had no effect on the level of Oat3 mRNA in female liver.
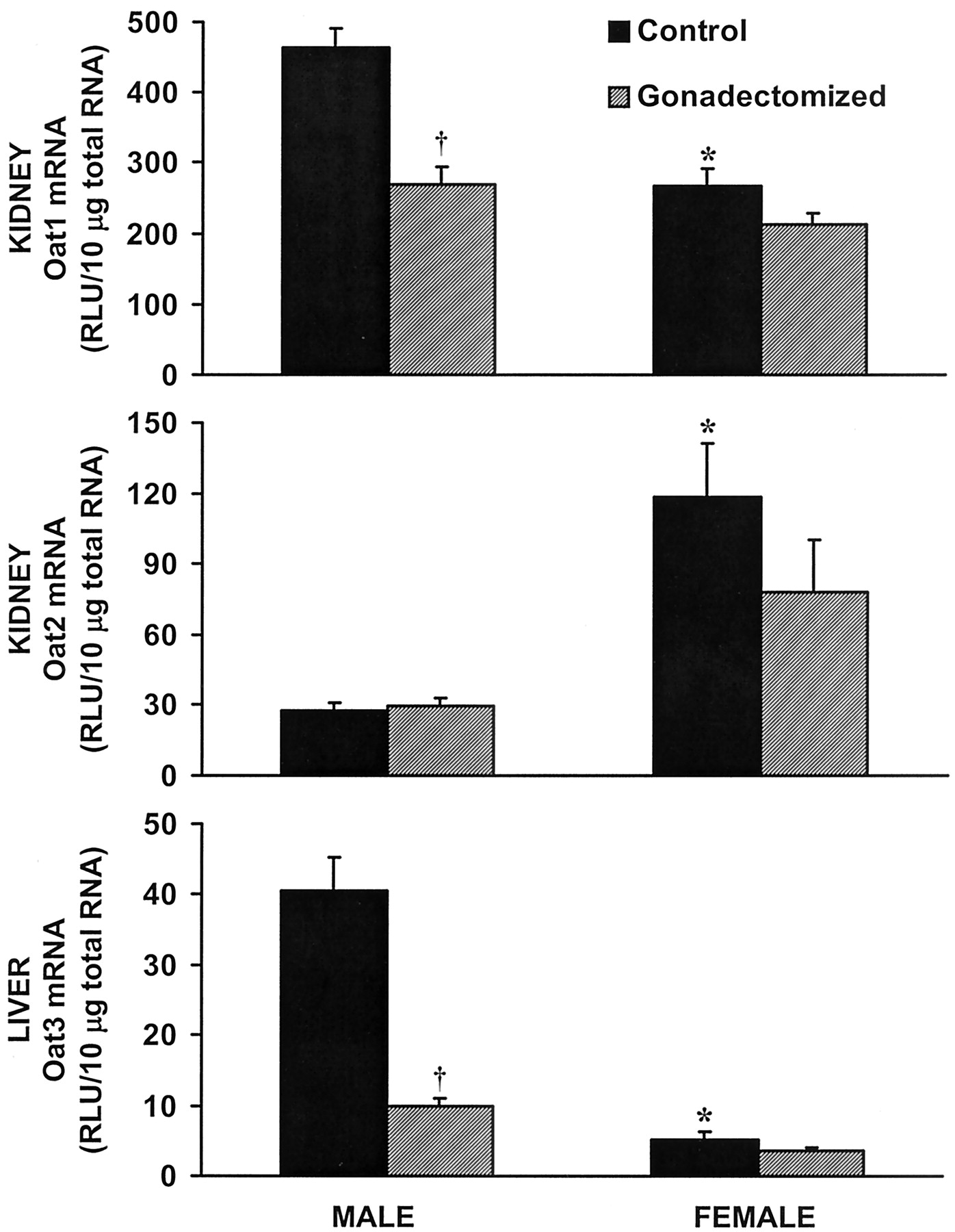
Effects of gonadectomy on Oat mRNA in kidney and liver.
Relative quantification of Oat mRNA by bDNA. Top, Oat1 mRNA levels in kidney. Middle, Oat2 mRNA levels in kidney. Bottom, Oat3 mRNA levels in liver. ∗, p ≤ 0.05 compared with intact male; †, p ≤ 0.05 compared with intact control. Bars represent RLU ± S.E.M., n = 5.
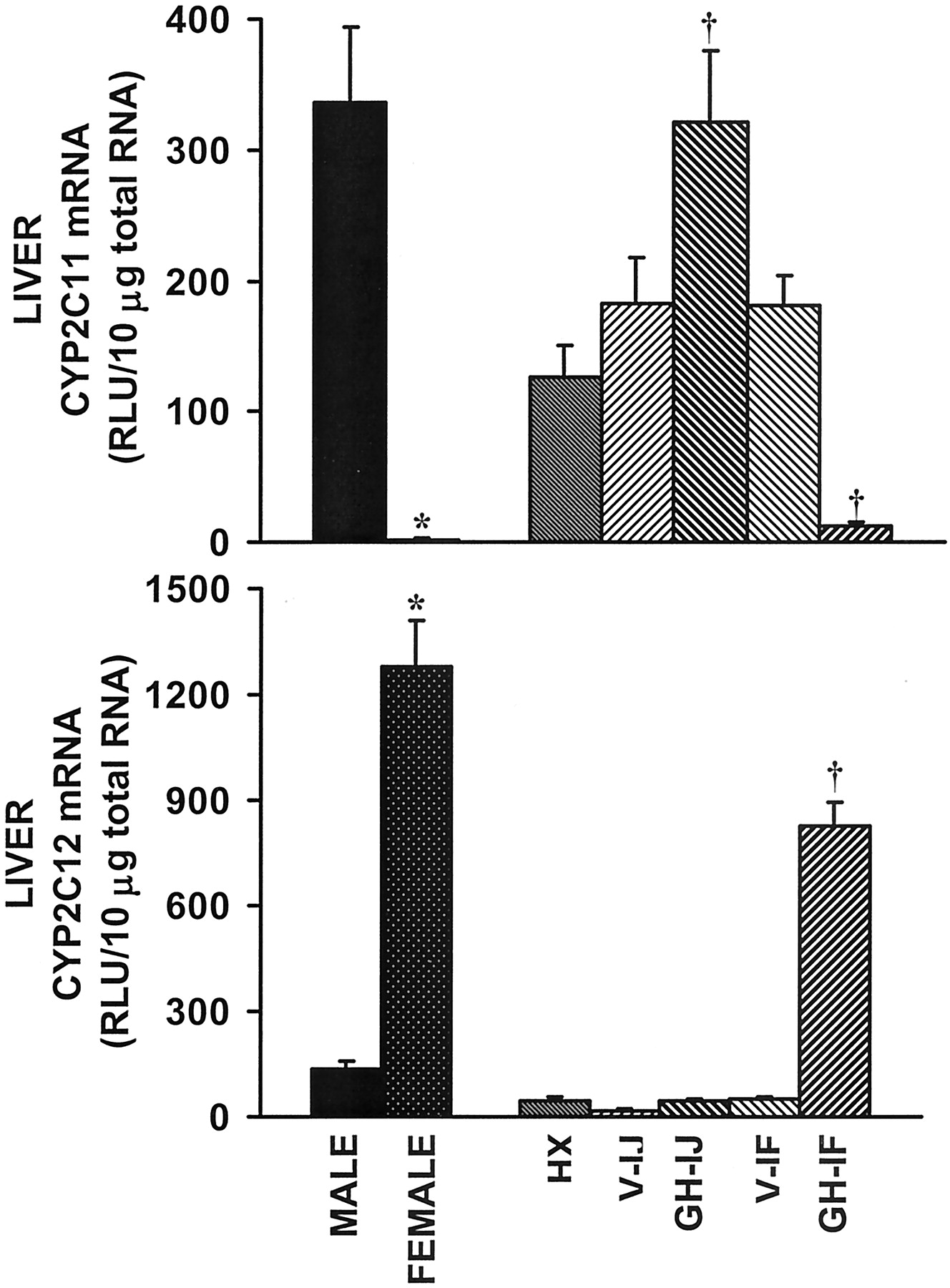
In addition to sex steroids, GH secretion patterns are pivotal factors in gender-specific expression of various genes (Gustafsson et al., 1983; Waxman et al., 1991; Liu and Klaassen 1996a,b; Oscarsson et al., 2001). Therefore, the influence of GH was investigated by comparing intact and HX rats, with HX rats that received GH replacement by either injection (male secretion pattern) or continuous infusion (female secretion pattern). Efficacy of GH replacement patterns was determined by measuring CYP2C11 and CYP2C12 mRNA levels in liver. CYP2C11 was high in intact male liver and nearly undetectable in female liver (Fig.2, top). After HX, CYP2C11 mRNA decreased in male and increased in female liver. Administration of growth hormone by injection increased CYP2C11 mRNA in HX rats to a level comparable with intact males, indicating that the GH injection regimen was effective. Additionally, GH infusion suppressed CYP2C11 mRNA levels, suggesting appropriate feminization of the liver by this treatment regimen. The treatments were further validated by observation of a dramatic increase in CYP2C12 mRNA after GH infusion to HX rats, as well as low levels of CYP2C12 mRNA after GH injection (Fig. 2, bottom).

Effects of HX and GH treatments on CYP2C11 and CYP2C12 mRNA in liver.
Relative quantification of CYP2C11 and 2C12 mRNA by bDNA. Male and female values were similar after HX so single bars represent average values of male and female rats per treatment. Top, CYP2C11 mRNA levels in liver. Bottom, CYP2C12 mRNA levels in liver. V-IJ, vehicle administered by injection; GH-IJ, growth hormone administered by injection; V-IF, vehicle administered by continuous infusion; GH-IF, growth hormone administered by continuous infusion. ∗,p ≤ 0.05 intact female compared with intact male; †, p ≤ 0.05 compared with untreated HX control. Bars represents mean RLU ± S.E.M.; intact controls,n = 5 to 6; HX and treated, n = 8 to 20.
As previously observed, control kidney Oat1 mRNA levels were higher in males than in females. HX caused male kidney levels of Oat1 mRNA to decrease to a level similar to that detected in intact control female kidney (Fig. 3, top). GH replacement, by injection or infusion, was ineffective in restoring normal physiological Oat1 mRNA levels in HX males. Additionally, Oat1 mRNA levels in female rats were unaffected by HX or either GH replacement profile.

Effects of HX and GH treatments on Oat mRNA in kidney and liver.
Relative quantification of Oat mRNA by bDNA. Male and female values were similar after HX, so single bars represent average values of male and female rats per treatment. Top, Oat1 mRNA levels in kidney. Middle, Oat2 mRNA levels in kidney. Bottom, Oat3 mRNA levels in liver. V-IJ, vehicle administered by injection; GH-IJ, growth hormone administered by injection; V-IF, vehicle administered by continuous infusion; GH-IF, growth hormone administered by continuous infusion. ∗,p ≤ 0.05 intact female compared with intact male; †, p ≤ 0.05 compared with untreated HX control. Bars represents mean RLU ± S.E.M.; intact controls,n = 5 to 6; HX and treated, n = 8 to 20.
Oat2 mRNA levels were significantly higher in control female kidney compared with control male kidney. Male kidney levels were not significantly altered after HX, whereas female levels were markedly decreased after HX (Fig. 3, middle). Furthermore, GH injections, simulating the male secretion pattern, did not further alter Oat2 mRNA levels in HX males or females compared with untreated HX rats. However, GH administered by continuous infusion, simulating the female secretion pattern, significantly increased Oat2 mRNA levels in both male and female HX rats compared with untreated HX controls.
Oat3 mRNA levels were higher in control male liver compared with control female liver. HX decreased Oat3 mRNA levels in male liver and increased Oat3 mRNA in female liver compared with intact controls (Fig.3, bottom). GH injections did not further significantly affect the level of Oat3 mRNA in HX male or female liver. However, administration of GH by infusion decreased Oat3 mRNA levels in liver of both HX males and females compared with untreated HX controls.
Discussion
The primary goal of this study was to identify the roles of gender-divergent hormone systems in the regulation of Oat mRNA levels. GH secretion and sex steroids appear to be the two primary systems responsible for male- (Oat1 in kidney and Oat3 in liver) and female-predominant (Oat2 in kidney) expression patterns. More specifically, androgens increase the level of Oat1 and Oat3 mRNA in male rat kidney and liver, respectively. Additionally, the female GH secretion pattern is at least partially responsible for the elevation of Oat2 mRNA in female kidney as compared with male kidney. The female GH secretion pattern also suppresses Oat3 mRNA in female liver, thus accentuating the gender difference in Oat3 mRNA expression.
Although the data presented here in combination with previous studies (Kobayashi et al., 2002; Kudo et al., 2002) provide evidence of androgen and GH regulation of rat Oat mRNA expression, there are complications in properly deciphering and fully understanding the data to date. The major difficulty lies in the complicated nature of hormonal interactions. HX provides a nice method of GH removal from the physiology of an in vivo system. However, removal of the pituitary eliminates other critical hormones, including thyroid-stimulating hormone, leuteinizing hormone, and follicle-stimulating hormone. Likewise, castration of male or female rats disrupts more than just the levels of androgens and estrogens. For example, castrated male rats exhibited lower amplitude GH pulses of shorter duration and less regularity than those of intact males (Jansson and Frohman, 1987). Moreover, administration of testosterone during adulthood to neonatally castrated male rats restored GH pulse frequency, height, and mean GH concentration to normal levels (Jansson and Frohman, 1987). Furthermore, estrogen administration to intact male rats resulted in elevated plasma GH trough levels and decreased pulse amplitudes, in effect feminizing the GH plasma profile (Mode et al., 1982). Therefore, although this study provides insight into the hormonal regulation of Oat mRNA expression, it is important to recognize the complicated nature of hormonal interactions.
Despite the complications of hormonal interactions, the present data may provide some insight into data regarding gender differences in the clearance of PAH and perfluorooctanoic acid (PFOA). Male rat kidneys reportedly accumulate PAH at a greater rate than female rat kidneys (Huang and McIntosh, 1955; Kleinman et al., 1966; Bowman and Hook, 1972). Gonadectomy of male rats reduced kidney PAH transport, which was restored by testosterone administration (Kleinman et al., 1966;Bräunlich et al., 1993). However, PAH transport was not affected by ovariectomy of female rats or estradiol administration to male rats (Kleinman et al., 1966; Bräunlich et al., 1993). Thus, estrogens did not influence the organic anion transport processes of the kidney. Transport of PAH is mediated primarily by a tertiary active process, which directly corresponds to the mechanism of Oat1 (Sekine et al., 1997). In agreement with previously observed effects of sex steroids on PAH transport, the present study demonstrated that rat Oat1 mRNA levels were higher in male than female kidney (Fig. 1, top) and decreased after castration of male rats. Another study demonstrated that testosterone administered to intact female rats increased the level of Oat1 mRNA in kidney (Kudo et al., 2002). Moreover, Cerrutti et al. (2001) showed that male rats had a higher number of functional carriers in the basolateral membrane of the proximal tubule than did females. Together, these data suggest that androgens increase Oat1 mRNA in male kidney compared with female kidney and provide molecular support for the previously described difference in PAH transport.
Although primarily androgens appear to be responsible for the gender difference in Oat1 mRNA levels, the influence of GH was investigated considering the influence sex steroids have on GH plasma profiles, as previously discussed. Moreover, HX was previously shown to decrease renal PAH transport (Farah et al., 1956; Misanko et al., 1977). Farah et al. (1956) further demonstrated that GH, thyroxine, and testosterone each partially restored PAH transport. HX decreased male levels of Oat1 mRNA in kidney without affecting female levels (Fig. 3, top). Furthermore, GH administered by injection or infusion had no significant affect on Oat1 mRNA levels. Because HX reduces sex steroid levels secondarily to the eradication of leuteinizing hormone and follicle-stimulating hormone, these data support the importance of androgens in elevating Oat1 mRNA levels in male kidney.
In contrast to PAH, renal clearance of PFOA is greater in female than male rats (Griffith and Long, 1980; Hanhijärvi et al., 1982; Kudo et al., 2002). Griffith and Long (1980) demonstrated that male rats developed hepatotoxicity at lower PFOA treatment levels than did females, and corresponding serum levels were 75–226 fold greater in males than in females. These differences were attributed to relatively slow urinary excretion in male as compared with female rats, and led to the conclusion that female rats possess an active secretory mechanism which is lacking or inactive in male rats (Hanhijärvi et al., 1982). Additionally, castration of male rats increased renal clearance of PFOA (Kudo et al., 2002). Considering renal clearance of PFOA is greater in female than male rats, Oat2, which is expressed at higher levels in female than male kidney, may be involved in PFOA excretion.
Both Kudo et al. (2002) and Kobayashi et al. (2002) report an increase in Oat2 mRNA levels in male kidney after castration. However, this increase is minimal and does not approach the level of expression in female kidney. In contrast, no change was observed in male kidney after castration in the present study (Fig. 1, middle). Additionally,Kobayashi et al. (2002) and Kudo et al. (2002) demonstrated that a small decrease in Oat2 mRNA levels occurs after testosterone administration to castrated males. However, Kobayashi et al. (2002)reported that estrogen administration to castrated males decreased Oat2 mRNA levels in kidney, whereas Kudo et al. (2002) described a slight increase in Oat2 mRNA levels in male kidney after estrogen administration. These variable results suggest that sex steroids are not the primary mediators of this gender difference. In agreement withKobayashi et al. (2002), HX markedly decreased Oat2 mRNA in female rat kidney (Fig. 3, middle). Furthermore, Oat2 mRNA levels were unaffected by HX in male rats. Continuous infusion of GH (female pattern) significantly increased Oat2 mRNA levels in HX rats compared with untreated controls (Fig. 3, middle). Although the level of Oat2 mRNA did not reach the physiological level of intact female rat kidney, clearly a continuous baseline of GH is important in producing the higher level of rat Oat2 mRNA in female compared with male kidney. Furthermore, administration of estrogen to intact males increased Oat2 mRNA levels (Kudo et al., 2002). Together with data that demonstrated feminization of plasma GH profiles after estrogen administration to intact males (Mode et al., 1982), these data support the conclusion that the female pattern of GH secretion is at least partially responsible for the elevated level of Oat2 mRNA in female kidney compared with male kidney. Furthermore, based on the response of Oat2 to HX, GH infusion and estrogen, together with data regarding PFOA clearance, this transporter may be further investigated as a potential member in the mechanism of PFOA clearance.
Although Oat3 mRNA is primarily expressed in kidney, a significant gender difference (male > female) was identified in liver (Buist et al., 2002; Kobayashi et al., 2002). Therefore, the potential influences of sex steroids and GH on Oat3 mRNA levels in liver were investigated. In agreement with Kobayashi et al. (2002), the present study demonstrated a decrease in Oat3 mRNA levels in male liver after castration. Additionally, testosterone administration to castrated males restored normal levels of Oat3 mRNA in male liver, whereas estrogen had no effect (Kobayashi et al., 2002). Oat3 mRNA levels in female liver were not responsive to ovariectomy (Fig. 1, bottom). As with CYP2C11, HX decreased male and increased female Oat3 mRNA levels in liver (Fig. 3, bottom). Administration of GH injections to HX rats did not significantly affect levels of Oat3 mRNA in liver of HX rats, but continuous infusion decreased hepatic levels. Considered with the data after gonadectomy, these data suggest dual regulation of Oat3 in which androgens increase and the female GH secretion pattern decreases the level of Oat3 mRNA in male and female rat liver, respectively. However, this does not rule out the possibility that other pituitary factors not replaced after HX in this study are involved in the elevation of Oat3 mRNA in male compared with female rat liver.
Gender differences in rodents are abundant but do not always translate into parallel differences in humans. However, understanding gender divergences in rodents is essential to properly interpret preclinical clearance and toxicology data. Although these differences may not be observed in humans, thorough characterization is critical to determining the most appropriate models for predicting human responses to certain classes of drugs, such as organic anions.
In conclusion, three distinct gender differences exist for mRNA levels of Oats. Oat1 mRNA levels are higher in male kidney than female kidney, Oat2 mRNA is present at markedly higher levels in female kidney than male kidney, and Oat3 mRNA is more abundant in male liver than in female liver. Furthermore, there seem to be distinct mechanisms mediating each gender-specific expression pattern as demonstrated by the various responses to gonadectomy, HX, and GH treatments. Androgens increased Oat1 mRNA in kidney, whereas continuous secretion of GH increased Oat2 mRNA in kidney. Both androgens and the female GH secretion pattern regulate Oat3 mRNA levels in liver, with androgens increasing and continuous GH secretion decreasing mRNA levels. Moreover, these differences may become important in explaining gender-specific discrepancies in drug disposition, particularly those related to excretion.
Acknowledgments
We thank Melinda Shelby, Jon Maher, Xingguo Cheng, and Drs. Angela Slitt and Tyra Leazer for technical assistance.
Footnotes
-
This study was supported by National Institutes of Health Grants ES-09649, ES-09716, and ES-07079.
- Abbreviations used are::
- P450
- cytochrome P450
- ST
- sulfotransferase
- Oatp1
- organic anion transporting polypeptide 1
- Oat
- organic anion transporter
- PAH
- para-aminohippurate
- GH
- growth hormone
- HX
- hypophysectomy or hypophysectomized
- OX
- ovariectomy or ovariectomized
- MOPS
- 3-(N-morpholino)propanesulfonic acid
- bDNA
- branched DNA
- PFOA
- perfluorooctanoic acid
- Received August 5, 2002.
- Accepted January 27, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}