


Abstract
MaxiPost [(3S)-(+)-(5-chloro-2-methoxyphenyl)-1,3-dihydro-3-fluoro-6-(trifluoromethyl)-2H-indole-2-one), or BMS-204352)] is a potent and specific maxi-K channel opener for potential use to treat stroke. This article describes structural characterization of a major human N-glucuronide metabolite of BMS-204352 and identification of the enzyme responsible for the N-glucuronidation reaction. After intravenous administrations of [14C]BMS-204352 (10 mg, 50 μCi) to eight healthy human subjects, one major metabolite M representing an average of 17% of the radioactive dose was excreted in pooled urine collected over 0 to 336 h after dosing. A major biliary metabolite of dogs dosed with [14C]BMS-204352 (5 mg/kg), which represented about 33% of the dose, has the same retention time and the same tandem mass spectrometry fragmentation pattern as the human urinary metabolite M. Four hundred fifty micrograms of the metabolite was isolated from the dog bile and analyzed by NMR. Long-range 1H-13C NMR experimentation indicated that the glucuronic acid moiety was at the nitrogen site. The N-glucuronide of BMS-204352 was stable up to 24 h at 37°C in the incubations at different pH values (3.0, 7.4, and 9.0) and with glucuronidases from Escherichia coli and Helix pomatia. Of the seven human UDP-glucuronosyltransferases (UGT) isozymes (1A1, 1A3, 1A4, 1A6, 1A7, 1A10, and 2B7) tested, only UGT2B7 produced metabolite M. UGT2B7-catalyzed N-glucuronidation of BMS-204352 exhibited Michaelis-Menten kinetics with a Km of 14.2 μM and Vmax of 0.29 nmol/min · mg of protein. Collectively, these results suggest that amide N-glucuronidation, a major elimination pathway of MaxiPost, is catalyzed by UGT2B7 in humans. This N-glucuronide represents a fully characterized amide N-glucuronide, and glucuronidation at the nitrogen represents a newly identified conjugation reaction for UGT2B7.
Glucuronidation represents a major biotransformation pathway that increases the elimination of many lipophilic drugs and endogenous compounds. UDP-glucuronosyltransferases (UGTs1) are a superfamily of enzymes residing in the endoplasmic reticulum and nuclear membrane of cells and catalyze glucuronidation at a nucleophilic functional group of oxygen (hydroxyl and carboxylic acid groups), nitrogen (amines and amides), and sulfur (thiols) with the formation of β-d-glucuronide conjugates (Miners and Mackenzie, 1991; King et al., 2000). There are two subfamilies of UDP-glucuronosyltransferases for xenobiotic metabolism in humans, UGT1A and UGT2B (Mackenzie et al., 1997). UGT2B7 is a very important hepatic enzyme for drug metabolism (Jin et al., 1993) and also exists in extrahepatic tissues including brain (Howe et al., 1992; King et al., 2000). UGT2B7 catalyzes the glucuronidation of the neurotransmitter serotonin (King et al., 1999) and many opioid drugs including the conversion of morphine to its more active morphine 6-glucuronide metabolite. A recent inhibition study and NMR analysis of an aglycone active site-fusion protein suggest that UGT2B7 has one binding site for aglycone binding (Coffman et al., 2001; Rios and Tephly, 2002). A recent search of literature failed to reveal any references for formation of N-glucuronides by UGT2B7.
MaxiPost (BMS-204352), chemically designated as (3S)-(+)-(5-chloro-2-methoxyphenyl)-1,3-dihydro-3-fluoro-6-(trifluoromethyl)-2H-indole-2-one) (Fig. 1), is a maxi-K channel opener and has the potential to prevent and treat ischemic stroke (Gribkoff et al., 2001). Stroke is a major cause of death and long-term disability, affecting more than 700,000 people in the United States annually (Williams et al., 1999). Acute ischemic stroke is the most common form, producing pathologically fatal levels of intracellular calcium (Ca2+) in neurons at risk. Maxi-K channels are large-conductance voltage- and Ca2+-activated K+ channel proteins (Chang et al., 1997). The fluorooxyindole BMS-204352 is a potent maxi-K channel opener and provided significant levels of cortical neuroprotection in rat models of stroke by augmenting an endogenous mechanism for regulating Ca2+ entry and membrane potential to protect the neurons (Cheney et al., 2001; Gribkoff et al., 2001). Positron emission tomography imaging showed that 18F-labeled BMS-204352 is rapidly and widely distributed in rats; the brain uptake was rapid and occurred at high levels (Kiesewetter et al., 2002). The compound had a brain/plasma exposure ratio of 9:1 following an intravenous bolus dose of 5 mg/kg in rats (Hewawasam et al., 2002). MaxiPost showed extensive metabolism in rats and dogs (Krishna et al., 2002) and exhibited protein covalent binding in plasma of rats dosed with the 14C-labeled drug (Zhang et al., 2003a).
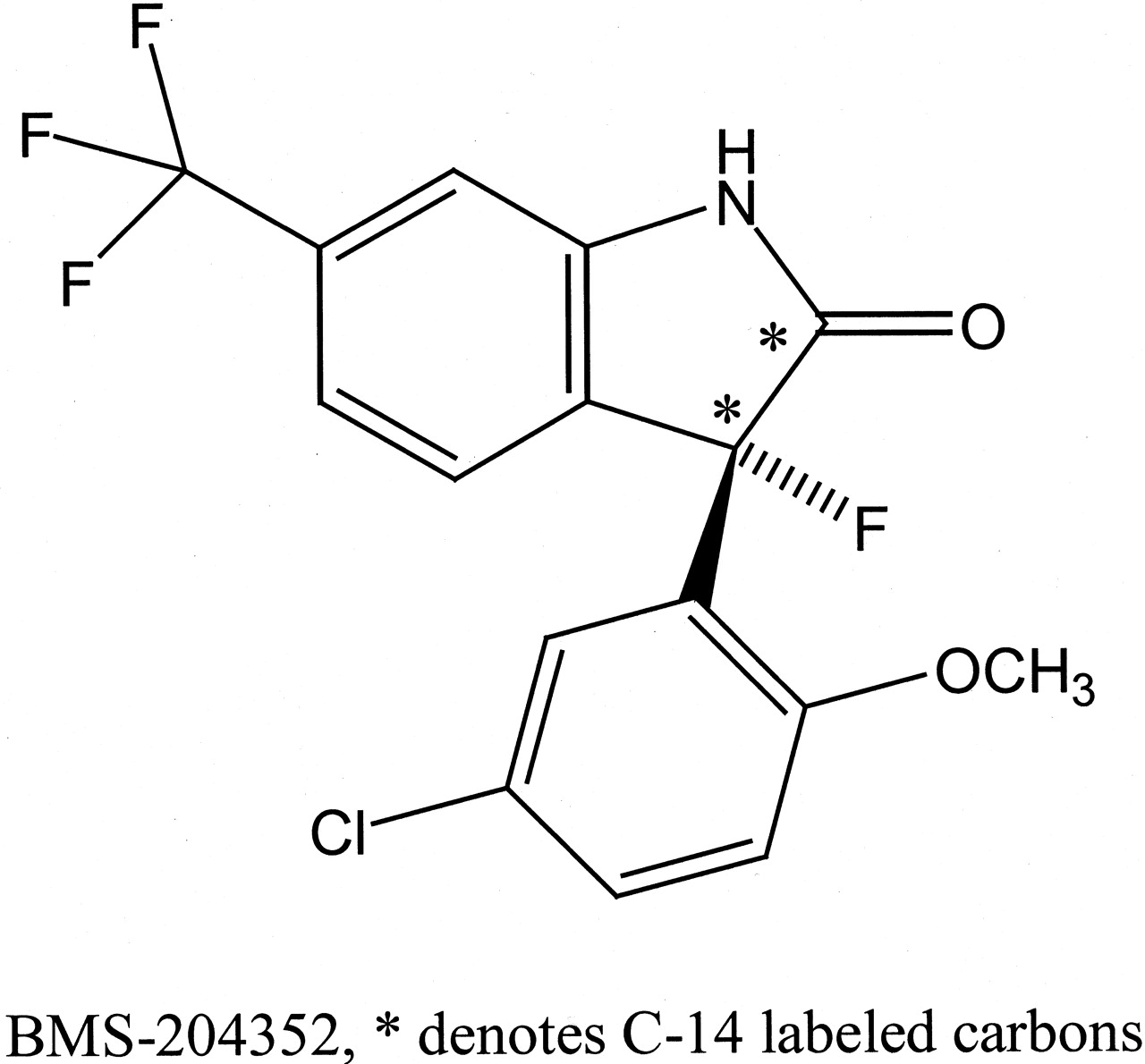
Structure of MaxiPost (BMS-204352).
In this study, N-glucuronidation was characterized as a major metabolic pathway of MaxiPost (BMS-204352) in humans by radioactive metabolite profiling, LC/MS, HPLC isolation, and NMR analyses. Furthermore, the enzyme that is responsible for formation of the N-glucuronide was characterized by using human cDNA-expressed UGTs and by enzyme kinetic analyses.
Materials and Methods
Materials. [14C]BMS-204352 (radiochemical purity 99.5%) (Fig. 1) was synthesized at Bristol-Myers Squibb Co. (Stamford, CT) (Dischino et al., 2003). Ammonium acetate, Tris-HCl, UDPGA, phenolphthalein glucuronic acid, glycine, and glucuronidase (type H-1 from Helix pomatia, 486,000 units/g, type VII-A from Escherichia coli, 1000 units per vial) were purchased from Sigma-Aldrich (St. Louis, MO). Ecolite liquid scintillation cocktail was purchased from MP Biomedicals (Irvine, CA). Pooled human liver microsomes from 10 subjects and UDP-gluconosyltransferase Supersomes (membrane fractions from insect cells infected with baculovirus-containing human cDNA UGT1A1, 1A3, 1A4, 1A6, 1A10, and 2B7) were purchased from BD Gentest (Woburn, MA), and UGT1A7 was purchased from PanVera Corp. (Madison, WI). MgCl2 was obtained from J.T. Baker (Phillipsburg, NJ). Acetonitrile-d3 (D 99.8%) was obtained from Cambridge Isotope Laboratories, Inc. (Andover, MA). All other chemicals used were of reagent grade or better. For solid-phase extraction, Oasis HLB C-18 cartridge columns were obtained from Waters (Milford, MA).
Sample Collections and Preparations. Human urine (at 24-h intervals for 336 h) was collected after single intravenous doses of [14C]BMS-204352 to eight healthy male subjects (10 mg, 50 μCi). A pooled urine sample was obtained by combining 1% of all urine collections from eight subjects over 0 to 336 h after dosing. Dog bile (0–96 h) was obtained from one male beagle dog after intravenous infusion of [14C]BMS-204352 (5 mg/kg, 2 μCi/mg). The pooled human urine (15 ml) was concentrated on a 5-ml C-18 cartridge. The radioactive material was eluted with methanol and concentrated under a stream of nitrogen. The residue was suspended in 0.2 ml of solvent containing water/acetonitrile (30:70, v/v) and centrifuged at 3000 rpm for 10 min before HPLC analysis. Dog bile (10 μl) was directly injected onto the HPLC column after centrifugation at 3000 rpm for 10 min. The dog bile was also coinjected with the human urine.
HPLC. HPLC was performed on a Shimadzu Class VP system equipped with two pumps (model LC-10AT), an autoinjector (SIL 10AD), and a diode array detector (SPD-M10A) (Shimadzu, Kyoto, Japan). A Zorbax RX C-18 column (4.6 mm × 250 mm, 5 μm) (Agilent Technologies, Palo Alto, CA) equipped with a guard column was used. HPLC analyses were performed by enclosing the column in an Eppendorf CH-30 column heater (Eppendorf-5 Prime, Inc., Boulder, CO) maintained at 35°C or at room temperature. The mobile phase flow rate was 1 ml/min. The retention time of the parent compound was confirmed before each run by radioactivity or UV detection. A gradient of two solvent systems, A and B, was used for HPLC analysis. Solvent A was water containing 15 mM ammonium acetate, pH 4.5. Solvent B consisted of methanol/solvent A (95:5, v/v). Two solvent gradients were used. Gradient I was used for in vivo metabolite profiling. In this gradient, solvent B started at 5% and then increased in a linear manner as follows: 45% (15 min), 56% (30 min), 64% (50 min), 80% (70 min), and 90% (75 min). Gradient II was used to analyze in vitro samples. In this gradient, solvent B started at 55% and then increased to 60% (10 min), 90% (30 min), 55% (31 min). Under gradient I conditions, the N-glucuronide of BMS-204352 and BMS-204352 had respective retention times of 37 and 62 min. Under gradient II conditions, the N-glucuronide of BMS-204352 and BMS-204352 had respective retention times of 20.5 and 29.5 min.
Radioactivity Detection. Radioactivity in biological samples was determined by mixing aliquots of the samples with 15 ml of Ecolite cocktail and counting with a Packard Tri-Carb 2250 liquid scintillation analyzer (PerkinElmer Life and Analytical Sciences, Boston, MA). In addition, HPLC effluent was collected at 15-s intervals after sample injection into 96-deep-well Lumaplates with a Gilson model 202 fraction collector (Gilson Medical Electronics, Middleton, WI). The effluent in the plates was dried with a SpeedVac (Savant Instruments, Holbrook, NY), and the plates were counted for 10 min per well with a TopCount scintillation analyzer (PerkinElmer Life and Analytical Sciences). Radiochromatographic profiles were prepared by plotting the cpm values in HPLC fractions against time-after-injection.
LC/MS. LC/MS analysis was performed on a Thermo Finnigan LCQ mass spectrometer with an electrospray ionization probe (Thermo Finnigan, San Jose, CA). Samples were analyzed in the negative ion mode. Half of the HPLC eluent was directed to the Thermo Finnigan LCQ mass spectrometer through a divert valve set to divert the flow from 0 to 10 min. From 10 min until the end of the HPLC run, the eluent flow was directed to the mass spectrometer. The capillary temperature was set at 220°C. The nitrogen gas flow rate, spray current, and voltages were adjusted to give a maximum sensitivity. The other half of the HPLC eluent was directed to a fraction collector.
NMR. NMR experiments were performed on a Bruker Avance 600 MHz spectrometer (Bruker, Newark, DE) equipped with a 3-mm triple resonance Nalorac gradient probe and operating at 600.07 MHz. One-dimensional 1H, two-dimensional correlation spectroscopy (proton-proton coupling), and two-dimensional 1H-13C heteronuclear multiple quantum coherence and heteronuclear multiple-bond correlation experiments were performed. All experiments were recorded at room temperature. The chemical shifts are reported in ppm relative to tetramethylsilane.
Isolation of N-Glucuronide of BMS-204352 from Dog Bile. Three 15-ml portions of 0- to 96-h collections of dog bile were each acidified with formic acid to 0.25% (v/v), mixed with 30 ml of ethyl acetate, vortexed, and centrifuged for 10 min. The resulting supernatant was removed and saved. The extraction procedure was repeated, and the supernatants from all three portions were combined. The combined supernatant, containing 77% of the radioactivity in the bile, was evaporated to dryness under a stream of nitrogen. The residue was dissolved in 2 ml of a buffer containing 30% A and 70% B and subjected to a two-step HPLC purification procedure.
A Zorbax SB C-18 column (9.4 mm × 250 mm, 5 μm) (Agilent Technologies) equipped with a guard column was used. The mobile-phase flow rate was 4 ml/min. In the first HPLC, two solvents A and B were the same as the ones used in the analytic HPLC. The gradient was as follows: solvent B was started at 30%, increased in a linear manner to 60% (30 min) and 80% (35 min). Under these conditions, the major metabolite was eluted at a retention time of 16 min. About 80% of the central peak was collected. The fractions from a total of 13 injections were combined, evaporated to dryness under a stream of nitrogen, and redissolved in 0.5 ml of acetonitrile. The reconstituted material was subjected to a second semipreparative HPLC. In the second HPLC, solvent C was water and solvent D consisted of 5% water in acetonitrile. The gradient was as follows: solvent D was started at 30%, maintained at 30% for 5 min, and then increased in a linear manner to 65% (25 min) and 90% (30 min). Under these conditions, one radioactive peak was eluted at a retention time of 18 min. About 80% of the peak at the center was collected. The fractions from a total of four injections were combined and evaporated to dryness under a stream of nitrogen. The residue was redissolved in 0.2 ml of acetonitrile-d3 and analyzed by NMR.
Incubations of N-Glucuronide of BMS-204352 in Buffers and with Glucuronidases. [14C]N-Glucuronide of BMS-204352 (4000 dpm) isolated from the dog bile was separately incubated in 0.5 ml of 100 mM potassium phosphate buffer at pH 3.0, 7.4, and 9.0. The incubations were stored at 37°C for 24 h and terminated by addition of 1 volume of acetonitrile. After centrifugation for 10 min at 3000 rpm, an aliquot of 0.10 ml was analyzed by HPLC. [14C]N-Glucuronide of BMS-204352 (3000 dpm) was separately incubated with β-glucuronidases from H. pomatia (2624 units/ml) and from E. coli (1000 units/ml) in 0.5 ml of 0.16 M sodium acetate buffer, pH 4.8. The incubations were done at 37°C for 24 h and terminated by the addition of 1 volume of acetonitrile. After centrifugation for 10 min at 3000 rpm, 0.10 ml of the solution was analyzed by HPLC and radioactivity counting. The positive control was the incubation with phenolphthalein β-glucuronide (1 mM) for 24 h. After addition of 3 volumes of 0.2 M glycine buffer, pH 10.4, the incubation mixtures with β-glucuronidases from both sources turned to dark red, indicating hydrolysis of the control glucuronide.
Incubations of [14C]BMS-204352 with HLM and Expressed UGTs. A stock solution of [14C]BMS-204352 (14 mM) was prepared in methanol, and aliquots were added to the incubation mixture. [14C]BMS-204352 (140 μM) was incubated with pooled HLM or human cDNA-expressed UGT insect cell microsomes (1 mg/ml), UDPGA (2 mM), alamethicin (50 μg/ml), and 10 mM MgCl2 in 0.25 ml of 50 mM Tris buffer, pH 7.5. The UGT isozymes were 1A1, 1A3, 1A4, 1A6, 1A7, 1A10, and 2B7. Incubations were for 2 h at 37°C and were terminated by addition of 0.25 ml of acetonitrile. A second set of incubations was conducted without alamethicin, which enhances other glucuronidation reactions (Fisher et al., 2000). After centrifugation for 10 min at 3000 rpm, 25 μl of the solution was analyzed by HPLC-radioactivity counting and LC/MS.
Determination of Km and Vmax. The stock solution of [14C]BMS-204352 (14 mM) was diluted with methanol, and aliquots were added to the incubation mixture to final methanol concentrations of less than 1% (v/v) of the incubation mixture. [14C]BMS-204352 at 1, 2.5, 5, 10, 20, 40, and 80 μM was incubated with UGT2B7 insect cell microsomes (in triplicate). Incubation mixtures (200 μl) contained 2 mM UDPGA, 10 mM MgCl2, 50 mM Tris buffer, pH 7.5, 25 μg/ml alamethicin, and 1.0 mg/ml UGT2B7 microsomes. Preincubation of the mixture without UDPGA was for 3 min at 37°C. After incubations for 30 min at 37°C, 200 μl of acetonitrile was added. After centrifugation for 10 min at 3000 rpm, an aliquot of 100 μl was analyzed by HPLC. Similarly, [14C]BMS-204352 at the above concentrations was incubated with HLM (0.4 mg/ml) in the presence of 50 μg/ml alamethicin for 20 min. The samples were analyzed by HPLC-radioactivity detection after termination with acetonitrile and centrifugation. The amounts of the product were calculated based on the percentage of conversion of the starting material. The glucuronidation reaction was linear to an incubation time up to 60 min under the conditions used. The reaction rate versus substrate concentration data were analyzed by nonlinear regression analysis by XLFit in Microsoft Excel (Microsoft, Redmond, WA) using the Michaelis-Menten equation: V = (Vmax · S)/(Km + S).
Results
Metabolite Profiling in Human Urine and Dog Bile. HPLC-radiochromatographic profiles of the pooled human urine (0–336 h, 8 subjects) and the dog bile (0–96 h) are shown in Fig. 2. An average of 37% of the radioactive dose was excreted in the pooled urine. The parent compound (P) accounted for 5% of the radioactivity in the pooled urine. The prominent radioactive metabolite M accounted for 45% of the radioactivity in the human urine, or 17% of the total radioactive dose. Overall, approximately 97% of the radioactive dose was excreted from human subjects during 0- to 336-h collections of urine and feces with 60% of the dose in feces. The dog excreted <5% of the radioactive dose in urine (0–96 h) with the N-glucuronide of BMS-204352 as a major radioactive component. About 66% of the radioactive dose was excreted into bile (0–96 h) in the dog. In the dog bile, the metabolite M accounted for 50% of the radioactivity in the sample.

HPLC-radiochromatographic profiles of pooled human urine (0–336 h, 8 subjects) and dog bile (0–96 h) following intravenous administrations of [14C]BMS-204352.
The samples were analyzed by HPLC under gradient I conditions as described under Materials and Methods.
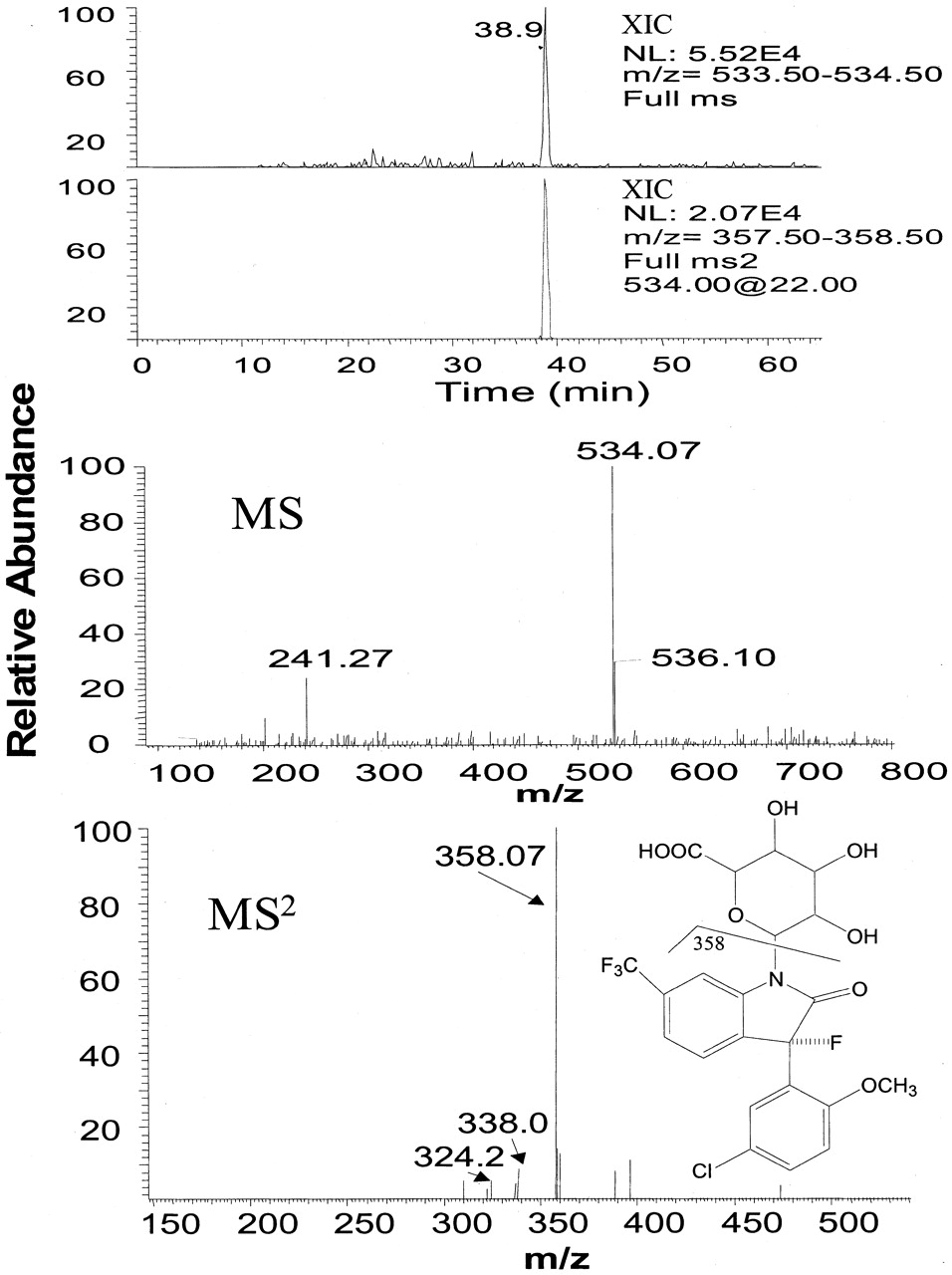
LC/MS analysis showed that human urinary metabolite M had a molecular ion (MH)+ at m/z 534, a major fragment ion at m/z 358 (loss of 176) in MS2 (Fig. 3), and major fragment ions at m/z 338 (MH+-176-HF) and 324 (MH+-176-HF-Me) in MS3 (not shown), indicating a structure consistent with a glucuronide of BMS-204352. In LC/MS analysis of dog bile, the metabolite M had the same molecular ion and the same major fragment ions as the metabolite M in the human urine. In addition, the metabolite M in human urine coeluted with the metabolite M in the dog bile, suggesting that they are the same metabolite.

LC/MS analysis of human urinary N-glucuronide of BMS-204352.
Top two panels, reconstructed ion chromatograms of m/z 534 in MS and m/z 358 in MS2 of m/z 534. Bottom two panels, MS and MS2 spectra of this metabolite. The samples were analyzed by HPLC under gradient I conditions as described under Materials and Methods.
Isolation and NMR Identification of N-Glucuronide of BMS-204352. Due to a low concentration of metabolite M in human urine, it was not feasible to isolate enough for definitive identification of the urinary metabolite by NMR. Therefore, dog bile was collected after an intravenous dose of [14C]BMS-204352 to isolate the metabolite in sufficient quantity for identification (by NMR and mass spectrometry). Metabolite M was purified from dog bile through ethyl acetate extraction and two steps of semipreparative HPLC. The purified material (0.45 mg) was analyzed by one-dimensional and two-dimensional NMR. The protons and carbons were assigned from analyzing one-dimensional 1H, two-dimensional correlation spectroscopy, and two-dimensional 1H-13C heteronuclear multiple quantum coherence and heteronuclear multiple-bond correlation experiments (Fig. 4, Table 1). The key long-range 1H-13C correlation observed (Fig. 5) between the proton at C-1′ and carbons at C-2 and C-5 clearly indicate that the d-glucuronic acid was attached to the nitrogen. Thus, the metabolite M is identified as an amide N-glucuronide of BMS-204352.

1H one-dimensional NMR spectrum of N-glucuronide of BMS-204352.
1H and 13C NMR chemical shift assignments of N-glucuronide of BMS-204352 Atom labels are shown in Fig. 4.
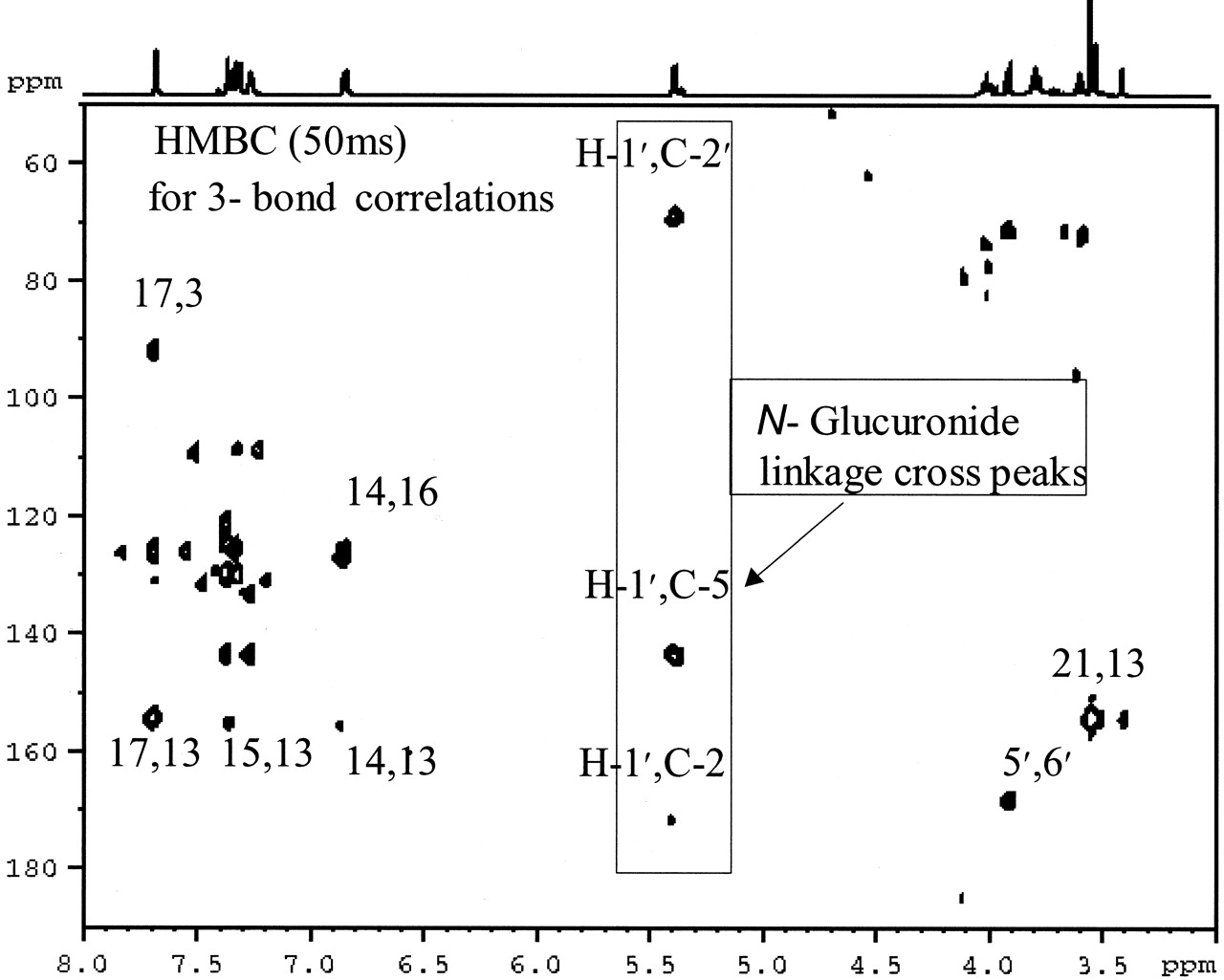
Long-range 1H-13C NMR correlation analysis of N-glucuronide of BMS-204352.
Stability of N-Glucuronide of BMS-204352. In the incubations of [14C]N-glucuronide of BMS-204352 in buffers at different pH levels, one degradation product accounting for <5% of the radioactivity used was eluted before the N-glucuronide of BMS-204352, which was observed in the buffers of all three buffers at pH levels 3.0, 7.4, and 9.0. Only the incubation at pH 3.0 generated a detectable level of BMS-204352. In the incubations of [14C]N-glucuronide of BMS-204352 with β-glucuronidases from both H. pomatia and E. coli, less than 10% of the N-glucuronide of BMS-204352 was hydrolyzed over 24 h. These results indicated that the N-glucuronide of BMS-204352 was resistant to hydrolysis at three different pH levels and by two β-glucuronidases tested.
Formation of N-Glucuronide of BMS-204352 by UGT2B7. Among the seven human cDNA-expressed UDP-glucuronosyltransferases (1A1, 1A3, 1A4, 1A6, 1A7, 1A10, and 2B7) tested, only the incubation of [14C]BMS-204352 with UGT2B7 in the presence of UDPGA produced a metabolite that had the same retention time (20.5 min) and LC/MS properties as the N-glucuronide of BMS-204352. The incubation with the pore-forming molecule alamethicin added (50 μg/ml) produced a larger quantity of the N-glucuronide of BMS-204352 (3 times) than the incubations without alamethicin added. The result is consistent with the known properties of other glucuronidation reactions (Fisher et al., 2000). These results suggest that formation of the N-glucuronide of BMS-204352 was catalyzed by UGT2B7 in vitro. Figure 6 shows that formation of the N-glucuronide follows Michaelis-Menten kinetics by cDNA-expressed UGT2B7 (Km = 14.22 ± 1.67 μM, Vmax = 0.29 ± 0.01 nmol/min · mg protein) and in the pooled HLM (Km = 6.44 ± 1.60 μM, Vmax = 0.48 ± 0.03 nmol/min · mg). The kinetic measurement in the incubations with the expressed UGT2B7 was consistent with that from incubations with the pooled HLM. The minor difference in Km values observed between two microsomal preparations was probably due to experimental variations. However, it is not known whether there is an unknown minor UGT isozyme that might also contribute to formation of the N-glucuronide of BMS-204352 in HLM. The results also suggest that BMS-204352 might be a potentially selective and relatively high affinity substrate for human UGT2B7.

Concentration-dependent rate of the formation of N-glucuronide of BMS-204352 in cDNA-expressed UGT2B7 and HLM in the presence of UDPGA.
The samples were analyzed by HPLC under gradient II conditions as described under Materials and Methods.
Discussion
Glucuronidation represents one of the major elimination pathways of BMS-204352 in humans and dogs. About 17% of the dosed radioactivity of [14C]BMS-204352 was excreted into human urine, and 33% of the dose was excreted in dog bile as the N-glucuronide of BMS-204352. Due to a low concentration of the metabolite in human urine, the metabolite was isolated from dog bile for structural elucidation by NMR. There are two possible linkages for the glucuronic sugar moiety to the amide functional group of BMS-204352: N-glucuronidation or tautomeric O-glucuronidation. The two-dimensional NMR experiments clearly demonstrated that the glucuronic acid linkage to BMS-204352 is through the amide nitrogen. Recently, two tautomeric S-glucuronides formed from thio-amides were reported (Ethell et al., 2003; Martin et al., 2003). Smith and Williams (1949) identified the first N-glucuronide of aniline in urine of rabbit dosed with aniline. Subsequently, many primary, secondary, and tertiary amines have been shown to be excreted as N-glucuronide metabolites (Green and Tephly, 1998). N-Glucuronidation of many primary and secondary aromatic amines has been suggested to have implications in the potential carcinogenicity of these compounds (Babu et al., 1992; Kadlubar et al., 1997). N-Glucuronidation of tertiary amines to quaternary ammonium-linked glucuronide metabolites contributes to the elimination of many therapeutic agents like tricyclic antidepressants and antihistamines. Several reports suggested formation of amide N-glucuronides. N-Glucuronide of 3,4,4′-trichlorocarbanilide was identified based on mass spectrometric analyses of plasma and urine of humans dosed with the drug (Gruenke et al., 1987). N-Glucuronide of carbamazepine was excreted into urine and characterized by mass spectrometric analysis (Lynn et al., 1978). Sulfadimethoxine and sulfathiazole might produce sulfonamide N-glucuronide metabolites (Bridges et al., 1961; Dulik and Fenselau, 1987). The glucuronide of BMS-204352 was the first example of glucuronidation linkages at amide nitrogen that was unambiguously identified by NMR.
Although N-glucuronide metabolites of the primary/secondary amines and N-glucuronides of N-hydroxylated amines were hydrolyzed to the parent compounds and glucuronic acid under mild, acidic conditions (Babu et al., 1992; Kadlubar et al., 1997), the quaternary-ammonium glucuronides were hydrolyzed under basic conditions (Dulik and Fenselau, 1987; Kowalczyk et al., 2000). Among the several quaternary-ammonium glucuronides tested for stability in the presence of β-glucuronidase from bovine liver, mollusks (H. pomatia), and bacteria (E. coli), only E. coli enzyme hydrolyzed all N-glucuronides (Kowalczyk et al., 2000). In contrast, N-glucuronide of BMS-204352 was resistant to hydrolysis at pH levels of 3.0, 7.4, and 9.0 and by mollusks' and bacterial β-glucuronidases.
Although definite identification of UGT isozymes (other than UGT1A1) was difficult at the present time due to lack of specific substrates and inhibitors for correlation studies and selective inhibition studies, a characterization of a UGT isozyme responsible for a glucuronidation reaction has become possible because of the progress made in the cloning and expression of UGTs. There were studies on identification of isozymes for N-glucuronidation of primary and secondary amines (Green and Tephly, 1998). UGT1A3 and 1A4 were involved in N-glucuronidation of substrates at the tertiary amine (Green and Tephly, 1998). Ten UGT isozymes (including 1A3, 1A4, and 2B7) failed to catalyze the formation of the N-glucuronides of nicotine and continine (Ghosheh and Hawes, 2002). Glucuronidation of BMS-204352 at the amide nitrogen represents a newly identified glucuronidation reaction by UGT2B7. UGT2B7 has been demonstrated to conjugate many drugs including morphine (Coffman et al., 1997), zidovudine (Barbier et al., 2000), epirubicin (Innocenti et al., 2001), estrogens and retinoic acid (Czernik et al., 2000), diclofenac (King et al., 2001), and rofecoxib (Zhang et al., 2003b). These reactions represent glucuronidations by UGT2B7 at nucleophilic functional groups of oxygen (carboxylic acid, phenol, and primary and secondary alcohols). Glucuronidation of morphine and zidovudine was competitively inhibited by UGT2B7 substrate and inhibitors such as oxazepam, naltriben, and etonitazenyl (Rios and Tephly, 2002). UGT2B7 is an inducible enzyme (Munzel et al., 1999) and had more than 10 single nucleotide polymorphisms (Holthe et al., 2003). There is a significant interindividual variability in glucuronidation of zidovudine and morphine (Pacifici et al., 1996; Holthe et al., 2002). Identification of the specific enzyme UGT2B7 for N-glucuronidation of MaxiPost will allow future investigation of potential drug-drug interaction as well as possible polymorphism in N-glucuronidation of MaxiPost, its major clearance pathway. The role of UGT2B7 in the metabolism of BMS-204352 in the brain is yet to be discovered.
In summary, a major elimination pathway of MaxiPost (BMS-204352) is an amide N-glucuronidation in humans. The responsible enzyme for formation of the N-glucuronide of BMS-204352 is UGT2B7. Both the amide N-glucuronide structure and the N-glucuronidation reaction catalyzed by UGT2B7 represent new findings. Additionally, BMS-204352 may serve as a relatively high affinity probe to study UGT2B7.
Acknowledgments
We thank Marc Ogan and Kent Rinehart for synthesis of [14C]BMS-204352 and Gary Skiles for critical review of the manuscript.
Footnotes
-
↵1 Abbreviations used are: UGT, UDP-glucuronosyltransferase; BMS-204352, (3S)-(+)-(5-chloro-2-methoxyphenyl)-1,3-dihydro-3-fluoro-6-(trifluoromethyl)-2H-indole-2-one); LC/MS, liquid chromatography/mass spectrometry; HPLC, high-performance liquid chromatography; UDPGA, uridine 5′-diphospho-glucuronic acid; HLM, human liver microsomes.
- Received October 29, 2003.
- Accepted January 29, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}