


Abstract
Human carbonyl reductase (CBR) activity accounts for a significant fraction of the metabolism of endogenous and xenobiotic carbonyl compounds. It is possible that genetic polymorphisms in CBR1 and CBR3 are key for the wide interindividual variability in the disposition of CBR drug substrates. We pinpointed a single nucleotide polymorphism in CBR3 (CBR3 V244M) that encodes for a V244 to M244 change. Blacks showed a higher frequency of the M244 allele (q = 0.51, n = 49) than did whites (q = 0.31, n = 70; p = 0.003). In addition, DNA variation panels from 10 ethnic groups presented a wide range of CBR3 V244M genotype distributions. Kinetic experiments with the recombinant CBR3 protein variants and menadione revealed that CBR3 M244 has significantly higher Vmax than does CBR3 V244 (Vmax CBR3 M244 = 40.6 ± 1.3 μmol/min · mg versus Vmax CBR3 V244 = 19.6 ± 2.0 μmol/min · mg, p = 0.002). In contrast, both isoforms presented similar Km values (Km CBR3 M244 = 22.9 ± 2.9 μM versus Km CBR3 V244 = 24.6 ± 3.2 μM, p = 0.43). Assays with NADP(H) demonstrated a higher VmaxNADP(H) (1.6-fold) and increased catalytic efficiency (VmaxNADP(H)/KmNADP(H)) for CBR3 M244 compared with CBR3 V244 (p = 0.013). Comparative three-dimensional analyses based on the structure of the homologous porcine carbonyl reductase suggested that the V244M substitution is positioned in a region critical for interactions with the NADP(H) cofactor. These studies demonstrate that the common CBR3 V244M polymorphism encodes for CBR3 isoforms with distinctive enzymatic properties.
Carbonyl reductases (CBRs) are members of the family of shortchain dehydrogenases/reductases. In humans, there are two monomeric carbonyl reductases, carbonyl reductase 1 (CBR1) and carbonyl reductase 3 (CBR3), which are encoded by different genes located 62 kilobases apart on chromosome 21 (CBR1 and CBR3) (Avramopoulos et al., 1992; Watanabe et al., 1998; Forrest and Gonzalez, 2000). CBR1 and CBR3 have 72% identity and 79% similarity at the amino acid level as indicated by global alignment analysis. CBRs metabolize a broad spectrum of endogenous and xenobiotic carbonyl compounds such as prostaglandins, steroids, and various pharmacological agents (Forrest and Gonzalez, 2000). Despite the prominent role of CBR-mediated drug metabolism, there have been no reports on the catalytic properties of CBR3 since the identification of the gene in 1998 (Watanabe et al., 1998). Several studies have described a wide range of interindividual variability in the metabolism of drugs that are CBR substrates (Hermans and Thijssen, 1992; Wong et al., 1992, 1993; Iwata et al., 1993; Ohara et al., 1995; Rady-Pentek et al., 1997; Maser et al., 2000). For example, the anticancer anthracyclines doxorubicin and daunorubicin are extensively reduced by CBRs in normal tissues as well as in tumors into their respective alcohol metabolites doxorubicinol and daunorubicinol (Forrest et al., 1990). Variable CBR activity may contribute to the unpredictable pharmacokinetics and pharmacodynamics of anthracyclines in adult and pediatric cancer patients (Ratain et al., 1991; Krischer et al., 1997; Singal and Iliskovic, 1998; Frost et al., 2002). We hypothesize that genetic polymorphism may play a relevant, although thus far uncharacterized role in modulating CBR activity. Thus, we pinpointed a novel nonsynonymous single nucleotide polymorphism in CBR3 that encodes for a V244 to M244 change in the protein. We determined the frequencies of the variant CBR3 alleles in genomic DNA samples from white and black liver donors, and in DNA variation panels representing different ethnic groups. Furthermore, the recombinant CBR3 V244 and CBR3 M244 isoforms were kinetically characterized with menadione and NADP(H). The distinctive catalytic properties of the CBR3 V244 and CBR3 M244 isoforms support the notion that the common CBR3 V244M genetic polymorphism may impact CBR-mediated biotransformation.
Materials and Methods
Samples. DNA samples were procured from 117 liver donors (whites, n = 70, and blacks, n = 47) from the Liver Tissue Resource (Pharmacogenetics of Anticancer Research Group, http://www.pharmacogenetics.org/). Human DNA variation panels were purchased from the Coriell Institute for Medical Research (Camden, NJ). Each panel contained DNA samples from individuals representing the following ethnic groups: Chinese, Indo-Pakistani, Japanese, Mexican, Middle Eastern, South American (Andes region), South East Asian (excluding Japanese and Chinese), Pacific, African (North of the Sahara), and African (South of the Sahara).
Genotyping. We investigated the CBR3 V244M polymorphism with two genotyping techniques. The first approach involved PCR amplification of CBR3 exon 3 followed by SNP detection by direct sequencing analysis (Nickerson et al., 1997). PCR primers were as follows: forward, 5′-TGGAGGACACAAAAAATGAGG-3′; and reverse, 5′-GAGAGAGAGAGAGAGAAAAGAAAG-3′. Each 50-μl PCR contained 10 ng of genomic DNA, nuclease-free water, 50 pmol of each primer, 2.5 units of AmpliTaq Gold DNA polymerase (PerkinElmer Life and Analytical Sciences, Boston, MA), 250 μM each deoxynucleotide-5′-triphosphate (Invitrogen, Carlsbad, CA), and Gene Amp PCR buffer (PerkinElmer Life and Analytical Sciences). Amplification conditions were 92°C for 5 min, 40 cycles of 92°C for 30 s, 56°C for 40 s, and 70°C for 50 s, followed by a final extension at 70°C for 10 min. PCR products (761 base pairs) were incubated with ExoSAP-IT (USB, Cleveland, OH) to remove unincorporated nucleotides and primers. PCR products were sequenced on an ABI Prism 3700 sequencer (Applied Biosystems, Foster City, CA) with the forward PCR primer and with the internal sequencing primer 5′-AGCAAGCTCCGAAGCAGA-3′. The resultant trace files were assembled and analyzed using the public National Cancer Institute SNP server (http://lpgws.nci.nih.gov/perl/snp/snp_cgi.pl). The second CBR3 V244M genotyping approach utilized the TaqMan technology with specific fluorescent probes labeled with FAM and VIC for discrimination of the G and A alleles, respectively (Applied Biosystems, Assays-by-Design). Genotyping reactions were performed according to the manufacturer's specifications in an MX 4000 thermal cycler (Stratagene, La Jolla, CA). Genotyping results were analyzed with the MX 4000 proprietary software. We performed extensive testing to evaluate the performance of the CBR3 V244M TaqMan assays. There was 100% concordance in the CBR3 V244M genotype cells determined by both direct sequencing and TaqMan, respectively.
Cloning and Expression of Human CBR3 V244 and CBR3 M244.CBR3 V244 and CBR3 M244 full-length cDNAs (1003 base pairs) were obtained by reverse transcription-PCR from liver RNA samples of individuals with CBR3 V244M homozygous G/G (valine244), and homozygous A/A (methionine244) genotypes, respectively. Total RNA (1 μg) was reverse-transcribed with oligo(dTs) (Invitrogen), and the cDNA was amplified with primers located at the 5′- and 3′-untranslated regions using the Expand Long Template PCR System (Roche Diagnostics, Indianapolis, IN). Primers were as follows: forward, 5′-GCTCAGCCATGTCATCCT-3′; and reverse, 5′-CTCCGAAGCAGACGTTTACC-3′. Amplification conditions were 92°C for 2 min, 45 cycles of 92°C for 35 s, 54°C for 40 s, and 68°C for 1:10 min, followed by a final extension at 68°C for 10 min. Purified PCR products were cloned into pET28a expression vectors (Novagen, Madison, WI). The inserts were fully sequenced with vector- and insert-specific primers. The CBR3 V244 and CBR3 M244 constructs were transfected into Escherichia coli BL21 (DE3)-competent cells using the heat shock method. Cells were plated on LB agar supplemented with kanamycin (30 μg/ml) for selection. Five clones were randomly picked and grown overnight in 100 ml of LB broth supplemented with kanamycin (15 μg/ml) to optimize expression. Selected cultures were expanded up to 2 liters (A600 nm ≈ 0.2) and grown at 37°C (A600 nm ≈ 0.6–0.7). Isopropyl-β-d-thiogalactoside was added to a final concentration of 1 mM, and the cultures were grown for another 3 to 4 h. Cells were pelleted by centrifugation, washed, and resuspended in lysis buffer (50 mM Na2HPO4, 300 mM NaCl, and 10 mM imidazole) supplemented with lysozyme (1 mg/ml), DNase I (5 μg/ml), and ribonuclease (10 μg/ml). Cell lysates were obtained by sonication (three pulses of 30 s at 4°C) and centrifuged for 20 min (24,000g at 4°C). The resulting cell extracts (supernatants) were filtered (0.45-μm membrane) and transferred to a vial with equilibrated nickel-nitrilotriacetic acid resin. Binding occurred in a rotator at 4°C overnight. Proteins were eluted with buffer containing 250 mM imidazole. The proteins were concentrated using Centricon membranes (YM10; 10,000 molecular weight cut-off) and extensively dialyzed for 24 h in phosphate-buffered saline at 4°C with four to five buffer changes (× 2.5 liters). After dialysis, protein fractions were desalted by chromatography on a Sephadex G-25 fine column (Amersham Biosciences Inc., Piscataway, NJ). Different fractions (e.g., before and after sonication, wash, and elutions) were collected for analysis by SDS-polyacrylamide gel electrophoresis followed by Coomassie staining and Western blotting with anti-His-tag antibodies. CBR3 M244 and CBR3 V244 were expressed as discrete 39-kDa single bands.
Kinetic Analysis. CBR3 V244 and CBR3 M244 enzymatic activities were measured by recording the rate of oxidation of the NADP(H) cofactor at 340 nm (NADPH molar absorption coefficient, 6220 M-1 cm-1). CBR3 concentrations were determined from the absorbance at 280 nm (molar extinction coefficient of 15,220 M -1cm-1; http://us.expasy.org). Protein concentrations and incubation times were standardized to ensure strict obedience to conditions of initial velocity (Vo). Experiments were performed in a Cary-Varian Bio 300 UV-visible spectrophotometer equipped with thermal control and proprietary software for quality control and enzyme kinetics data analysis. Reactions were incubated at 37°C and monitored for 5 min at an acquisition rate of 10/s (3000 readings). Assay mixtures (1.0 ml) contained 0.1 mM NADP(H) (Sigma-Aldrich, St. Louis, MO), potassium phosphate buffer (pH 7.4, 0.1 M), and enzyme (CBR3 V244 or CBR3 M244). Menadione (Sigma-Aldrich) concentrations ranged from 3 to 200 μM. For the determination of the kinetic constants of the cofactor, NADP(H) concentrations ranged from 10 to 200 μM, with menadione at a fixed concentration of 150 μM. Enzymatic velocities were automatically calculated by linear regression of the ΔAbs/Δ time points. Regression coefficients r ≥ 0.98 were obtained for all substrate concentrations. Km and Vmax values were obtained by nonlinear regression analysis with Sigma Plot version 8.0 (SPSS Inc., Chicago, IL). Excellent agreement between the experimental and predicted Km and Vmax values was obtained in all cases with regression coefficients r ≥ 0.9 and CV ≤ 10%, respectively.
Results and Discussion
The CBR3 V244M polymorphism was pinpointed from the dbSNP database (refSNP ID: rs1056892). First, we sought to confirm the polymorphism in DNA samples from liver donors. Therefore, we analyzed the CBR3 V244M genotype distributions in 70 DNA samples from white donors, and in 47 DNA samples from black donors. In whites, the genotype distribution was as follows: 47.1% were homozygous for the G allele (p = 0.69), 44.3% were heterozygous G/A, and 8.6% were homozygous for the A allele (q = 0.31). In contrast, 17.0% of blacks presented the homozygous G/G genotype (p = 0.49), 69.8% were heterozygous G/A, and 19.1% were homozygous A/A (q = 0.51). Allele distributions were in Hardy-Weinberg equilibrium in both groups (chi square test, p = 0.98 for whites and p = 0.14 for blacks). Comparisons of the allelic frequencies indicated that the CBR3 V244M genotype distributions differed significantly between blacks and whites (chi square = 11. 7, p = 0.003) (Fig. 1). We extended our observations by analyzing the CBR3 V244M genotype distributions in 10 human variation panels from the Coriell Institute. CBR3 V244M allele frequencies varied widely among the groups (Table 1). For example, 85.7% of individuals from the Pacific group were homozygous for the G allele (V244), whereas 55.6% of the samples from the Indo-Pakistani panel presented the homozygous A/A genotype (M244).

CBR3 V244M genotype distributions in whites (n = 70) and blacks (n = 49). Allelic frequencies for whites were p = 0.69 (G), q = 0.31 (A), and those for blacks were p = 0.49 (G), q = 0.51 (A).
Distribution of CBR3 V244M genotypes in DNA human diversity panels
The percentages corresponding to each genotype category are indicated in parentheses.
To evaluate the functional impact of the CBR3 V244M polymorphism, we first characterized the catalytic properties of the recombinant CBR3 V244 and CBR3 M244 isoforms with the prototypical quinone substrate menadione. Menadione or vitamin K3 is considered a good CBR substrate in humans and other species with Km values in the 5 to 50 μM range (Wermuth et al., 1988; Park et al., 1991; Inazu et al., 1992; Bohren et al., 1994; Schaller and Wermuth, 1999; Sugiyama et al., 2004). The Km values for CBR3 V244 and CBR3 M244 did not differ (Student's t test, p = 0.43). However, both isoforms showed significant differences in Vmax (Table 2). CBR3 M244 presented a 2-fold higher Vmax than did CBR3 V244 (Student's t test, p = 0.002). The higher catalytic efficiency (Vmax/Km) of the CBR3 M244 isoform results in an enzyme that is 100% more efficient in catalyzing the reduction of substrate per unit of time.
Kinetic constants of CBR3 V244 and CBR3 M244
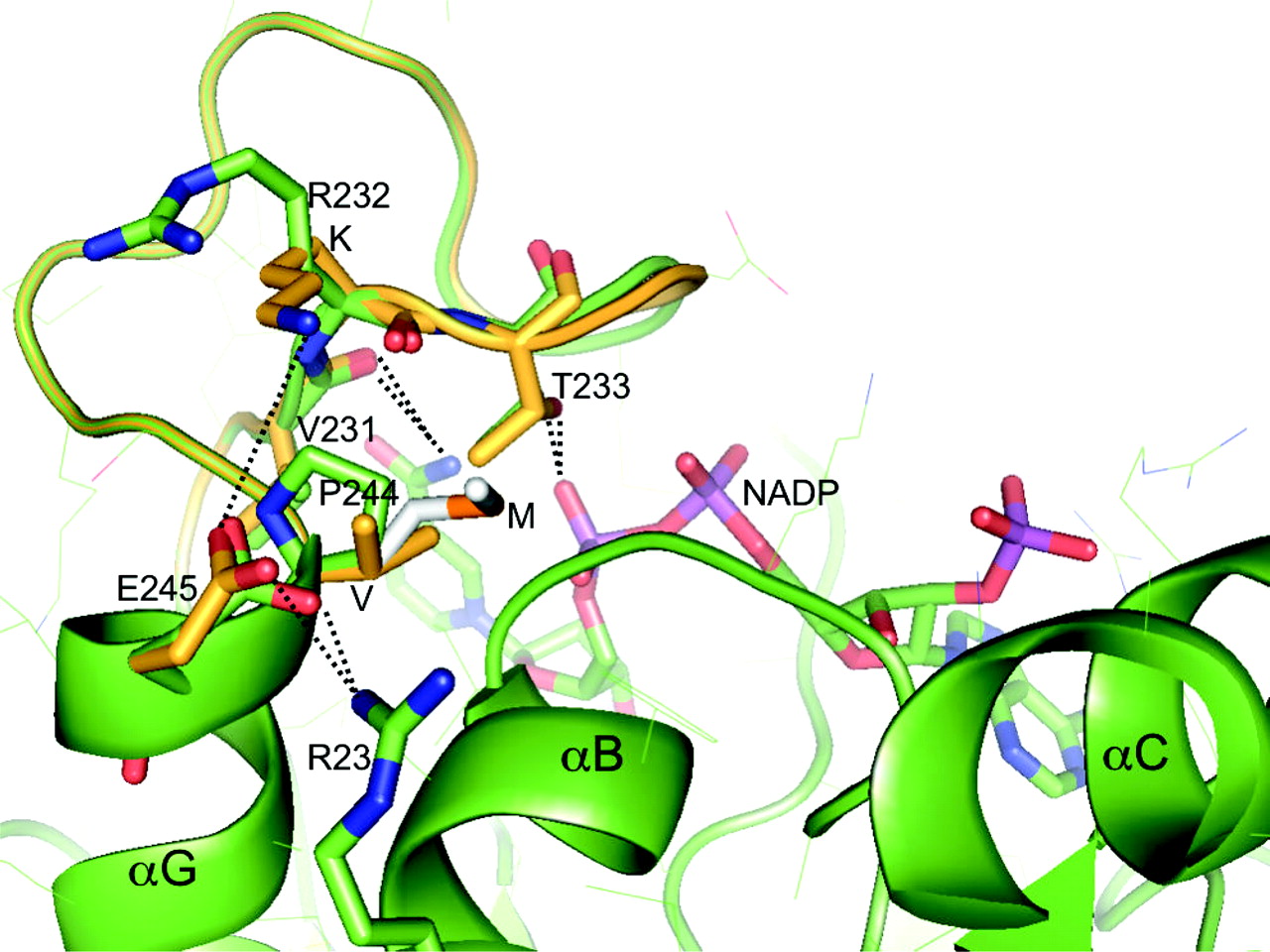
To further characterize the impact of the V244M substitution, we performed kinetic studies with varying concentrations of NADP(H) (Sciotti and Wermuth, 2001; Sugiyama et al., 2004). Interestingly, KmNADP(H) values were similar for both CBR3 isoforms, but the VmaxNADP(H) of CBR3 M244 was 1.6-fold higher than the Vmax-NADP(H) of CBR3 V244 (Student's t test, p = 0.013; Table 2). Thus, the kinetic data suggest that the V244M substitution may not affect direct interactions between the NADP(H) cofactor and the neighboring amino acids on the CBR3 peptide backbone. Instead, the higher apparent VmaxNADP(H) of the CBR3 M244 isoform may be reflecting a faster association-dissociation rate for NADP(H)/NADP per catalytic cycle. To explore whether the three-dimensional structure of the enzyme can provide any clues to this possibility, we constructed a model of human CBR3 and examined the effect of the V244M polymorphism. The human CBR3 structure was derived from the crystal structure of porcine testicular carbonyl reductase (PTCR) (Ghosh et al., 2001), with which it shares about 70% sequence identity (global alignment analysis; GCG-SeqWeb version 2.0, Accelrys, San Diego, CA), by substituting amino acids in the regions surrounding the coenzyme and the substrate binding sites, and then optimizing conformations of the backbone and side chains in those regions. The molecular modeling software CHAIN (Sack, 1988) was used for this purpose. Figure 2 shows a modeling of V244 and M244 side chains on the three-dimensional structure of the NADP(H) binding region of human CBR3. The residue at position 244 is a proline in PTCR (P243 according to the PTCR numbering), which is the amino-terminal residue of the helix αG, adjacent to the NADP(H) binding site (Fig. 2). The model was obtained first by mutating the proline side chain to a valine and then optimizing the conformation of the loop between residues 230 and 235 such that the backbone carbonyl group of K232 can now approach the freed backbone amide group of V244 for a possible hydrogen bond. Small differences between human CBR3 (Fig. 2, shown in yellow) and PTCR (Fig. 2, shown in green) in the conformation of this loop are likely, which could also result from several substitutions in the region 236–241 between PTCR and human CBR3. Interestingly, this segment of the polypeptide chain has two polar contacts to the NADP(H) molecule (T233 side chain to the pyrophosphate moiety and V231 carbonyl to the amide group of the nicotinamide ring; Fig. 2) that could be important for catalysis. Furthermore, a small movement of the loop could be accompanied by shifting of the K232 (R231 in PTCR) side chain to a closer proximity to E245, such that a new salt bridge is formed between K232 and E245, which already has a salt bridge with R23. Thus, more tightening of this loop is anticipated in human CBR3, in comparison with PTCR. A valine to methionine substitution at this position (the M244 side chain is shown with gray carbon atoms; Fig. 2) due to the CBR3 SNP is nicely accommodated in a hydrophobic pocket between α-helices G and B. Thus, a further compacting of the tertiary structure of this region due to increased hydrophobic interaction could result in CBR3 M244. This structural variation may influence the docking and/or release of the cofactor during enzymatic catalysis. Although a detailed analysis of structural differences due to the SNP is impossible in the absence of experimental CBR3 V244 and CBR3 M244 structures, the modeling exercise suggests alterations in the dinucleotide-binding interactions, which may explain the observed higher catalytic efficiency of CBR3 M244.

The three-dimensional structure of porcine testicular carbonyl reductase (PTCR) in green with modeled human CBR3 in yellow, shown at the site of V244M substitution. V244 is a proline (P) in PTCR. The M244 side chain is shown with gray carbon and orange sulfur atoms. Other side chain atoms are also color-coded: carbon, green or yellow; oxygen, red; nitrogen, blue; and phosphorus, purple. A NADP molecule as observed in the crystal structure of PTCR is also shown. Side chains and ligands of interest are drawn as thick bonds. The rest of the PTCR side chains are drawn in thin lines. Hydrogen bond- and salt bridge-forming interactions relevant to the discussion are shown by dotted lines in both PTCR and CBR3 structures. See text for detailed explanation.
In conclusion, our results indicate that the CBR3 V244M polymorphism is common among different ethnic groups and encodes for CBR3 protein isoforms with distinctive catalytic properties. Our observations warrant further research to elucidate the role of polymorphic CBR3 in the variable metabolism of CBR drug substrates.
Acknowledgments
We thank Dr. Mary V. Relling and the Pharmacogenetics of Anticancer Agents Liver Tissue Resource (http://www.pharmacogenetics.org/) for valuable liver tissue samples. The excellent assistance of Jessica Haas, Vanessa Gonzalez, and Erick Vasquez is gratefully acknowledged.
Footnotes
-
This work was supported by grants from The Kapoor Foundation and from the Lance Armstrong Foundation (principal investigator, Smita Bhatia), and by National Institutes of Health Grant CA16056.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.104.002006.
-
ABBREVIATIONS: CBR, carbonyl reductase; PCR, polymerase chain reaction; SNP, single nucleotide polymorphism; PTCR, porcine testicular carbonyl reductase.
- Received September 16, 2004.
- Accepted November 9, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}