


Abstract
The purpose of the present study was to explore the utility of sandwich-cultured rat hepatocytes as an in vitro tool to examine drug interactions at the hepatic transport level. Rhodamine 123 was used as a model substrate for P-glycoprotein-mediated biliary excretion. Effects of various types of P-glycoprotein modulation on the biliary excretion index (BEI; a relative measure of the extent of biliary excretion) and the in vitro biliary clearance (CLbile) were determined. Significant reductions in rhodamine 123 BEI and CLbile were noted in the presence of the P-glycoprotein inhibitors verapamil (30–100 μM) and progesterone (100 μM). The P-glycoprotein activator quercetin (10–100 μM) enhanced rhodamine 123 CLbile by approximately 4-fold, with only a minor effect on BEI, suggesting that quercetin had a more pronounced effect on uptake at the basolateral membrane rather than excretion across the canalicular membrane. Treatment of hepatocytes for 48 h with dexamethasone (10 μM) resulted in significant enhancement of CLbile, whereas rifampin (5–50 μM) increased both BEI and CLbile, indicating that the inducing effects of dexamethasone and rifampin were occurring at the basolateral and canalicular membranes, respectively. Total rhodamine 123 uptake in sandwich-cultured rat hepatocytes was partly saturable and was affected by the presence of typical Oatp1a4 substrates (digoxin, quinine, d-verapamil, 17β-estradiol-d-17β-glucuronide). In summary, sandwich-cultured rat hepatocytes are a useful tool to study mechanisms of hepatobiliary drug disposition and to predict the potential for drug interactions in hepatic transport.
The utility of sandwich-cultured rat hepatocytes as an in vitro model to predict the extent of biliary excretion of compounds was first established by Liu et al. (1999). Clearly, the important benefit of culturing hepatocytes in a collagen-sandwich configuration is that normal cell polarity is maintained, resulting in the presence of a basolateral (sinusoidal) and an apical (canalicular) membrane domain, each expressing different proteins involved in hepatic drug transport.
In a previous study, we optimized and validated the sandwich-cultured rat hepatocyte model for the in vitro study of biliary drug excretion mediated by P-glycoprotein. Inhibition of P-glycoprotein by the potent inhibitor GF120918 [N-(4-[2-(1,2,3,4-tetrahydro-6,7-dimethoxy-2-isoquinolinyl)ethyl]-phenyl)-9,10-dihydro-5-methoxy-9-oxo-4-acridine carboxamide] resulted in impaired in vitro biliary excretion of known P-glycoprotein substrates such as rhodamine 123 and digoxin, whereas substrate accumulation in the hepatocytes was unaffected. These results were consistent with P-glycoprotein expression on the canalicular membrane in this in vitro model, which is in agreement with the canalicular expression of P-glycoprotein in the normal hepatocyte in vivo (Thiebaut et al., 1987; Silverman and Schrenk, 1997).
One promising application of the sandwich-cultured rat hepatocyte model is the prediction of potential drug-drug interactions that occur at the level of hepatic transporters. Much less is known about transporter-mediated drug interactions compared with metabolism-based drug interactions. However, many studies have reported increased or reduced intestinal absorption of P-glycoprotein substrates following P-glycoprotein inhibition (Sababi et al., 2001) or induction (Greiner et al., 1999; Hamman et al., 2001), respectively. Inhibition of P-glycoprotein-mediated transport of digoxin by quinine or quinidine also has been shown to alter the renal and biliary elimination of digoxin (Hedman et al., 1990).
In the present study, the applicability of sandwich-cultured rat hepatocytes to predict P-glycoprotein-mediated hepatic drug interactions was demonstrated by studying the effects of various types of in vitro P-glycoprotein modulation (inhibition, activation, induction) on the in vitro hepatobiliary disposition of rhodamine 123, which is an established high-affinity P-glycoprotein substrate (Km = 13.5 μM; Shapiro and Ling, 1997). In addition, the unique ability of the sandwich-cultured rat hepatocyte model, due to maintenance of cell polarity in vitro, to simultaneously provide an assessment of hepatic uptake processes at the basolateral membrane, and biliary excretion processes at the canalicular membrane, was demonstrated in this study.
Materials and Methods
Animals. Male Wistar rats (Charles River Laboratories, Inc., Wilmington, MA) weighing 220 to 270 g were used for hepatocyte isolation. Animals had free access to water and food prior to surgery. All animal procedures were compliant with the guidelines of the Institutional Animal Care and Use Committee (University of North Carolina at Chapel Hill).
Materials. Collagenase (type I, class 1) was obtained from Worthington Biochemicals (Freehold, NJ). Dulbecco's modified Eagle's medium (DMEM) and insulin were purchased from Invitrogen (Carlsbad, CA). Rhodamine 123, dexamethasone, digoxin, verapamil, quinidine, quinine, progesterone, quercetin, rifampicin, 17β-estradiol-d-17β-glucuronide (E217G), bovine serum albumin (BSA), Triton X-100, 10× DMEM, soybean trypsin inhibitor, and fetal bovine serum (FBS) were purchased from Sigma-Aldrich (St. Louis, MO). ITS culture supplement (6.25 mg/ml insulin, 6.25 mg/ml transferrin, 6.25 μg/ml selenous acid, 5.35 mg/ml linoleic acid, and 1.25 g/ml BSA) and rat-tail collagen (type I) were obtained from BD Biosciences Discovery Labware (Bedford, MA). The reagents for total protein measurement with the bicinchoninic acid method were obtained from Pierce Chemical (Rockford, IL). All other chemicals and reagents were of analytical grade and were readily available from commercial sources.
Isolation and in Vitro Culture of Primary Rat Hepatocytes. Hepatocytes were isolated from male Wistar rats (220–270 g) using a collagenase perfusion as described previously (Annaert et al., 2001) with one modification: 4 mg of soybean trypsin inhibitor was added to the perfusion buffer at the same time that 50 to 75 mg of collagenase was added. The use of this trypsin inhibitor appeared to enhance cell yield and viability following hepatocyte preparations.
Preparation of Cell Culture Dishes and Sandwich-Cultured Hepatocytes. Culture dishes were coated with a collagen solution (1.5 mg/ml final concentration) prepared by neutralizing a mixture of 4 ml of rat-tail type I collagen, 4 ml of deionized water, and 1 ml of 10× DMEM with 1 ml of 0.2 N NaOH solution (final pH ∼7.4). Before plating the hepatocytes, 60-mm Permanox culture dishes (Nalge Nunc International, Naperville, IL) were coated with 0.2 ml of ice-cold neutralized type I collagen solution and placed overnight in a 37°C humidified incubator (Stericult 200; Thermo Forma, Marietta, OH). Each dish was rinsed with 3.0 ml of serum-free DMEM to hydrate the collagen prior to the addition of 3.0 × 106 cells. At 1 to 2 h after plating the cells, unattached cells were removed by replacing the cell plating medium with DMEM containing 5% FBS, 50 U/ml penicillin, 50 μg/ml streptomycin, 4 mg/l insulin, and 0.1 μM dexamethasone. At 24 h after plating (day 1), the medium was aspirated and the cells were overlaid with 200 μl of rat-tail collagen type I solution (1.5 mg/ml, pH 7.4) to obtain a “sandwich” configuration. One hour later, FBS-free DMEM (3.0 ml/dish, containing 1% ITS, 50 U/ml penicillin, 50 μg/ml streptomycin, 4 mg/l insulin, 0.1 μM dexamethasone) was added onto the cultures. The medium was changed every day with fresh FBS-free DMEM until day 4, when accumulation experiments were performed. For induction experiments, hepatocytes were treated with potential inducers (1 or 10 μM verapamil, 10 or 50 μM dexamethasone, 5 or 50 μM rifampicin) starting at 24 h after plating the hepatocytes by adding the appropriate concentrations to FBS-free DMEM. Treatment was continued for either 72 h or 48 h, by replacing the medium with freshly prepared medium every 24 h. Hepatocytes that were treated for 48 h received blank FBS-free DMEM (“washout”) during the last 24 h before the start of accumulation experiments.
Accumulation Experiments (Day 4). For rhodamine 123 accumulation studies, cells were rinsed twice with 2.0 ml of standard Hanks' balanced salt solution (HBSS at pH 7.4; 0.185 g/l CaCl2·2H2O, 0.09767 g/l MgSO4, 0.4 g/l KCl, 0.06 g/l KH2PO4, 0.35 g/l NaHCO3, 8.0 g/l NaCl, 0.04788 g/l Na2HPO4, 1.0 g/l glucose) and incubated in 3.0 ml of the same buffer for 10 min at 37°C. Subsequently, cells were incubated in 3.0 ml of 1 to 100 μM rhodamine 123 dissolved in either standard or Ca2+/Mg2+-free HBSS (pH 7.4; 0.4 g/l KCl, 0.06 g/l KH2PO4, 0.35 g/l NaHCO3, 8.0 g/l NaCl, 0.04788 g/l Na2HPO4, 1.0 g/l glucose, 0.38 g/l EGTA) for 30 min and rinsed four times with 3.0 ml of ice-cold standard HBSS. For experiments in which the effect of P-glycoprotein inhibition (verapamil, progesterone, quinidine) or activation (quercetin) was investigated, equal concentrations of the modulator were present in standard and Ca2+/Mg2+-free HBSS during the 30-min incubation periods. For experiments in which the effect of Oatp1a4 substrates (digoxin, d-verapamil, quinine, E217G) on basolateral uptake of rhodamine 123 was investigated (incubation with standard buffer only), these compounds were present in the standard HBSS during the 30-min incubation period. After rinsing, hepatocytes were lysed with 3.0 ml of 0.5% Triton X-100 solution by placing plates on a rotator for 20 min at room temperature. Cell lysates were analyzed by fluorescence spectroscopy (PerkinElmer LS50B; PerkinElmer Life and Analytical Sciences, Boston, MA) for determination of rhodamine 123 concentrations. Accumulation was normalized to the protein content of the hepatocytes in each dish as measured in duplicate aliquots with the bicinchoninic acid method using BSA as standard (0.2–1 mg/ml). All accumulation data were corrected for nonspecific binding (<0.2%) to collagen-coated, hepatocyte-free culture dishes.
Data Analysis. The biliary excretion index (BEI) was calculated using B-CLEAR technology (Qualyst, Inc., Research Triangle Park, NC) by dividing the difference in substrate accumulation between standard HBSS (cellular plus canalicular accumulation) and Ca2+-free HBSS (cellular accumulation) by the accumulation in standard HBSS (Liu et al., 1999). Alternatively, in vitro biliary clearance (CLbile; ml/min/mg protein) was calculated according to:  where C represents the initial concentration in the incubation medium. In vitro biliary clearance values were determined based on 200 mg of protein/g of liver and 40 g of liver/kg of rat body weight to obtain clearance values in ml/min/kg (Seglen, 1976). For the kinetic characterization of rhodamine 123 accumulation into sandwich-cultured rat hepatocytes, the following equation was used:
where C represents the initial concentration in the incubation medium. In vitro biliary clearance values were determined based on 200 mg of protein/g of liver and 40 g of liver/kg of rat body weight to obtain clearance values in ml/min/kg (Seglen, 1976). For the kinetic characterization of rhodamine 123 accumulation into sandwich-cultured rat hepatocytes, the following equation was used:  where Kd represents the rate constant for the nonsaturable accumulation component, and Km and Vmax represent the kinetic parameters for the saturable (Michaelis-Menten) component. The best fit of the equation to the rhodamine 123 accumulation data was obtained by nonlinear least-squares regression analysis using the Solver function in Microsoft Excel 2000 v.9.0 (Microsoft, Redmond, WA).
where Kd represents the rate constant for the nonsaturable accumulation component, and Km and Vmax represent the kinetic parameters for the saturable (Michaelis-Menten) component. The best fit of the equation to the rhodamine 123 accumulation data was obtained by nonlinear least-squares regression analysis using the Solver function in Microsoft Excel 2000 v.9.0 (Microsoft, Redmond, WA).
Statistics. Statistical data analysis was performed with the software program JMP v.3.2.6 (SAS Institute, Cary, NC). One-way ANOVA was used to compare BEI or biliary clearance values for multiple groups of data, followed by a Dunnett's post hoc test to allow pairwise comparisons of each treatment group with the control group. The criterion for statistical significance was p < 0.05. In the case of induction experiments (see Figs. 3 and 4), the method described by Larson (1992) was applied to allow statistical analysis (ANOVA, Dunnett's test) of summary data (averages, S.D., n) obtained for individual hepatocyte culture preparations.

Effect of typical inducers on BEI and biliary clearance of rhodamine 123 in sandwich-cultured rat hepatocytes. Hepatocytes were treated with inducers at concentrations as indicated for 72 h, before measuring rhodamine 123 accumulation. Hepatocytes were incubated in quadruplicate with rhodamine 123 (1 μM) for 30 min either in standard HBSS or in Ca2+/Mg2+-free HBSS. Black bars and striped bars represent average values (±S.E.M.) for relative BEI (top panel) or biliary clearance (bottom panel) as percentage of control values for hepatocytes from two different liver preparations. None of the BEI or biliary clearance values obtained for treatment groups were statistically different from control.
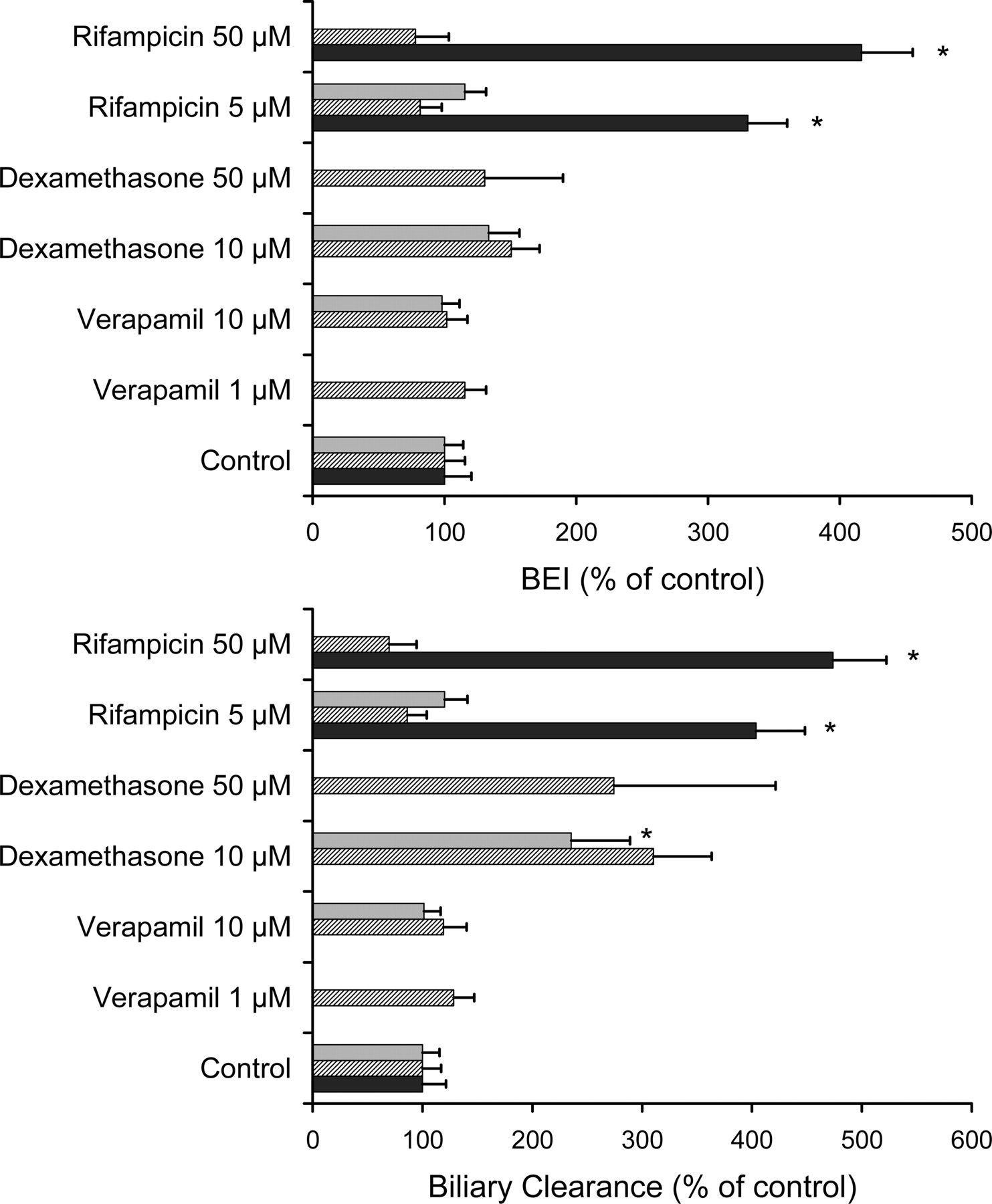
Effect of typical inducers on BEI and biliary clearance of rhodamine 123 in sandwich-cultured rat hepatocytes. Hepatocytes were treated with inducers at concentrations as indicated for 48 h, followed by a 24-h washout period, before measuring rhodamine 123 accumulation. Hepatocytes were incubated in quadruplicate with rhodamine 123 (1 μM) for 30 min either in standard buffer or in Ca2+/Mg2+-free buffer. Black bars, striped bars, and gray bars represent average values (±S.E.M.) for relative BEI (top panel) or biliary clearance (bottom panel) as a percentage of control values for hepatocytes from three different liver preparations. * denotes statistically significantly different from control value based on ANOVA/Dunnett's test (p < 0.05).
Results
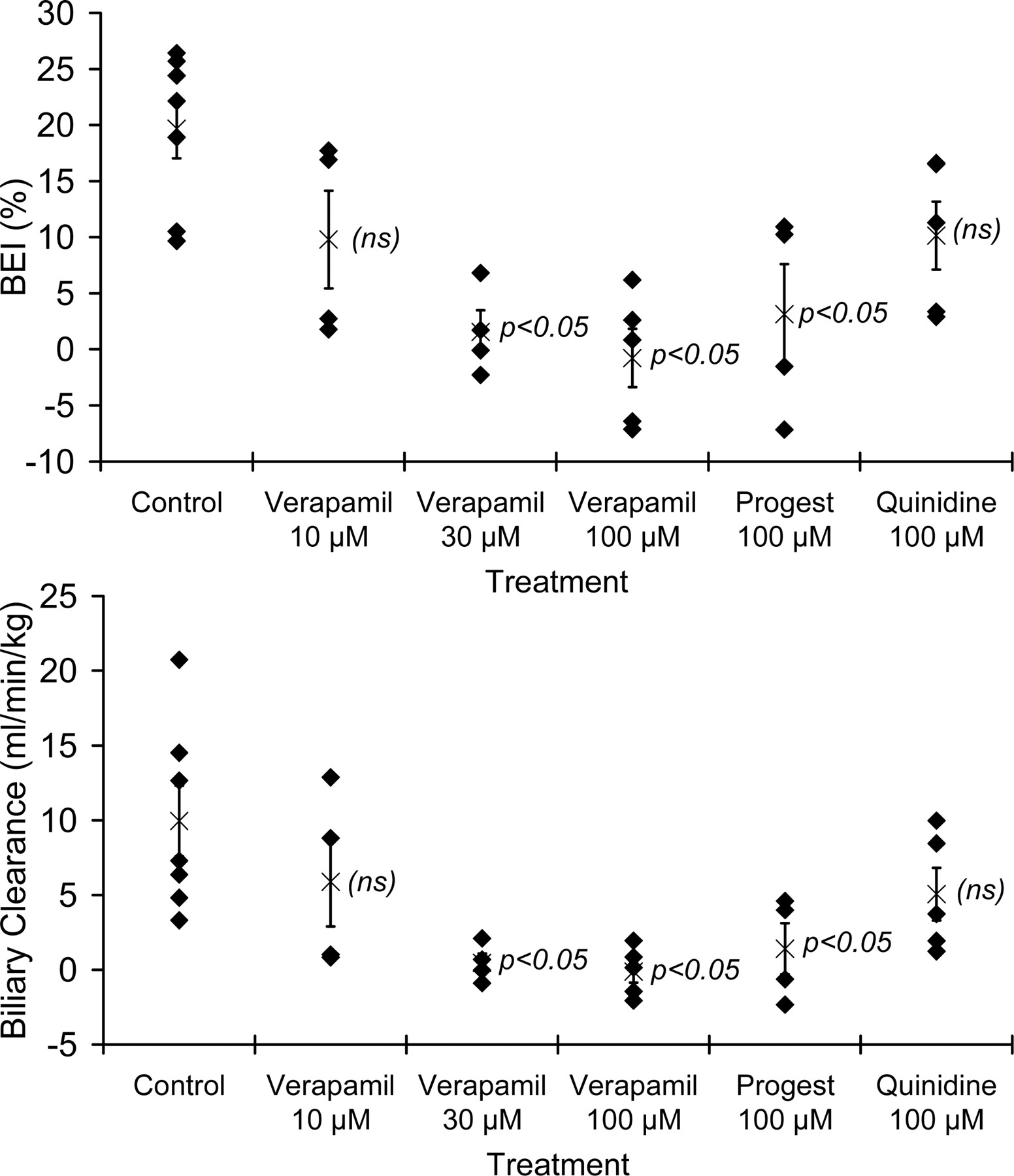
Inhibition of Rhodamine 123 Biliary Excretion. The effects of P-glycoprotein inhibition on the in vitro biliary excretion of rhodamine 123 were determined by measuring accumulation in day 4 sandwich-cultured rat hepatocytes in standard HBSS (representing intracellular + canalicular amounts of rhodamine 123) as compared with Ca2+/Mg2+-free HBSS (representing intracellular amounts of rhodamine 123), in the absence and in the presence of various “classical” P-glycoprotein inhibitors (verapamil, progesterone, quinidine). Differences in cellular + canalicular accumulation versus cellular accumulation represent biliary excretion as established previously (Annaert et al., 2001). The BEI values and CLbile values were calculated to express the relative and absolute extent of biliary excretion under various incubation conditions. As shown in Fig. 1, addition of 30 to 100 μM verapamil to the incubation buffer resulted in a concentration-dependent inhibition of in vitro rhodamine 123 biliary excretion; 100 μM verapamil completely inhibited rhodamine 123 biliary excretion. Progesterone (100 μM) also significantly reduced rhodamine 123 BEI and CLbile, when added to the culture medium. Quinidine (100 μM) also tended to decrease rhodamine 123 biliary excretion, although the differences failed to reach statistical significance. Rhodamine 123 BEI and CLbile were affected to similar extents compared with control for each inhibitor tested.

Effect of coincubation of classical P-glycoprotein inhibitors on BEI (percentage; top panel) and biliary clearance (ml/min/kg; bottom panel) of rhodamine 123. Sandwich-cultured rat hepatocytes were incubated in quadruplicate with rhodamine 123 (1 μM) for 30 min either in standard HBSS or in Ca2+/Mg2+-free HBSS, and in the absence (control) or presence of various inhibitors [verapamil, progesterone (Progest), quinidine; concentrations as indicated]. Diamonds represent average BEI or biliary clearance values obtained with hepatocyte cultures prepared from individual livers. Average values (±S.E.M.) across all hepatocyte preparations (n = 7) are also shown (×). p < 0.05 denotes statistically significant difference compared with control based on ANOVA/Dunnett's test. (ns) denotes no statistically significant difference from control.
Increased Rhodamine 123 Accumulation and Biliary Excretion in the Presence of the Flavonoid Quercetin. To evaluate the effects of P-glycoprotein activation on the hepatobiliary disposition of rhodamine 123 in sandwich-cultured rat hepatocytes, cells were coincubated with increasing concentrations of the flavonoid quercetin. The relative effects of quercetin on rhodamine 123 BEI and CLbile are shown in Fig. 2. Both BEI and CLbile tended to increase in the presence of quercetin. However, the effect only reached statistical significance for the CLbile value at 25 μM quercetin, indicating that quercetin primarily was affecting intracellular accumulation of rhodamine 123 (∼4-fold) with a much less pronounced effect on (P-glycoprotein-mediated) biliary excretion (∼1.5-fold). Figure 2 also illustrates that the effects of quercetin appear to be maximal at a concentration of approximately 25 μM.

Effect of increasing concentrations of quercetin (0–100 μM) on BEI (top panel) and biliary clearance (bottom panel) of rhodamine 123 in sandwich-cultured rat hepatocytes. Hepatocytes were incubated in quadruplicate with rhodamine 123 (1 μM) for 30 min either in standard HBSS or in Ca2+/Mg2+-free HBSS, and in the absence (control) or presence of increasing concentrations of quercetin. Triangles represent average BEI or biliary clearance values obtained with hepatocyte cultures prepared from individual livers. Average values (±S.E.M.) across all hepatocyte preparations (n = 6) are also shown (circles). p < 0.05 denotes statistically significant difference compared with control based on ANOVA/Dunnett's test.
Altered Biliary Excretion of Rhodamine 123 after Treatment with Inducing Agents. To evaluate the effects of increased transport protein expression, sandwich-cultured rat hepatocytes were cultured in the presence of established inducing agents, including verapamil (1–10 μM), rifampicin (5–50 μM), and dexamethasone (10–50 μM). Hepatocytes were cultured with inducers, either for 72 h (Fig. 3) or for 48 h, followed by a 24-h washout (Fig. 4), prior to determination of rhodamine 123 BEI and CLbile. When hepatocytes were treated for 72 h, none of the treatments significantly affected rhodamine 123 BEI and CLbile, although there appeared to be a clear trend toward a ∼2- to 3-fold increase in rhodamine 123 BEI and CLbile following a 10 μM verapamil treatment in one of the hepatocyte preparations. Interestingly, treatment of the cells for 72 h with 50 μM rifampicin appeared to reduce rhodamine 123 BEI and CLbile to negligible levels. However, when hepatocytes were cultured with 5 to 50 μM rifampicin for 48 h followed by a 24-h washout, rhodamine 123 BEI and CLbile increased up to 4-fold for one hepatocyte culture preparation (Fig. 4), whereas verapamil did not exert any effect. Culturing the cells for 48 h in the presence of 10 to 50 μM dexamethasone resulted in a 2- to 3-fold enhancement of rhodamine 123 CLbile, whereas the effects on rhodamine 123 BEI were negligible, consistent with increased intracellular accumulation of rhodamine 123. It is noteworthy that the 72-h treatment with 50 μM rifampicin, in addition to abolishing both BEI and CLbile, also resulted in a significant (by about 35%) decrease in rhodamine 123 accumulation (data not shown).
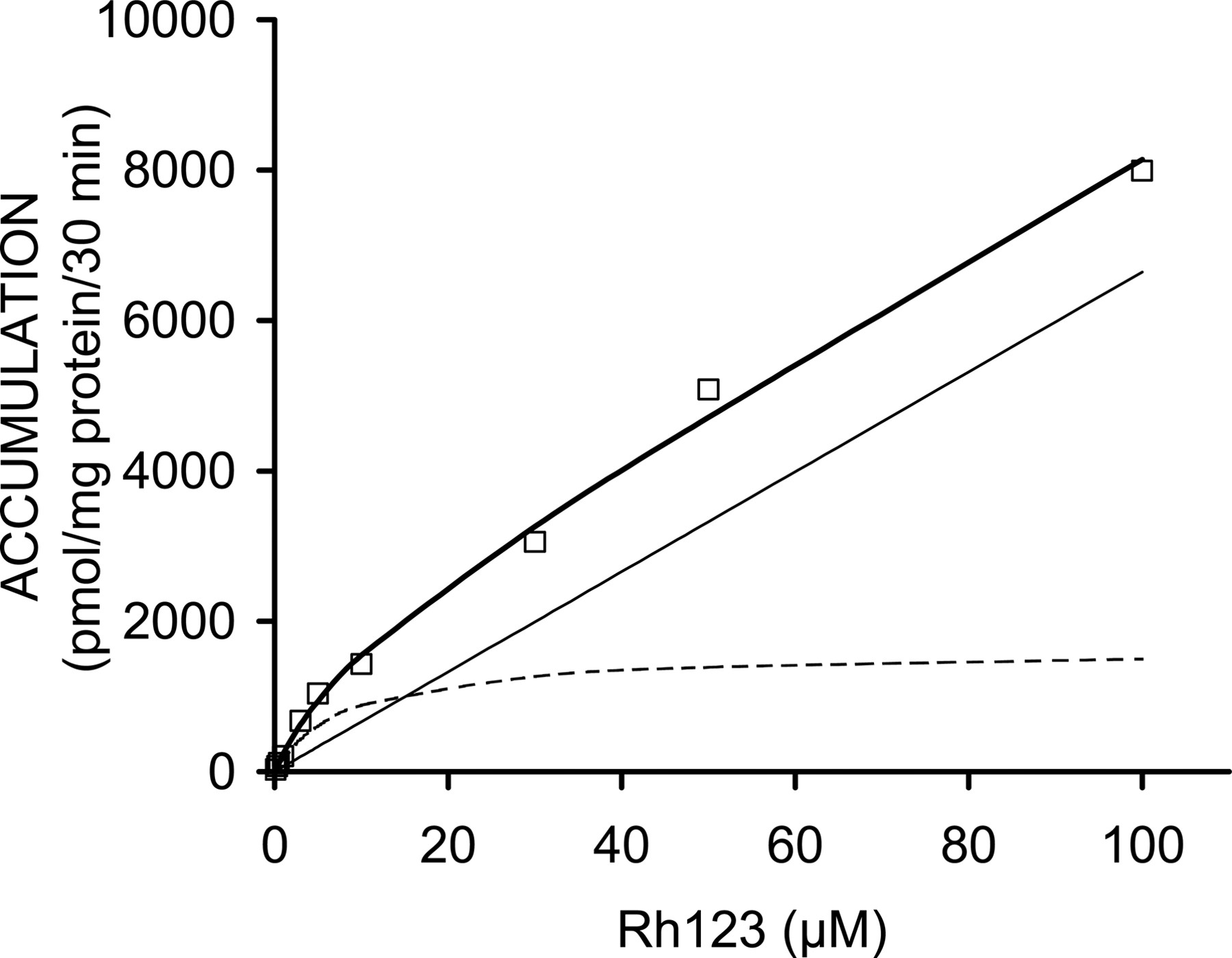
Concentration-Dependent Accumulation of Rhodamine 123. The concentration dependence of rhodamine 123 accumulation in sandwich-cultured rat hepatocytes is shown in Fig. 5. The accumulation data were described best by a model incorporating a linear (nonsaturable) and a Michaelis-Menten (saturable) component, with a Km of 8.4 μM and a Vmax of 54 pmol/(min · mg protein) for the saturable process. The rate constant (Kd) for nonsaturable uptake was 2.2 μl/(min · mg protein).

Concentration dependence of rhodamine 123 accumulation in sandwich-cultured rat hepatocytes (30 min). Open squares represent measured data for accumulation over 30 min in standard buffer. The thick solid line represents the best fit of a model (eq. 2) containing a saturable (dotted line) and a nonsaturable (thin solid line) component.
Rhodamine 123 Accumulation Was Reduced in the Presence of 1to100 μM Digoxin. When sandwich-cultured rat hepatocytes were coincubated with 1 μM rhodamine 123 and 1 to 100 μM concentrations of the P-glycoprotein and Oatp1a4 substrate, digoxin, rhodamine 123 BEI remained unaffected (see Fig. 6). However, the CLbile of rhodamine 123 showed a tendency to decrease by ∼40% at digoxin concentrations of 1 to 100 μM. Whereas the effect on CLbile failed to reach statistical significance, a significant effect was observed when rhodamine 123 accumulation values at various digoxin concentrations were compared (see Fig. 6). These results are consistent with the hypothesis that digoxin reduces the basolateral uptake of rhodamine 123, whereas the relative extent of rhodamine 123 canalicular excretion remains unaltered.

Effect of digoxin on hepatobiliary disposition of 1 μM rhodamine 123 in sandwich-cultured rat hepatocytes. Top panel, black and gray bars represent average rhodamine 123 accumulation following incubations in standard or Ca2+/Mg2+-free HBSS, respectively. Bottom panel, black and gray bars represent average rhodamine 123 BEI and biliary clearance values, respectively, in the presence of increasing digoxin concentrations. In both panels, bars for the control group and for the incubations at 30 μM digoxin represent average values across three hepatocyte preparations (average ± S.E.M.); bars at 1 μM and 100 μM digoxin represent average values obtained after quadruplicate incubations with hepatocytes from a single preparation (one liver). * denotes statistically significant difference from control value based on ANOVA/Dunnett's test (p < 0.05). (ns) denotes no statistically significant difference from control.
Rhodamine 123 Accumulation Was Modulated by Typical Oatp1a4 Substrates. The possible involvement of Oatp1a4 in the saturable uptake of rhodamine 123 in sandwich-cultured rat hepatocytes was evaluated further by measuring rhodamine 123 accumulation in standard buffer in the presence of 300 μM concentrations of the Oatp1a4 substrates d-verapamil, E217G, and quinine. Figure 7 illustrates that both d-verapamil and quinine reduced rhodamine 123 accumulation by ∼40%. The extent of reduction in rhodamine 123 accumulation was the same as that observed with 10 μM digoxin. Surprisingly, rhodamine 123 accumulation was enhanced more than 2-fold during coincubation with the Oatp1a4 substrate E217G.

Effect of typical Oatp1a4 substrates on accumulation of 1 μM rhodamine 123 into sandwich-cultured rat hepatocytes (standard conditions). Bars represent average (±S.E.M.; n = 4) values for accumulation over 30 min obtained with hepatocytes isolated from a single liver. * denotes significant difference from control value based on ANOVA/Dunnett's test (p < 0.05).
Discussion
Treatment of sandwich-cultured rat hepatocytes with known P-glycoprotein inhibitors (Fig. 1) resulted in reductions in both BEI and CLbile, without significantly affecting intracellular rhodamine 123 accumulation. These results are consistent with the previously reported effects of the potent P-glycoprotein inhibitor GF120918 (0.5–2 μM) on the in vitro biliary excretion of rhodamine 123 in sandwich-cultured rat hepatocytes (Annaert et al., 2001). Among the P-glycoprotein inhibitors tested in the present study, verapamil exhibited the strongest inhibition of rhodamine 123 secretion, whereas quinidine appeared to be the least efficient inhibitor of canalicular P-glycoprotein. Interestingly, comparable effects of rhodamine 123 BEI and CLbile were observed with 10 μM verapamil and 100 μM quinidine. These results are in agreement with the ∼10-fold higher P-glycoprotein affinity constant for quinidine (2.6 μM) as compared with verapamil (0.3 μM) in porated Caco-2 cells (Doppenschmitt et al., 1999).
Compared with P-glycoprotein inhibition, much less is known about the mechanisms of P-glycoprotein activation except for several reports describing putative stimulation of P-glycoprotein activity by flavonoids (Phang et al., 1993; Chieli et al., 1995; Conseil et al., 1998; Mitsunaga et al., 2000). The weak effect of quercetin on rhodamine 123 BEI observed in the present study appeared to be maximal at 25 μM, which is consistent with the “biphasic” effect of this flavonoid on vincristine accumulation in cultured mouse brain capillary endothelial cells reported previously by Mitsunaga et al. (2000). Intracellular rhodamine 123 accumulation in the presence of quercetin was enhanced much more than rhodamine 123 secretion into bile canaliculi, as reflected by the much larger effect of quercetin on CLbile values than on the BEI (relative measure of canalicular rhodamine 123 translocation). Higher intracellular rhodamine 123 accumulation would have resulted in a different interpretation (i.e., P-glycoprotein inhibition) of the effect of quercetin if unpolarized cells or hepatocytes cultured in a conventional configuration had been used. This result illustrates one advantage of studying hepatic transport processes in relevant cell models such as sandwich-cultured rat hepatocytes. Our results also are consistent with previous data generated by Chieli et al. (1995), showing that coincubation with flavonoids (including quercetin) had opposite effects on accumulation of the P-glycoprotein substrates rhodamine 123 and doxorubicin in cultured hepatocytes. The decreased accumulation of doxorubicin in the latter study was consistent with P-glycoprotein activation by flavonoids, and the increased accumulation of rhodamine 123 in the presence of quercetin (apparent P-glycoprotein inhibition) was attributed to a more important effect of quercetin on membrane fluidity, thus directly altering rhodamine 123 uptake and intracellular compartmentalization, despite the effects of P-glycoprotein activation.
To evaluate the possible effects of canalicular P-glycoprotein modulation on in vitro disposition of rhodamine 123 in sandwich-cultured rat hepatocytes, cells were cultured with modulators either for 72 h or for 48 h followed by a 24-h washout period prior to the determination of in vitro biliary excretion of rhodamine 123. After a 72-h treatment, only verapamil (1–10 μM) tended to increase the BEI and CLbile of rhodamine 123 (Fig. 3) in one hepatocyte preparation. However, the effect of verapamil disappeared when a 24-h washout was included (Fig. 4). This suggests that the effects of verapamil treatment that were observed after the 72-h treatment may be more related to P-glycoprotein activation rather than P-glycoprotein induction. Rifampicin significantly increased rhodamine 123 BEI and CLbile in one hepatocyte culture preparation when a 24-h washout was performed after the 48-h treatment period (Fig. 4). This finding appears to be consistent with some P-glycoprotein induction by rifampicin in sandwich-cultured rat hepatocytes. However, induction with rifampicin was not observed in two other hepatocyte preparations, which may be explained by variability in induction response across different hepatocyte cultures. In addition, it should be noted that the P-glycoprotein-inducing effects of rifampicin are reported to typically occur extensively in humans and other nonrodent species (Schuetz et al., 1996; Greiner et al., 1999), as opposed to rats and mice. On the other hand, Hanafy et al. (2001) reported reduced intestinal absorption of the P-glycoprotein substrate talinolol following rifampicin pretreatment in rats. The fact that rifampicin also has been reported to inhibit P-glycoprotein (Fardel et al., 1995) explains the complete blockage of rhodamine 123 biliary excretion when sandwich-cultured rat hepatocytes were cultured with 50 μM rifampicin for 72 h (without washout), which can be attributed to direct inhibition by rifampicin that had remained in the cells after treatment. The results illustrate the necessity to include a washout period to unambiguously evaluate compounds with both inducing and inhibiting properties. Treatment of the hepatocytes with 10 to 50 μM dexamethasone did not result in any significant effect on rhodamine 123 BEI. However, dexamethasone produced a 2- to 3-fold increase in the in vitro biliary clearance of rhodamine 123 in sandwich-cultured rat hepatocytes. This combination of increased biliary clearance and unchanged BEI values following treatment with dexamethasone indicates that dexamethasone primarily enhances accumulation of rhodamine 123 across the basolateral membrane. Although hepatic rhodamine 123 uptake is believed to occur via passive diffusion (Johnson et al., 1981; Altenberg et al., 1994; Chieli et al., 1995), induction of a putative transporter mediating hepatic sinusoidal uptake of rhodamine 123 could be a mechanism responsible for the effect of dexamethasone treatment. It is noteworthy that an influx transporter has been hypothesized to mediate rhodamine 123 uptake into a human cancer cell line (Cho et al., 2000) as well as rhodamine 123 transport across the basolateral membrane of Caco-2 cells (Troutman and Thakker, 2003). The data shown in Fig. 5 indicate that a saturable mechanism contributes substantially to the basolateral uptake of rhodamine 123 in the low micromolar concentration range. Among the various families of up-take transporters that are localized on the hepatic basolateral membrane, the organic anion-transporting polypeptide (Oatp) and organic cation transporter families are best characterized to date (Kusuhara and Sugiyama, 2001). Since organic cation transporters have been implicated primarily in the hepatic uptake of smaller organic cations, we postulated that one of the Oatp isoforms present in rat liver, particularly rat Oatp1a4, may play a role in rhodamine 123 uptake in sandwich-cultured rat hepatocytes. Cvetkovic et al. (1999) demonstrated previously that many P-glycoprotein substrates also are recognized by Oatp1a4. To test the hypothesis that Oatp1a4 is involved in rhodamine 123 uptake in sandwich-cultured rat hepatocytes, the effect of various concentrations of digoxin, a high-affinity Oatp1a4 substrate (Reichel et al., 1999), was examined. Figure 6 illustrates that coincubation of sandwich-cultured rat hepatocytes with 1 μM digoxin significantly decreased rhodamine 123 accumulation, whereas rhodamine 123 BEI was not significantly altered by digoxin concentrations up to 100 μM, suggesting that digoxin interacts primarily with basolateral rhodamine 123 uptake. The fact that canalicular P-glycoprotein-mediated rhodamine 123 transport was not inhibited by digoxin may be explained by the much lower intracellular accumulation of digoxin as compared with rhodamine 123 (Annaert et al., 2001). In addition, inhibition of P-glycoprotein by digoxin has not been reported, which at least partly may be due to the relatively low affinity of digoxin for P-glycoprotein (apparent EC50 = 58 μM in Caco-2 monolayers; Stephens et al., 2001). On the other hand, the effects of digoxin on basolateral rhodamine 123 transport can be explained by the very high affinity of digoxin for Oatp1a4 (Km = 0.2–1 μM; Noé et al., 1997; Sugiyama et al., 2002). The role of Oatp1a4 in rhodamine 123 transport was demonstrated further by studying the effects of other Oatp substrates (d-verapamil, E217G, and quinine) on basolateral rhodamine 123 uptake. As shown in Fig. 7, a 300 μM concentration of either d-verapamil or quinine and only 10 μM digoxin resulted in comparable inhibitory effects on rhodamine 123 accumulation. These results are in agreement with data from Dagenais et al. (2001), who examined the inhibition of [d-penicillamine2,5]-enkephalin transport, an Oatp1a4 substrate, by these compounds in rat brain. Interestingly, E217G enhanced rhodamine 123 accumulation by more than 2-fold, which is consistent with a report by Sugiyama et al. (2002), showing enhanced Oatp1a4-mediated transport of the Oatp1a4 substrate taurocholate in Oatp1a4-expressed LLC-PK1 cells in the presence of E217G. In the latter study, the existence of at least two substrate recognition sites on rat Oatp1a4 was proposed to explain this observation. In addition to the results from the coincubation experiments shown in Fig. 7, it is noteworthy that pretreatment of sandwich-cultured rat hepatocytes with 50 μM rifampicin also revealed reduced intracellular accumulation of rhodamine 123. This finding is consistent with data from Fattinger et al. (2000), illustrating inhibition of Oatp1a4 (Ki of 1.4 μM) but not Oatp1a1 by rifampicin in rat hepatocytes. Many of the effects observed in the present study with respect to changes in rhodamine 123 accumulation in sandwich-cultured rat hepatocytes also may be explained by effects on intracellular sequestration of rhodamine 123. However, the specific effects of digoxin and E217G on rhodamine 123 accumulation, as well as the inducing effect of dexamethasone, strongly support carrier-mediated transport of rhodamine 123, presumably by Oatp1a4, across the sinusoidal membrane of the rat hepatocyte. Consistent with these findings, Turncliff et al. (2004) recently demonstrated induction of Oapt1a4 in sandwich-cultured rat hepatocytes.
In conclusion, these results illustrate that sandwich-cultured rat hepatocytes are a promising in vitro tool for studying potential drug interactions (inhibition, activation, induction) at the level of hepatic transport. The ability of sandwich-cultured rat hepatocytes to form canalicular networks and exhibit cell polarity makes this model suitable for mechanistic studies and for reliably predicting the contribution of transport processes occurring on each membrane domain. In particular, the results obtained in this study revealed that some of the known P-glycoprotein modulators also altered rhodamine 123 accumulation into the hepatocyte, implicating a basolateral transporter mediating hepatic rhodamine 123 uptake. Future challenges involve demonstrating the potential of sandwich-cultured rat hepatocytes to assess the interplay between hepatic metabolism and transport, thus allowing the prediction of hepatobiliary disposition of drug metabolites. Finally, extending the sandwich technique to the culture of human hepatocytes seems promising for the ultimate in vitro prediction of hepatobiliary drug disposition in humans.
Footnotes
-
This study was supported by Grant R01 GM41935 from the National Institutes of Health. K.L.R.B. is co-founder of Qualyst, Inc. and a member of the Scientific Advisory Board for Qualyst, Inc.; Qualyst, Inc. has exclusively licensed the sandwich-cultured hepatocyte technology for quantification of biliary excretion (B-CLEAR).
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.104.001669.
-
ABBREVIATIONS: GF120918, N-(4-[2-(1,2,3,4-tetrahydro-6,7-dimethoxy-2-isoquinolinyl)ethyl]-phenyl)-9,10-dihydro-5-methoxy-9-oxo-4-acridine carboxamide; BEI, biliary excretion index (percentage); CLbile, biliary clearance (ml/min/kg); E217G, 17β-estradiol-d-17β-glucuronide; HBSS, Hanks' balanced salt solution; DMEM, Dulbecco's modified Eagle's medium; BSA, bovine serum albumin; FBS, fetal bovine serum; Oatp, organic anion-transporting polypeptide; ANOVA, analysis of variance.
-
↵1 Present address: Preclinical Pharmacokinetics, Johnson & Johnson Pharmaceutical Research & Development, a division of Janssen Pharmaceutica NV, Turnhoutseweg 30, B-2340 Beerse, Belgium.
- Received July 30, 2004.
- Accepted December 10, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}