


Abstract
We investigated the mechanism of renal tubular secretion of the dopamine D2 receptor agonist pramipexole in rats, focusing on organic cation transporters 1 and 2. The uptake of [14C]pramipexole by Xenopus oocytes injected with complementary RNA of either rat organic cation transporter (rOCT) 1 or rOCT2 was significantly higher than that by water-injected oocytes: the kinetic parameters, Km and Vmax, of pramipexole uptake were 49.5 μM and 234 pmol/60 min/oocyte for rOCT1, and 16.9 μM and 12.8 pmol/60 min/oocyte for rOCT2. Pramipexole was taken up into kidney slices in a time- and concentration-dependent manner, and Eadie-Hofstee plots revealed the involvement of two saturable components. The kinetic parameters, Km1 and Vmax1, of the high-affinity component were 12.9 μM and 10.7 nmol/15 min/g kidney, respectively. The uptake of [14C]pramipexole by rOCT1, rOCT2, and kidney slices was inhibited by procainamide and corticosterone, which are selective inhibitors of rOCT1 and rOCT2, respectively. The IC50 values of procainamide and corticosterone for the uptake of [14C]pramipexole by rOCT1, rOCT2, and kidney slices were 7.7, 167.0, and 47.0 μM and 163.7, 10.7, and 47.7 μM, respectively. These results demonstrate that both rOCT1 and rOCT2 are involved in the renal uptake of pramipexole across the basolateral membrane of the proximal tubular epithelial cells.
Conventional ergot-derived dopamine agonists, e.g., bromocriptine and pergolide, are widely used in the treatment of early-stage Parkinson's disease. However, the aminobenzothiazole compound pramipexole (Fig. 1), a potent dopamine autoreceptor agonist that stimulates members of the postsynaptic D2 receptor family (D2, D3, and D4), improving the symptoms of Parkinson's disease (Mierau and Schingnitz, 1992; Mierau et al., 1995; Domino et al., 1997), is being introduced for the treatment of both early- and advanced-stage Parkinson's disease. The absolute bioavailability of pramipexole after oral administration in humans is over 90%. The drug is excreted into urine via kidney, mainly (about 90%) as the unchanged form, and the renal clearance accounts for 80% of the total clearance (Wright et al., 1997; Hubble, 2000). The renal excretion of pramipexole involves both glomerular filtration and tubular secretion (Wright et al., 1997). A population pharmacokinetic analysis based on the data of Japanese patients indicated that the total clearance of pramipexole was decreased about 25% by coadministration of the Parkinson's disease agent amantadine (Yamamura et al., 2004). Amantadine, a cationic compound, is eliminated via renal tubular secretion (Wu et al., 1982) and is a substrate of rat organic cation transporter (rOCT)1 and rOCT2 (Goralski et al., 2002).

Chemical structure and position of the radiolabel (*) of pramipexole.
OCTs are a family of polyspecific organic cation transporters responsible for the transport of organic cations, including xenobiotics and endogenous compounds. The OCT family currently consists of three members: OCT1, OCT2, and OCT3 (Grundemann et al., 1994; Okuda et al., 1996; Schweifer and Barlow, 1996; Gorboulev et al., 1997; Wu et al., 1998a) and is distantly related to OCTN1 (Tamai et al., 1997) and OCTN2 (Tamai et al., 1998; Wu et al., 1998b), which are classified as organic cation/carnitine transporters. In rodents, OCT1 is expressed in liver, kidney, and small intestine (Grundemann et al., 1994; Schweifer and Barlow, 1996; Jonker et al., 2001), whereas in humans, OCT1 is primarily expressed in the liver (Gorboulev et al., 1997). OCT2 is predominantly expressed in the kidney in rodents and humans. In rat kidney, rOCTs are expressed in the basolateral membrane of the S1 and S2 segments (rOCT1) and S2 and S3 segments (rOCT2) of the proximal tubule, and they have a significant role in the first step of tubular secretion (Martel et al., 1999; Chen et al., 2002; Slitt et al., 2002). A study using OCT gene-disrupted mice demonstrated that the kidney-to-plasma concentration ratios of prototypic organic cation tetraethylammonium (TEA) in OCT1(-/-), OCT2(-/-), and OCT1/2(-/-) mice at steady state were 2-, 2-, and 6-fold lower, respectively, than that in wild-type mice, suggesting that both OCT1 and OCT2 are involved in the uptake of TEA by the kidney (Jonker et al., 2003).
We hypothesized that renal handling of pramipexole is mediated by OCT isoforms expressed on the kidney basolateral membrane. In the present study, we attempted to characterize pramipexole transport from blood to kidney proximal tubular cells by using rOCT1- and rOCT2-expressing Xenopus oocytes and rat kidney slices.
Materials and Methods
Chemicals. [14C]Pramipexole, (S)-2-amino-4,5,6,7-tetrahydro-6-propylaminobenzothiazole dihydrochloride monohydrate (58 mCi/mmol), and unlabeled pramipexole were synthesized by Boehringer Ingelheim GmbH (Ingelheim, Germany). [14C]Tetraethylammonium ([14C]TEA) and [14C]mannitol were purchased from American Radiolabeled Chemicals (St. Louis, MO) and Amersham Biosciences UK, Ltd. (Little Chalfont, Buckinghamshire, UK), respectively. Corticosterone, procainamide, TEA, and amantadine were purchased from Sigma-Aldrich (St. Louis, MO). All other chemicals and reagents were commercial products of reagent grade.
Transport Studies in Xenopus Oocytes. To express transporter molecules in Xenopus oocytes, pSPORT1 vector (Invitrogen, Carlsbad, CA) containing rOCT1, rOCT2, or rOCT3, each provided by Dr. V. Ganapathy (Medical College of Georgia, Atlanta, GA), was linearized with BamH1 or NotI. The sense complementary RNA (cRNA) was prepared by in vitro transcription with T7 RNA polymerase in the presence of ribonuclease inhibitor and RNA cap analog using a mMESSAGEmMACHINE kit (Ambion, Austin, TX). Mature (stage V-VI), defolliculated oocytes were selected from Xenopus laevis (Hamamatsu Creature Materials, Hamamatsu, Japan) and maintained in modified Barth's medium [88 mM NaCl, 1 mM KCl, 0.33 mM Ca(NO3)2, 0.41 mM CaCl2, 0.82 mM MgSO4, 2.4 mM NaHCO3, 10 mM HEPES, pH 7.4) containing 50 mg/l gentamycin. The next day, oocytes were injected with 50 nl of H2O/oocyte in the presence or absence of 25 or 50 ng of rOCT1, rOCT2, or rOCT3 cRNA. The oocytes were then incubated at 18°C in modified Barth's medium containing 50 mg/l gentamycin.
At 3 or 4 days after cRNA injection, the oocytes were transferred to a 24-well plate and preincubated in 0.5 ml of transport buffer (82.5 mM NaCl, 2.5 mM KCl, 1 mM Na2HPO4, 1 mM sodium pyruvate, 1 mM CaCl2, 5 mM HEPES, pH 7.4) for 30 min. After this preincubation, transport buffer (0.5 ml) containing a radiolabeled compound with or without an inhibitor was added to initiate the uptake reaction. The cells were incubated at room temperature for a designated time, and then washed three times with 1 ml of ice-cold transport buffer to terminate the uptake. The oocytes were solubilized with 5% SDS for 120 min. The radioactivity was measured using a liquid scintillation counter (Tri-Carb 2500TR; PerkinElmer Life and Analytical Sciences, Boston, MA) after the addition of 3 ml of scintillation cocktail, Clearsol II (Nacalai Tesque, Kyoto, Japan). Uptake was expressed as the cell-to-medium ratio (microliters/oocyte) obtained by dividing the uptake amount by the concentration of substrate in the transport buffer. To estimate the kinetic parameters for the uptake by rOCT1 and rOCT2, the initial uptake rates in rOCT1- and rOCT2-injected oocytes after subtraction of the uptake rate in water-injected oocytes were fitted to the following equation by means of nonlinear least-squares regression analysis using the MULTI program (Yamaoka et al., 1981): v = Vmax × s/(Km + s), where v is the initial uptake rate of substrate (pmol/60 min/oocyte), s is the substrate concentration in the medium (μM), Km is the Michaelis-Menten constant (μM), and Vmax is the maximum uptake rate (pmol/60 min/oocyte). The half-inhibitory concentration (IC50) values of inhibitors were obtained by examining their inhibitory effects on rOCT1- and rOCT2-mediated uptake of TEA or pramipexole, assuming competitive inhibition according to the following equation: C/M+I = C/M × (1 + I/IC50), where C/M represents the cell-to-medium ratio and the subscript (+I) represents the value in the presence of inhibitor. I is the concentration of inhibitor (μM). IC50 values were estimated by the nonlinear least-squares method using the MULTI program (Yamaoka et al., 1981).
Uptake by Kidney Slices. Uptake studies using kidney slices were carried out as described in a previous report (Urakami et al., 1999). The slices of whole kidneys were prepared with a microslicer (Zero 1; Dosaka EM, Kyoto, Japan), and the renal pelvis portion was removed. Slices (0.3 mm thick) of whole kidneys from male Sprague-Dawley rats were put in ice-cold oxygenated incubation buffer (120 mM NaCl, 16.2 mM KCl, 1 mM CaCl2, 1.2 mM MgSO4, and 10 mM NaH2PO4/Na2HPO4, adjusted to pH 7.5). Three slices, weighing 10 to 30 mg, were randomly selected and then preincubated in a 12-well plate with 1 ml of oxygenated incubation buffer in each well at 37°C for 5 min. After preincubation, the incubation buffer was replaced with incubation buffer containing [14C]pramipexole or [14C]TEA with or without an inhibitor to initiate the uptake reaction. The apparent uptake of [14C]mannitol was evaluated in independent experiments (n = 3) to estimate the volume of water adhering to the kidney slices during an incubation. The mean value of the adherent water volume was then used to correct apparent values for the uptake of [14C]pramipexole and [14C]TEA. The uptake studies were carried out at 37°C for an appropriate time, and then each slice was rapidly removed from the incubation buffer, washed in ice-cold saline, blotted on filter paper, weighed, and dissolved in 1 ml of Soluene 350 (PerkinElmer Life and Analytical Sciences). The radioactivity was measured using a liquid scintillation counter (Tri-Carb 2500TR; PerkinElmer Life and Analytical Sciences) after the addition of 5 ml of scintillation cocktail, Hionic Fluor (PerkinElmer Life and Analytical Sciences). Uptake was expressed as the slice-to-medium ratio (ml/g kidney) obtained by dividing the uptake amount by the concentration of substrate in the incubation buffer. To estimate the kinetic parameters for the uptake of pramipexole and TEA, the initial uptake rate was fitted to the following equation by means of nonlinear least-squares regression analysis using the MULTI program (Yamaoka et al., 1981): v = Vmax1 × s/(Km1 + s) + Vmax2 × s/(Km2 + s) + Pdif × s, where v is the initial uptake rate of substrate (nmol/15 min/g kidney), s is the substrate concentration in the medium (μM), Km is the Michaelis-Menten constant (μM), Vmax is the maximum uptake rate (nmol/15 min/g kidney), and Pdif is the diffusion constant (ml/15 min/g kidney). The IC50 values were obtained as described above.
Statistical Analysis. The two-tailed unpaired Student's t test was used to assess the significance of difference between two sets of data. Differences were considered to be statistically significant when P was <0.05.
Results
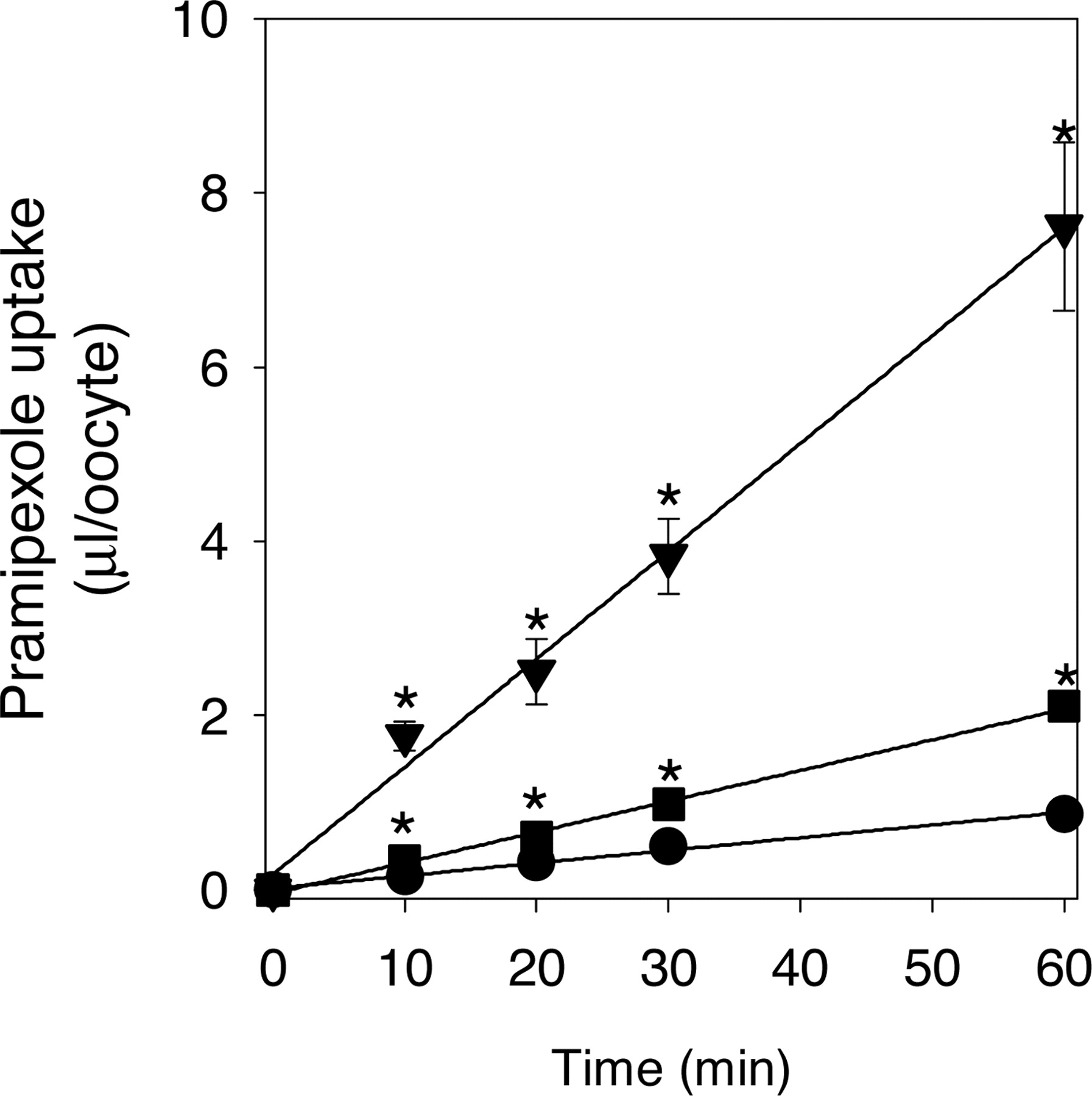
Uptake of pramipexole by Xenopus oocytes expressing rOCT1 or rOCT2. The time course of [14C]pramipexole uptake by Xenopus oocytes injected with rOCT1 cRNA, rOCT2 cRNA or water is shown in Fig. 2. [14C]Pramipexole was taken up into oocytes time-dependently, and the uptakes by oocytes expressing rOCT1 and rOCT2 were significantly higher than that by water-injected oocytes (p < 0.05). The initial uptake was assessed as the uptake at 60 min in the following kinetic and inhibition studies. The concentration dependence of the initial uptake of [14C]pramipexole by Xenopus oocytes expressing rOCT1 or rOCT2 after subtraction of the uptake by water-injected oocytes was studied in the concentration ranges of 5 μM to 1000 μM for rOCT1 and 5 μM to 100 μM for rOCT2 (Fig. 3). The Km and Vmax values of pramipexole transport by rOCT1 and rOCT2 were 49.5 ± 2.04 μM and 234 ± 5.27 pmol/60 min/oocyte, and 16.9 ± 2.54 μM and 12.8 ± 0.99 pmol/60 min/oocyte, respectively.

Time courses for the uptake of [14C]pramipexole by water-injected (circles), rOCT1 cRNA-injected (triangles), and rOCT2 cRNA-injected (squares) oocytes. The substrate concentration was 5 μM. Each point represents the mean ± S.E. from 5 to 10 oocytes, and ⋆ indicates a significant difference from the uptake of water-injected oocytes (P < 0.05).

Eadie-Hofstee plots for the initial uptake rates of [14C]pramipexole by rOCT1 cRNA-injected (A) and rOCT2 cRNA-injected (B) oocytes after subtraction of the initial uptake rate by water-injected oocytes. v, uptake in pmol/60 min/oocyte. v/[S], μl/60 min/oocyte. The substrate concentrations used were 5 to 1000 μM for rOCT1 and 5 to 100 μM for rOCT2. Each point represents the mean ± S.E. from 5 to 10 oocytes.
Uptake of Pramipexole and TEA by Kidney Slices.Figure 4, A and B, shows the time courses of the uptake of [14C]pramipexole and [14C]TEA, respectively, by kidney slices. Both [14C]pramipexole and [14C]TEA were transported into kidney slices in time-dependent manner. Taking into account the linearity of the plots, the uptake at 15 min was taken as the initial uptake in the following kinetic and inhibition studies. The concentration dependence of the uptake of pramipexole and TEA was studied in the concentration range of 1 μM to 5 mM. Eadie-Hofstee plots of pramipexole and TEA uptake are shown in Fig. 4, C and D, respectively. Analysis of the uptake of pramipexole and TEA revealed involvement of two saturable components and one nonsaturable component. The Km and Vmax values for pramipexole of the high- and low-affinity components were 12.9 ± 4.9 μM and 10.7 ± 3.7 nmol/15 min/g kidney, and 3347 ± 486 μM and 2884 ± 514 nmol/15 min/g kidney, respectively. The Km and Vmax values for TEA of the high- and low-affinity components were 101 ± 28.2 μM and 307 ± 72.2 nmol/15 min/g kidney, and 980 ± 209 μM and 1411 ± 199 nmol/15 min/g kidney, respectively. The Pdif values of the uptake of [14C]pramipexole and [14C]TEA were 0.39 ± 0.063 ml/15 min/g kidney and 0.049 ± 0.062 ml/15 min/g kidney, respectively.

Time profiles and Eadie-Hofstee plots of the uptake of [14C]pramipexole (A and C) and [14C]TEA (B and D) by rat kidney slices. A and B, the substrate concentration used was 5 μM. C and D, the uptake of [14C]pramipexole and [14C]TEA by kidney slices was measured at a concentration between 1 μM and 5 mM pramipexole and TEA for 15 min at 37°C. Adherent water volume was determined based on the uptake of [14C]mannitol by the three kidney slices, and the mean value of representative adherent water volume (0.22 ± 0.03, A and B; and 0.13 ± 0.003 ml/15 min/g kidney, C and D; mean ± S.E., n = 3) was used to correct the apparent uptake of [14C]pramipexole and [14C]TEA.
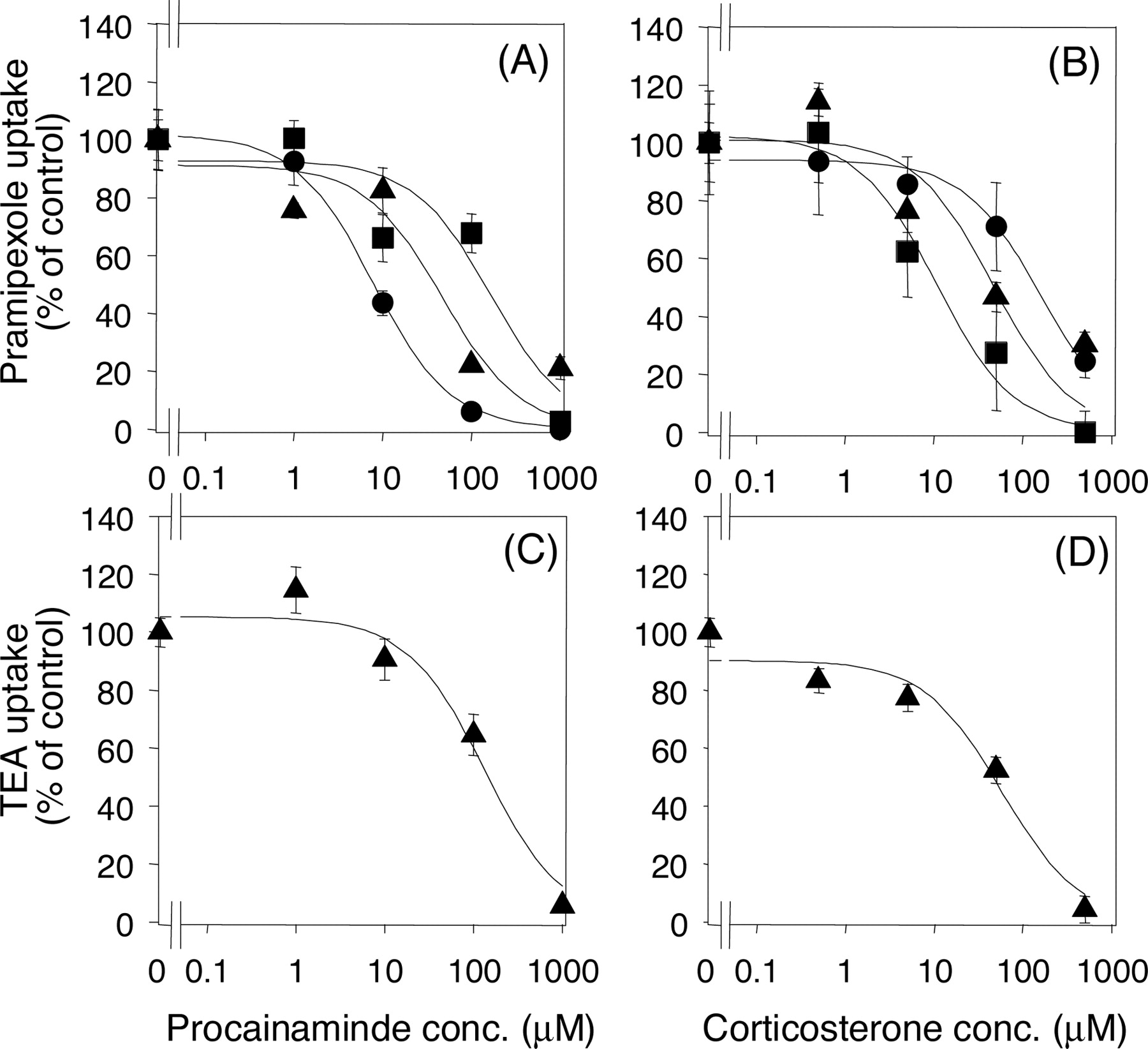
Inhibitory Effects of Procainamine and Corticosterone on [14C]Pramipexole and [14C]TEA Uptake by rOCT1- and rOCT2-Injected Oocytes and Kidney Slices. The involvement of rOCT1 and rOCT2 in the renal uptake of pramipexole and TEA was examined by using procainamide and corticosterone, which have differential affinities for rOCT1 and rOCT2, as inhibitors. The previously reported IC50 values of procainamide for TEA uptake by rOCT1 and rOCT2 were 20 and 445 μM, respectively, and those of corticosterone were 151 and 4 μM, respectively (Arndt et al., 2001). The concentration-dependent inhibitory effects of procainamide and corticosterone on the uptake of pramipexole and TEA by rOCT1, rOCT2, or kidney slices are shown in Fig. 5. The IC50 values of procainamide for the uptake of pramipexole by rOCT1, rOCT2, and kidney slices were 7.7, 167.0, and 47.0 μM, respectively (Table 1). The IC50 values of corticosterone were 163.7, 10.7, and 47.7 μM, respectively (Table 1). The IC50 values of procainamide and corticosterone for TEA uptake by kidney slices were 133.2 ± 42.2 and 59.0 ± 22.4 μM, respectively.
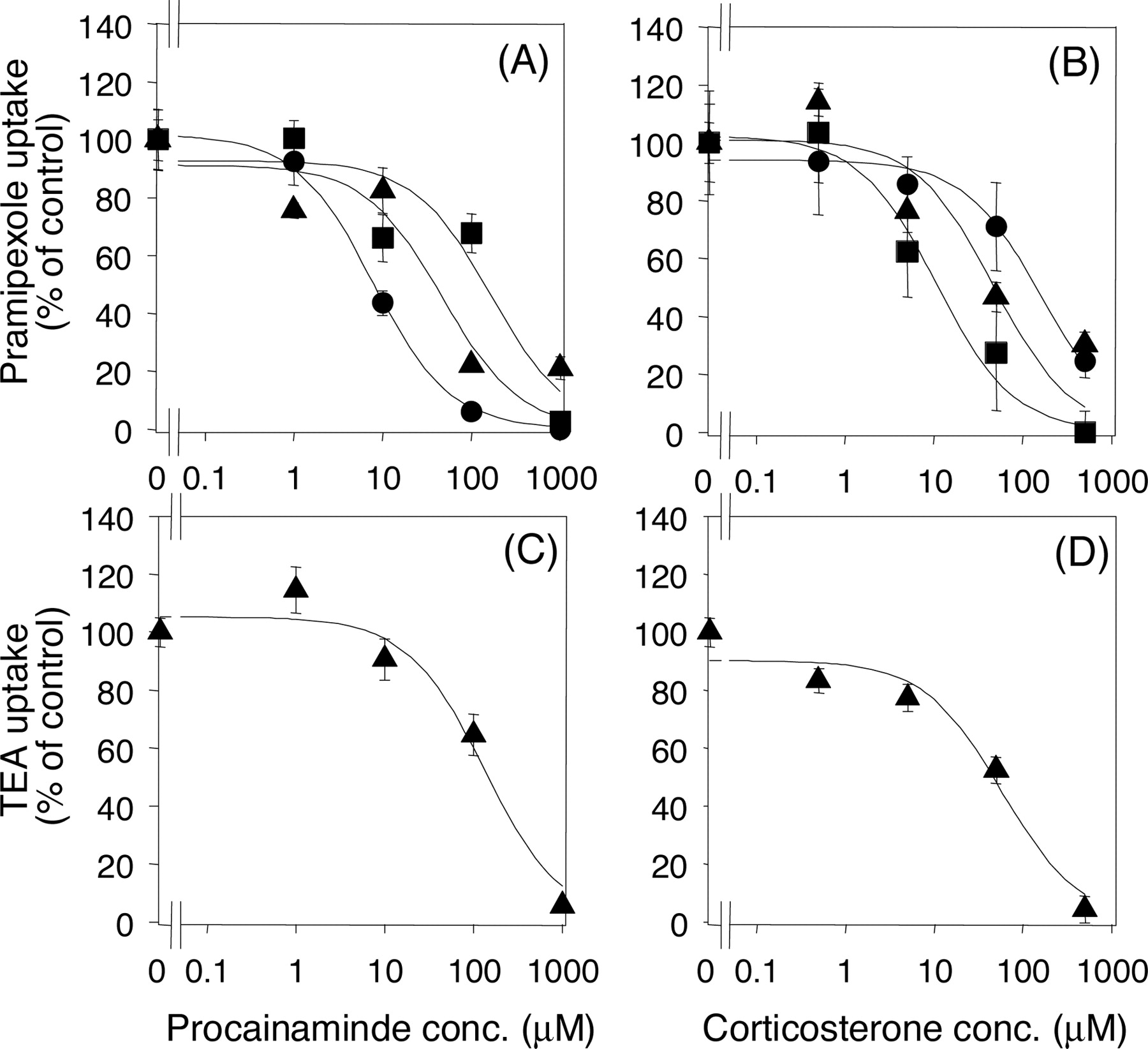
Inhibitory effects of procainamide and corticosterone on the uptake of [14C]pramipexole (A and B) and [14C]TEA (C and D) by rOCT1, rOCT2, and kidney slices. The saturable uptake of [14C]pramipexole and [14C]TEA in the presence of inhibitor was determined after subtraction of nonspecific uptake (evaluated as the uptake rate of the respective 14C-labeled drug in the presence of 1 mM unlabeled pramipexole or TEA). The values reported are expressed as a percentage of the corrected saturable uptake in the absence of any inhibitor. Circles, squares, and triangles represent the uptake by rOCT1, rOCT2, and kidney slices, respectively. The substrate concentration used was 5 μM. The solid line represents the fitted line obtained by nonlinear regression analysis. Each point represents the mean ± S.E. from 5 to 10 oocytes and 3 kidney slices.
The IC50 values of procainamide, corticosterone, and amantadine for [14C]pramipexole uptake Data shown in Figs. 5 and 6 were used to determine the IC50 values of procainamide, corticosterone, and amantadine for the uptake of pramipexole by rOCT1- and rOCT2-injected oocytes and rat kidney slices.
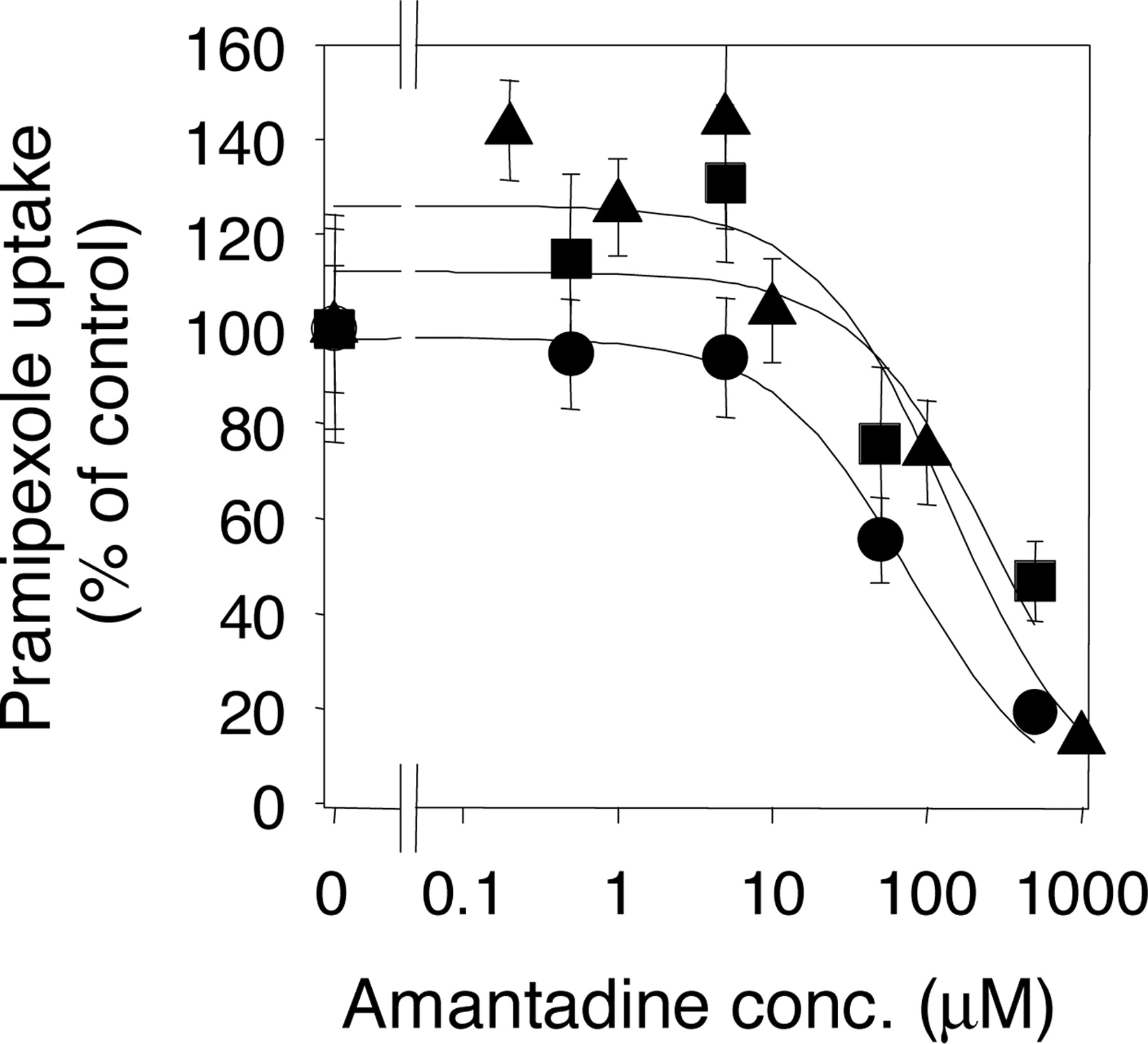
Inhibitory Effect of Amantadine on the Uptake of [14C] Pramipexole by rOCT1, rOCT2, and Kidney Slices. To confirm the involvement of rOCT1 and rOCT2 in the drug interaction between amantadine and pramipexole, we examined the inhibitory effect of amantadine on pramipexole uptake by rOCT1, rOCT2, and kidney slices (Fig. 6). The uptake of [14C]pramipexole by rOCT1, rOCT2, and kidney slices was inhibited by amantadine in a concentration-dependent manner, and the IC50 values were 76.3, 270.9, and 149.4 μM, respectively (Table 1).

Inhibitory effect of amantadine on the uptake of [14C]pramipexole by rOCT1, rOCT2, and kidney slices. The saturable uptake of [14C]pramipexole was expressed as the value after subtraction of the uptake rate in the presence of 1 mM pramipexole (nonspecific uptake). The values are expressed as a percentage of the saturable uptake in the absence of any inhibitor. Circles, squares, and triangles represent the uptake by rOCT1, rOCT2, and kidney slices, respectively. The substrate concentration used was 5 μM. The solid line represents the fitted line obtained by nonlinear regression analysis. Each point represents the mean ± S.E. from 5 to 10 oocytes and 3 kidney slices.
Discussion
Pramipexole is a full agonist with high selectivity for the D2 dopamine receptor family (Mierau and Schingnitz, 1992; Mierau et al., 1995; Domino et al., 1997) and is being introduced for the treatment of both early and advanced Parkinson's disease. After oral administration in humans, pramipexole is absorbed well and is mainly excreted in the urine as the unchanged form (Wright et al., 1997; Hubble, 2000). A population pharmacokinetic analysis in Japanese patients demonstrated that coadministration of amantadine decreased the total clearance of pramipexole by 25% (Wu et al., 1982; Goralski et al., 2002; Yamamura et al., 2004). In kidney, members of the organic cation transporter OCTN and OCT families are expressed at the apical and basolateral membranes, respectively (Urakami et al., 1998; Pietig et al., 2001; Tamai et al., 2001, 2004), and are involved in the urinary excretion of cationic compounds. Among them, OCT isoforms such as rOCT1, rOCT2, and human OCT2 are expressed on the basolateral membrane of proximal tubular cells and are important in the first step of renal clearance of cationic drugs. Thus, we hypothesized that OCT isoforms that are expressed on the basolateral membrane of kidney tubular cells take up pramipexole into the cells. Here, we have characterized pramipexole transport from blood to kidney proximal tubular cells by using kidney slices, and we examined the involvement of rOCT1 and rOCT2 transporters by using Xenopus oocytes expressing these transporters to identify the underlying mechanism of the drug interaction between pramipexole and amantadine.
The uptake of pramipexole by Xenopus oocytes expressing rOCT1 or rOCT2 was significantly higher than that by oocytes injected with water alone. The uptake of pramipexole showed both time and concentration dependence, and the results confirmed that pramipexole is a substrate of both rOCT1 and rOCT2 transporters (Figs. 2 and 3).
The uptake of [14C]pramipexole and [14C]TEA by rat kidney slices increased with time and was saturable, demonstrating that transporters are involved (Fig. 4). The Km values for the uptake of pramipexole by rOCT1, rOCT2, and rat kidney slices were 49.5 μM, 16.9 μM, and 12.9 μM, respectively. In the case of the typical organic cation TEA, the Km value for the uptake by rat kidney slices was 101 μM, which is almost the same as the reported Km values of TEA uptake by rOCT1 (95-129 μM) and rOCT2 (91 μM) expressed in Xenopus oocytes (Arndt et al., 2001). Thus, the Km values of organic cations transported by rOCT1 and rOCT2 seem to be similar to those obtained in the kidney slice studies. In particular, a good agreement of the Km values for the uptake of pramipexole among rOCT1, rOCT2, and kidney slices (high-affinity component) was observed, indicating that pramipexole is taken up by the kidney via rOCT1 and rOCT2.
To confirm the contributions of rOCT1 and rOCT2 to the high-affinity component of the renal uptake of pramipexole, we examined the inhibitory effect of procainamide and corticosterone using rOCT1- and rOCT2-expressing oocytes and rat kidney slices. Procainamide and corticosterone are putatively selective inhibitors of rOCT1 and rOCT2, respectively, because there are clear differences in the IC50 values of these drugs toward rOCT1- and rOCT2-mediated TEA transport (20 ± 11 versus 445 ± 262 μM for procainamide and 151 ± 26 versus 4 ± 0.7 μM for corticosterone) (Arndt et al., 2001). The obtained IC50 values of procainamide for the rOCT1- and rOCT2-mediated transport of pramipexole were 7.7 and 167.0 μM, respectively, and those of corticosterone were 163.7 and 10.7 μM, respectively (Table 1). This result clearly shows that these compounds can be selective inhibitors of rOCT1 and rOCT2 if used at appropriate concentrations. If pramipexole and TEA are transported by renal epithelial cells via only rOCT1 or rOCT2, the IC50 values in kidney slices should be comparable with those of rOCT1- or rOCT2-injected oocytes. However, the IC50 values in rat kidney slices were between those of rOCT1- and rOCT2-injected oocytes. Furthermore both rOCT1 and rOCT2 are expressed in the rat kidney (Thomas et al., 2003), supporting the idea that both rOCT1 and rOCT2 are involved in the uptake of pramipexole and TEA by rat kidney slices. The results of our inhibition study with procainamide and corticosterone on the uptake of TEA by rOCT1, rOCT2, and rat kidney slices are in good agreement with the above results. The two different approaches, i.e., kinetic and specific inhibition studies using the Xenopus oocyte gene expression system and kidney slices, both indicate that pramipexole is taken up by the kidney via both rOCT1 and rOCT2 in a manner similar to TEA.
It is noteworthy that there was an ∼12-fold difference in the values of transport efficiency, Vmax/Km (9.45 μl/60 min/oocyte for rOCT1, 0.76 μl/60 min/oocyte for rOCT2) for the uptake of pramipexole by rOCT1- and rOCT2-injected oocytes, after correction for the amount of injected cRNA. Without information on the numbers of functionally expressed transporter proteins in oocyte, the transport efficiency of rOCT1 and rOCT2 cannot be compared directly. If it is assumed that the expression efficiency of functional protein from cRNA is the same between rOCT1 and rOCT2, it is suggested that pramipexole is more efficiently transported by rOCT1 than by rOCT2. In addition, TEA is transported 3 times more efficiently by rOCT1 than by rOCT2 (our data; Arndt et al., 2001). These results apparently conflict with the finding that TEA and pramipexole are transported by both rOCT1 and rOCT2 in kidney slices. This issue may be explained by the different expression levels of rOCT1 and rOCT2 in the rat kidney. There is little information about the absolute expressed amount of protein of rOCT1 and rOCT2 in kidney. Although there may be a difference in amplification efficiency, the mRNA expression of rOCT2, determined by branched DNA signal amplification assay using multiple hybridization sequences in the probe sets for each transporter, was about 4 times higher than that of rOCT1 (Slitt et al., 2002). The difference in the expression levels of rOCT1 and rOCT2 may compensate for the difference in the transport efficiencies (Vmax/Km) for the uptake of pramipexole and TEA by rOCT1 and rOCT2.
The Eadie-Hofstee plots for the uptake of pramipexole and TEA by kidney slices indicated that multiple transporters are involved. The basolateral membrane of kidney tubules contains rat organic anion transporters (OAT) 1 and 3 as well as rOCT3 transporter (van Montfoort et al., 2003). It is reported that TEA is transported by rOCT3 but not by rOAT1 and 3 (Sekine et al., 1998; Uwai et al., 1998; Kusuhara et al., 1999). The Km value of the uptake of TEA by rOCT3 is 2500 μM (Kekuda et al., 1998), which is close to the Km value of the low-affinity component obtained in the present kidney slice experiments. Therefore, it is possible that rOCT3 is involved in the uptake of TEA as the low-affinity component observed in kidney slices. The uptake of [14C]pramipexole by rOCT3-injected oocytes (0.51 ± 0.068 μl/30 min/oocyte) was significantly higher than that by water-injected oocytes (0.34 ± 0.043 μl/30 min/oocyte) (p < 0.05, data not shown). Furthermore, 100 μM and 1 mM pramipexole inhibited the uptake of p-aminohippurate by rat kidney slices to 84 and 74% of that without pramipexole (data not shown), indicating that pramipexole has a low affinity for OAT isoforms expressed in the kidney. Based on these results, it is possible that pramipexole is taken up by the kidney via rOCT3, rOAT1, or rOAT3 at a concentration about 5000 times greater than the clinically relevant plasma concentration (Hubble, 2000), indicating that the low-affinity component has little role in the transport of pramipexole at the clinical concentration.
To investigate the involvement of rOCT1 and rOCT2 in drug interaction between pramipexole and amantadine, we examined the inhibitory effect of amantadine on the uptake of pramipexole by rOCT1, rOCT2, and kidney slices. Amantadine inhibited the uptake of pramipexole by rOCT1- and rOCT2-expressing oocytes and rat kidney slices in a concentration-dependent manner, suggesting that an organic cation-specific mechanism is at least partly involved in this drug interaction. However, the IC50 values of amantadine for pramipexole uptake by rOCT1, rOCT2, and rat kidney slices were more than 10 times higher than the clinically relevant concentration (Hayden et al., 1985). To elucidate the mechanism underlying the drug interaction between pramipexole and amantadine in humans, additional studies, e.g., to determine the affinity of pramipexole for human OCTs, will be needed.
In conclusion, the present study has demonstrated that both rOCT1 and rOCT2 are involved in the renal uptake of pramipexole across the basolateral membrane of the proximal tubular epithelial cells.
Footnotes
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.104.002519.
-
ABBREVIATIONS: r, rat; OCT, organic cation transporter; cRNA, complementary RNA; OAT, organic anion transporter; TEA, tetraethylammonium.
- Received September 27, 2004.
- Accepted January 3, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}