


Abstract
The common marmoset (Callithrix jacchus), a New World primate species, is potentially a useful animal model for preclinical studies in drug development. However, cytochrome P450 (P450) enzymes have not been fully identified and characterized in marmosets. In this study, we identified P450 2A6 cDNA with the sequence highly identical (91–94%) to human P450 2A6, 2A7, and 2A13 cDNA and cynomolgus monkey P450 2A23, 2A24, and 2A26 cDNA. Among the tissue types examined, marmoset P450 2A6 mRNA was most abundantly expressed in livers where P450 2A6 protein was also detected by immunoblotting. Phylogenetic analysis showed that marmoset P450 2A6 was more closely clustered with human and cynomolgus monkey P450 2As than P450 2As of dog, rat, and mouse (the species also used in drug metabolism). Marmoset P450 2A6 heterologously expressed in Escherichia coli membranes efficiently catalyzed 7-ethoxycoumarin O-deethylation, similar to human P450 2A6 and 2A13 and cynomolgus monkey P450 2A23, 2A24, and 2A26, but much less effectively coumarin 7-hydroxylation, showing some difference as well. Interestingly, marmoset P450 2A6 and cynomolgus monkey P450 2A23 catalyzed phenacetin O-deethylation, which is catalyzed by human P450 1A2 and 2A13, but not by P450 2A6. Marmoset P450 2A6 also exhibited catalytic activity toward testosterone by the multiple sites, but not rat P450 2A-specific testosterone 7α-hydroxylation activity. These results indicated that marmoset P450 2A6 had functional characteristics different from those of human and cynomolgus monkey P450 2As in terms of partially different substrate specificities and catalytic activities, indicating its importance of further studies for P450 2A-dependent drug metabolism in marmosets.
Introduction
Nonhuman primate species, especially the Old World monkey such as cynomolgus monkeys (Macaca fascicularis) and rhesus monkeys (Macaca mulatta), are widely used for preclinical assessment of metabolism, pharmacokinetics, and toxicity of drugs because of their genetic closeness and phenotypic similarities to humans. The common marmoset (Callithrix jacchus), a New World monkey, has benefits such as small body size, ease handling, high fertility, and early sexual maturity (Orsi et al., 2011). Therefore, marmosets have attracted considerable attention as a useful animal model in various fields such as neuroscience, stem cell research, drug metabolism, toxicology, and immune and autoimmune diseases (Sasaki et al., 2005; Orsi et al., 2011; Carrion and Patterson, 2012; Jagessar et al., 2013; Kishi et al., 2014)
Cytochrome P450s (P450s) are the major enzymes that play a role in the metabolism of various endogenous and exogenous compounds, such as steroids, environmental carcinogens, and drugs. Human P450s contains 57 functional genes and 58 pseudogenes, including those of the P450 1–3 subfamilies important for drug metabolism. The human P450 2A subfamily comprises P450 2A6, 2A7, and 2A13 (Fernandez-Salguero and Gonzalez, 1995) with their amino acid sequences being highly homologous to each other (92–94%). Human P450 2A6, mainly expressed in the liver, is a major catalyst of coumarin and precarcinogens, including many N-nitrosamines, 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone, and aflatoxin B1 (Yamazaki et al., 1992; Koskela et al., 1999; Su et al., 2000; He et al., 2006). Human P450 2A7 mRNA expression has been shown in the liver, but heterologously expressed P450 2A7 showed no catalytic activity (Ding et al., 1995). Human P450 2A13 mRNA has been shown to be highly expressed in the lungs, and the respiratory tract such as nasal mucosa, and shares several substrates with P450 2A6 (Su et al., 2000). In macaques, more than 20 P450s were identified and characterized (Uno et al., 2011). The cynomolgus monkey P450 2A subfamily consists of P450 2A23, 2A24, and 2A26, which have high sequence identities (89–95%) to human P450 2As at amino acid level. As with human P450 2A6, cynomolgus monkey P450 2A23, 2A24, and 2A26 mRNAs were expressed predominantly in the liver; their proteins catalyzed coumarin 7-hydroxylation (Uno et al., 2007; Uehara et al., 2010).
Species differences for drug metabolism are an important issue for drug development. Marmosets and rats are reportedly poor coumarin 7-hydroxylators, unlike mice, cynomolgus monkeys, and humans (Lake et al., 1989; Steensma et al., 1994), indicating the differences of P450 2A enzymes in marmosets as compared with cynomolgus monkeys and humans. Marmoset P450 2A enzymes expressed in liver could account for the lower coumarin 7-hydroxylation activity in marmosets, similar to rats. However, in marmosets, P450 2A isoforms have not been identified to date, although we previously reported P450 2A gene was expressed in marmoset liver, similar to human livers (Shimizu et al., 2014). In this study, therefore, we isolated novel P450 2A6 cDNA from marmoset liver and analyzed its sequence identity, gene structure, tissue expression pattern, and drug-metabolizing activity and compared it with corresponding human and cynomolgus monkey P450 2As.
Materials and Methods
Chemicals and Tissue Samples.
Coumarin, 7-hydroxycoumarin, phenacetin, acetaminophen, and testosterone were purchased from Wako Pure Chemicals (Osaka, Japan). 6β-Hydroxytestosterone and 7-ethoxycoumarin were purchased from Sigma-Aldrich (St. Louis, MO). Oligonucleotides were synthesized by Sigma-Aldrich (Tokyo, Japan). Pooled liver microsomes from marmosets, cynomolgus monkeys, and humans were purchased from Corning Life Sciences (Woburn, MA). Brain, lung, liver, kidney, and small intestine were collected from 16 marmosets (nine males and seven females, 2–6 years of age) at the Central Institution for Experimental Animals (Kawasaki, Japan). The study was approved by the Institutional Animal Care and Use Committee. Liver microsomes from individual marmosets were prepared as described previously (Uehara et al., 2014). All other chemicals of highest grade commercially available were obtained.
cDNA Cloning of Marmoset P450 2A6.
Marmoset P450 2A6 cDNA was isolated by reverse transcription (RT)-polymerase chain reaction (PCR) using total RNA from liver tissue. Briefly, total RNA from the marmoset liver was extracted with RNeasy Mini Kit (Qiagen, Valencia, CA). The first-strand cDNA was prepared from the total RNA (1 μg) by RT using SuperScript III RT reverse transcriptase (Invitrogen, Carlsbad, CA) and oligo(dT) primers, according to the manufacturer’s instructions. The PCR was performed with KOD-Plus-Neo DNA polymerase (Toyobo, Osaka, Japan) on an ABI GeneAmp PCR System 9700 thermocycler (Applied Biosystems, Foster City, CA). The primers used, 5′-CCAAAGTCCGCAGTATAAAGGCAAAC-3′ and 5′-CTGCACTGAAACAGCCCTCACTCA-3′, were designed to amplify the entire coding region of a gene sequence highly identical to human P450 2A6 that was found in the marmoset genome using BLAT (University of California Santa Cruz Genome Bioinformatics). Thermal cycler condition included an initial 2-minute denaturation at 94°C, followed by 35 cycles of the following: 98°C for 10 seconds, 65°C for 30 seconds, and 68°C for 2 minutes, with a final 5-minute extension at 68°C. The amplified fragments were cloned into pCR4 vectors using Zero Blunt TOPO PCR Cloning Kit for Sequencing (Invitrogen). The insert of each recombinant clone was sequenced using ABI PRISM BigDye Terminator v3.0 Ready Reaction Cycle Sequencing Kit (Applied Biosystems) with an ABI PRISM 3730 DNA Analyzer (Applied Biosystems).
Determination of Marmoset P450 2A6 Exon 4 Sequences.
Marmoset P450 2A6 gene sequences except around exon 4 were found in the marmoset genome using BLAT. Marmoset P450 2A6 exon 4 sequences were amplified by PCR to determine exon-intron structure. PCR was carried out using AmpliTaq Gold 360 DNA Polymerase (Applied Biosystems) under the following conditions: 10 minutes at 95°C, followed by 10 seconds at 95°C, 20 seconds at 60°C, and 2 minutes at 72°C for 30 cycles. The primers 5′-CTTCTCCATCACCACCCTGC-3′ and 5′-AACTGGGTAGGGAGCATCTG-3′ were designed to amplify the exon 4 sequences, based on the marmoset P450 2A6 gene sequence. PCR products were cloned into pGEM-T easy vectors using pGEM-T Easy Vector System (Promega, Madison, WI), and sequenced, as described earlier.
Quantitative Real-Time RT-PCR.
The expression levels of marmoset P450 2A6 mRNA in brains, lungs, livers, kidneys, and small intestines were measured by real-time RT-PCR. RNA was extracted from the five tissues using RNeasy Mini Kit, as described earlier. An equal amount of total RNA was pooled from eight male marmosets for each tissue type. For each tissue type, pooled total RNA (1 µg) was reverse transcribed using SuperScript III RT reverse transcriptase and random primers (Invitrogen) in a final volume of 20 µl, according to manufacturer’s instructions. The RT products were used for real-time PCR in reaction mixture (20 µl), which was performed with an ABI PRISM 7500 sequence detection system (Applied Biosystems). The PCR primers were 5′-CAGTTTAAGAAGAGTGATGCATTTGTG-3′ and 5′-GATGGTGGTGAGGAACAGAAAGA-3′. Thermal cycler conditions were 2 minutes at 50°C and 10 minutes at 95°C, followed by 40 cycles at 95°C for 15 seconds and at 60°C for 1 minute. All samples were analyzed in triplicate. Data were normalized to 18S rRNA level to determine relative expression levels of marmoset P450 2A6 mRNA.
Immunoblotting.
The expression of P450 2A6 protein in marmoset liver microsomes was confirmed by immunoblotting, as described previously (Uehara et al., 2011), with some minor changes, as follows. In brief, recombinant P450 protein (0.1 pmol P450/lane) or tissue microsomal fractions from brains, lungs, livers, kidneys, and small intestines (15–100 μg/lane) were run on 10% sodium dodecyl sulphate–polyacrylamide gel, and the resolved proteins were transferred onto polyvinylidene difluoride membrane (Merck Millipore, Billerica, MA). The membranes were blocked with 0.5% skim milk at room temperature for 30 minutes, incubated with anti-human P450 2A6 antibodies (1:5,000; Nosan, Yokohama, Japan) at room temperature for 1 hour, and then with rabbit anti-mouse secondary antibodies (1:10,000; Santa Cruz Biotechnology, Santa Cruz, CA) at room temperature for 20 minutes. P450 2A protein bands were visualized by an ECL Prime Western Blotting Detection System (GE Healthcare, Buckinghamshire, UK), according to the manufacturer’s instructions.
Heterologous Expression of P450 2A Protein in Escherichia coli.
For enzyme assays, recombinant marmoset P450 2A6 was prepared by heterologous expression in Escherichia coli using expression plasmids. The N terminus modification was carried out by PCR, as described previously (Uehara et al., 2010), using the primers 5′-GGAATTCCATATGGCTCTGTTATTAGCAGTTTTTGTGGCCTTGCTGGCCTG-3′ and 5′-GCTCTAGACTCAGCGGGGCAGGAAGC-3′. PCR product was subcloned into the pCW vector containing human NADPH-P450 reductase cDNA via the cloning sites NdeI and XbaI (underlined). The sequences of the inserts were verified by DNA sequencing, as described earlier. The expression plasmids of human P450 2A6 and 2A13 and cynomolgus monkey P450 2A23, 2A24, and 2A26 were prepared, as described previously (Yamazaki et al., 2002; Fukami et al., 2008; Uehara et al., 2010). The membrane fractions were prepared as described previously (Yamazaki et al., 2002). The concentrations of P450 protein and NADPH-P450 reductase were estimated, as described previously (Phillips and Langdon, 1962; Omura and Sato, 1964).
Enzyme Assays.
7-Ethoxycoumarin O-deethylation, coumarin 7-hydroxylation, and phenacetin O-deethylation by recombinant proteins (marmoset P450 2A6, human P450 2A6 and 2A13) and marmoset and human liver microsomes were measured, as described previously (Yamazaki et al., 2002), with some minor changes, as follows. In brief, a typical incubation mixture (0.25 ml) contained recombinant protein (5 pmol) or liver microsomes (0.125 mg/ml), a NADPH-generating system (0.25 mM NADP+, 2.5 mM glucose 6-phosphate, and 0.25 U/ml glucose 6-phosphate dehydrogenase), and substrate (1–800 μM 7-ethoxycoumarin, 1–1000 μM coumarin or phenacetin) in 100 mM potassium phosphate buffer (pH 7.4). The reaction was started by adding NADPH, incubated at 37°C for the optimal time (7-ethoxycoumarin and coumarin, 10 minutes; phenacetin, 20 minutes), and terminated by addition of 25 μl 60% HClO4 (w/v) or 0.1 M HCl. Reaction mixtures were centrifuged at 10,000g for 5 minutes, and supernatants were analyzed directly by high-performance liquid chromatography with a fluorescence or a UV detector. To determine Km and Vmax values, data were fitted to an integrated form of the Michaelis-Menten equation using Kaleidagraph (Synergy Software, Reading, PA). All the data of enzyme activity were analyzed using the mean of duplicated determinations.
Bioinformatics.
The homology search was carried out using BLAST (National Center for Biotechnology Information). The multiple alignment of sequences was conducted using a Genetyx system (Software Development, Tokyo, Japan). The phylogenetic tree was created with DNASIS Pro (Hitachi Software, Tokyo, Japan). The P450 amino acid sequences from GenBank used for the analysis included the following: human P450 2A6 (NP_000753), 2A7 (NP_000755), 2A13 (NP_000757), and 2C9 (NP_000762); cynomolgus monkey P450 2A23 (NP_001272197), 2A24 (NP_001272277), and 2A26 (NP_001270957); rhesus monkey P450 2A23 (NP_001035306), 2A24 (NP_001035305), and 2A26 (NP_001191360); dog P450 2A7 (NP_001041492), 2A13 (NP_001032422), and 2A25 (ABB43281); rabbit P450 2A10 (NP_001164520) and 2A11 (NP_001164519); rat P450 2A1 (NP_036824), 2A2 (NP_036825), 2A3 (NP_036674), 2A4 (NP_034127), and 2A5 (NP_031838); and mouse P450 2A12 (NP_598418) and 2A22 (NP_001094937). The marmoset P450 2A6 amino acid sequence was deduced from the cDNA identified in this study.
Results
Molecular Cloning and Sequence Analysis of Marmoset P450 2A6 cDNA.
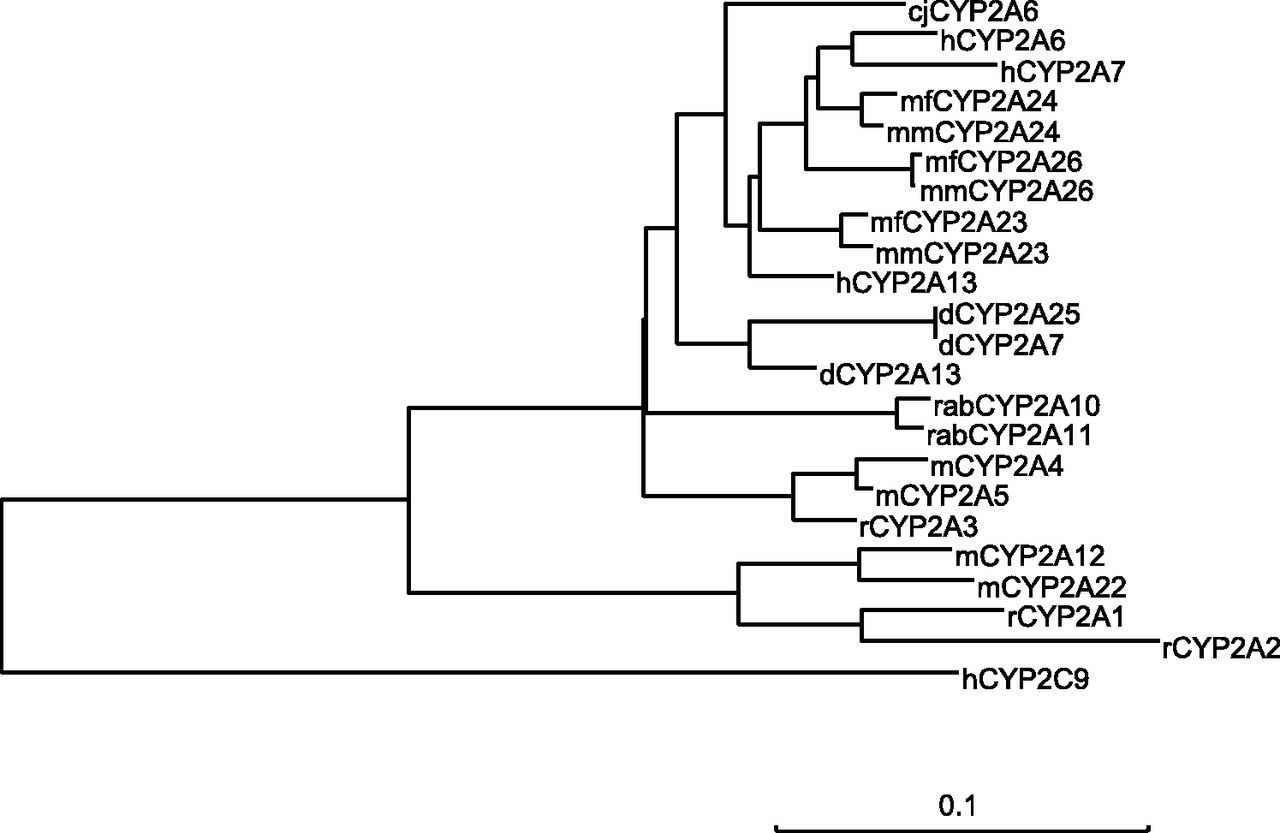
By analyzing the marmoset genome data using BLAT, the novel marmoset P450 2A (P450 2A6) gene showing a high sequence identity to human P450 2A6 cDNA was found. By RT-PCR using gene-specific primers, marmoset P450 2A6 cDNA (1485 bp) containing the entire coding region was isolated from livers. The deduced amino acids (494 residues) contained the primary sequence structure characteristic of P450 proteins, such as substrate recognition sites and the heme-binding region (Fig. 1). By comparing P450 2A cDNA sequences in marmoset and other species, marmoset P450 2A6 showed the highest sequence identity to human P450 2A13 (94%), followed by human P450 2A6 (93%) and 2A7 (91%) (Table 1). Marmoset P450 2A6 cDNA showed higher sequence identities to P450 2A cDNAs of cynomolgus monkeys (93%), rhesus monkeys (93%), and humans (91–94%) than those of dogs, rabbits, rats, and mice. Phylogenetic analysis of P450 2A amino acid sequences in marmosets and other species indicated that marmoset P450 2A6 was closely clustered with human P450 2As, together with cynomolgus monkey and rhesus monkey P450 2As, unlike dog, rabbit, rat, and mouse P450 2As (Fig. 2).

Alignment of the deduced amino acid sequences of marmoset P450 2A6. Marmoset (cj) P450 2A6 amino acid sequences were aligned with cynomolgus monkey (mf) P450 2A23, 2A24, and 2A26, and human (h) P450 2A6, 2A7, and 2A13 amino acid sequences. The putative heme-binding region and the putative substrate recognition site (SRS) are shown by broken and solid lines, respectively. Identical and conservatively changed amino acids are indicated by asterisks and dots, respectively, under the sequences.
Similarity of the nucleotide and the deduced amino acid sequences of marmoset P450 2A6 with other P450 2A isoforms

Phylogenetic analysis of marmoset P450 2A6. A phylogenetic tree was created by the neighbor-joining method using P450 2A amino acid sequences of marmoset (cj), human (h), cynomolgus monkey (mf), rhesus monkey (mm), dog (d), rabbit (rab), rat (r), and mouse (m). Human P450 2C9 was used as an outgroup. The scale bar indicates the evolutionary distance corresponding to 10% amino acid differences.
Analysis of Marmoset P450 2A6 Gene Structure.
The exon-intron structure of marmoset P450 2A6 gene was determined by analysis of the marmoset genome data using BLAT; however, the gene sequence of exon 4 was lacking in the genome data, and thus was determined by PCR and sequencing. The marmoset P450 2A6 gene spanned >7.9 kb and consisted of nine exons, similar to human and cynomolgus monkey P450 2A genes. The sizes of the exons and introns ranged from 142 to 188 bp and 239 to 1808 bp, respectively (Table 2). The exact sizes of exon 1 and exon 9 are currently unknown because the sequences of the full-length marmoset P450 2A6 cDNA, including the 5′ and 3′ untranslated regions, were not determined in this study.
Exon-intron boundary sequences of the marmoset P450 2A6 gene
By aligning the marmoset P450 2A6 cDNA sequence against the marmoset genome, the intron-exon boundaries were determined. Exon and intron sequences are shown in small and capital letters, respectively.
Tissue Distribution of P450 2A6 mRNA.
To determine the expression pattern of P450 2A6 mRNA in marmoset tissues, real-time RT-PCR was performed with gene-specific primers in the brain, kidney, liver, lung, and small intestine. Marmoset P450 2A6 mRNA was expressed notably most abundantly in the liver, followed by lung and brain (Fig. 3). These results indicated the predominant expression of marmoset P450 2A6, similar to human P450 2A6.
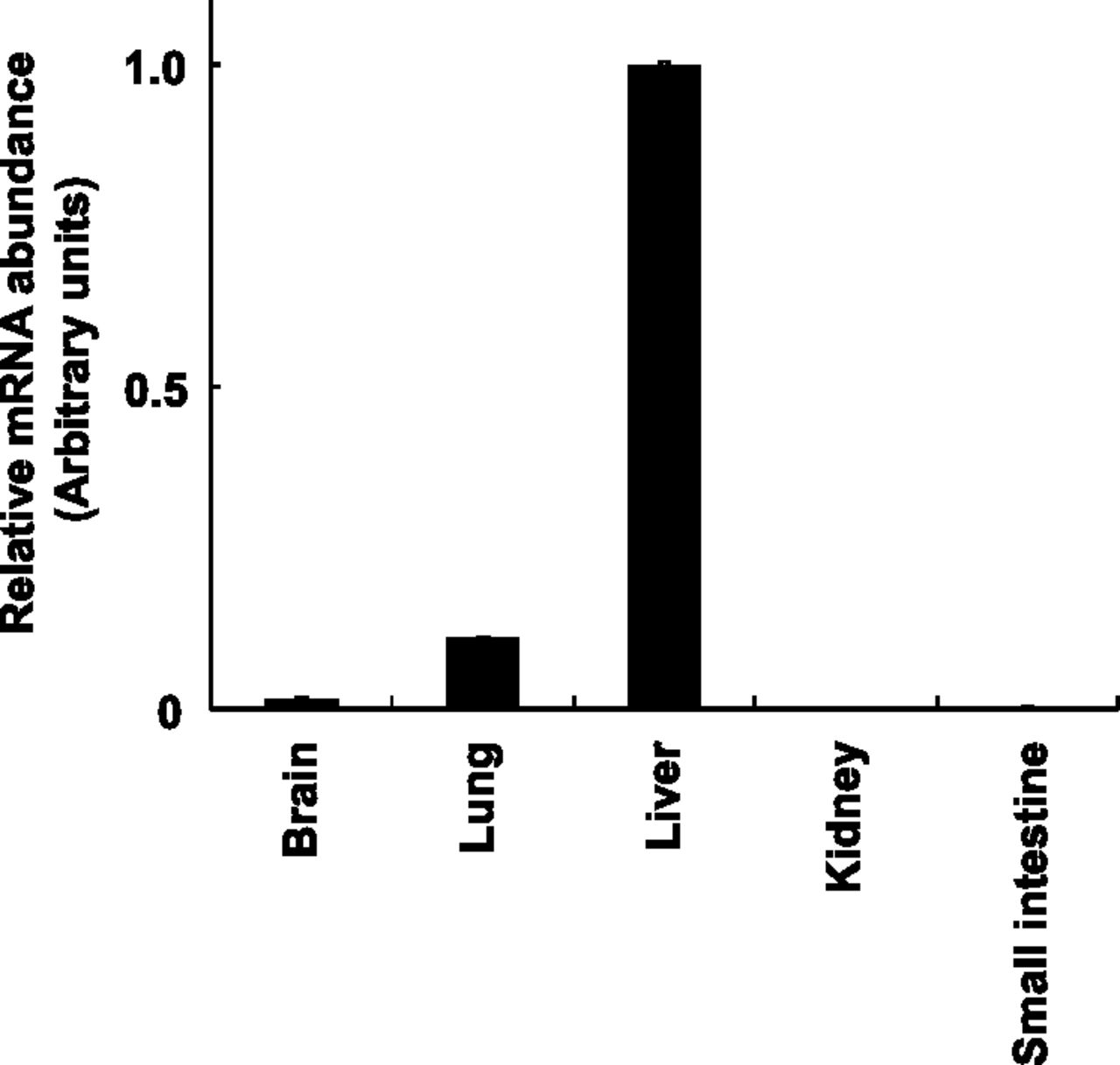
Tissue distribution of P450 2A6 mRNA in marmoset. Expression levels of marmoset P450 2A6 mRNA were measured in brains, lungs, livers, kidneys, and small intestines by real-time RT-PCR and were normalized to 18S rRNA expression levels. All values represent the mean ± S.D. of triplicate determinations. The expression level of marmoset P450 2A6 mRNA in the liver (2.2 × 104 P450 2A6 mRNA/18S RNA) was adjusted to 1, with which all other values were compared.
Expression of Marmoset P450 2A6 Protein in Liver.
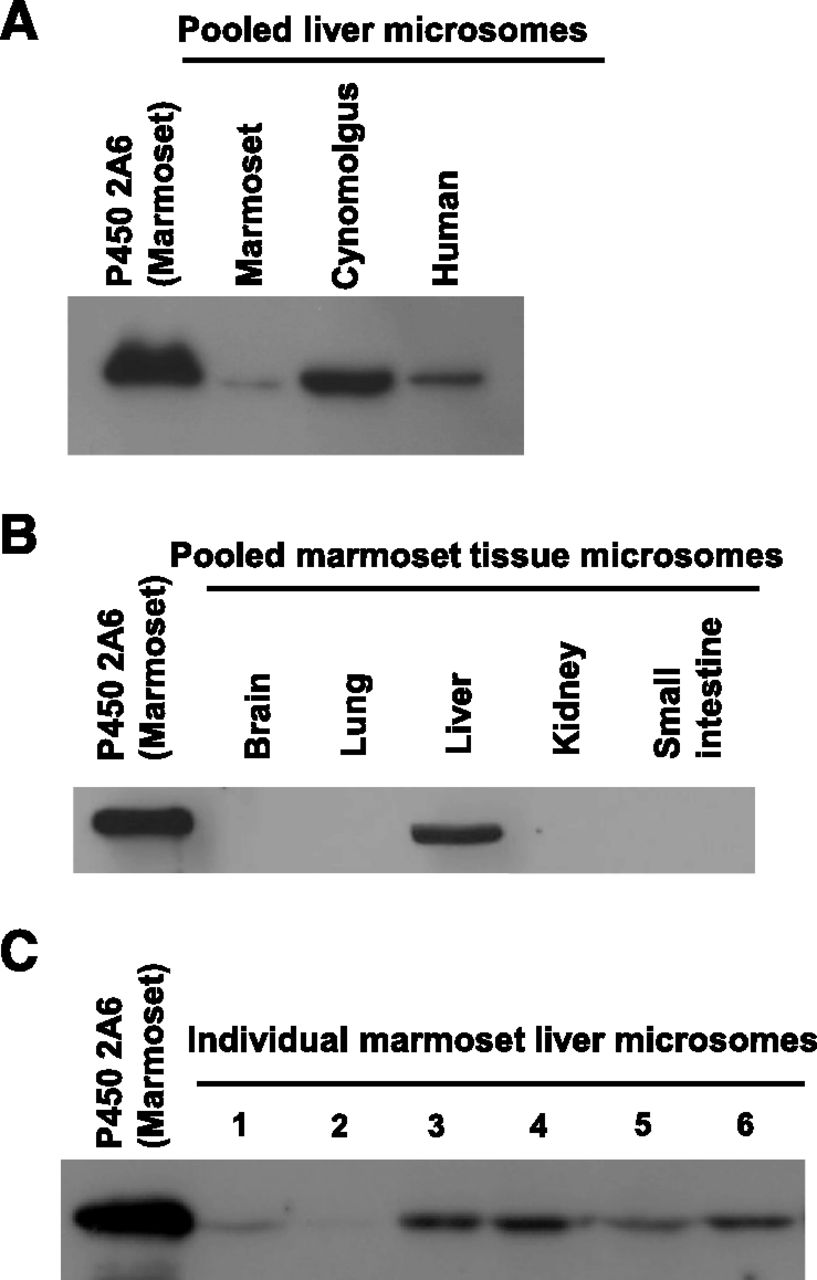
To examine the expression of P450 2A6 protein in marmoset livers, immunoblotting was performed using anti-human P450 2A6 antibodies. The recombinant marmoset P450 2A6 protein was detected by anti-human P450 2A6 antibodies (Fig. 4A), indicating that the antibody was able to react with marmoset P450 2A6 protein. Similarly, a single band (approximately 50 kDa) was detected with marmoset, cynomolgus monkey, and human liver microsomes (Fig. 4A), indicating that P450 2A protein was expressed in the liver from marmosets, similar to cynomolgus monkeys and humans. Marmoset P450 2A6 protein was predominantly detected in liver microsomes among microsomal fractions from five marmoset tissues, including brains, lungs, livers, kidneys, and small intestines tested in this study (Fig. 4B). Moreover, expression of P450 2A protein varied between the six marmoset livers (Fig. 4C), suggesting a large interanimal variation of hepatic P450 2A6 content in marmosets.

Immunoblot analysis of P450 2A6 protein in marmoset tissues. Recombinant marmoset P450 2A6 (0.1 pmol P450/lane) and liver microsomes (15 μg/lane) of marmosets, cynomolgus monkeys, and humans, and tissue microsomal fractions prepared from marmoset brains, lungs, livers, kidneys, and small intestines (15–100 μg microsomal protein/lane) were analyzed by immunoblotting analysis using anti-human P450 2A6 antibodies. Recombinant marmoset P450 2A6 protein was reacted using anti-human P450 2A6 antibodies. The bands of P450 2A proteins were detected in pooled liver microsomes of marmosets, cynomolgus monkeys, and humans (A); marmoset tissue microsomal fractions (100 μg microsomal protein/lane) (B); and individual male (lanes 1–3) and female (lanes 4–6) liver microsomes (15 μg microsomal protein/lane) of marmosets (C).
Activity Characterization and Kinetic Analysis.
To investigate the metabolic activity of marmoset P450 2A6, enzyme assays were carried out using human P450 2A probe substrates, coumarin, 7-ethoxycoumarin, and phenacetin. The kinetics analysis for drug oxidation activities by marmoset P450 2A6, cynomolgus monkey P450 2A23, 2A24, and 2A26, and human P450 2A6 and 2A13 was carried out by nonlinear regression analysis. For coumarin 7-hydroxylation, marmoset P450 2A6 indicated low affinity (Km = 192 μM) and low capacity (Vmax = 0.053 minute−1), showing a markedly lower intrinsic clearance (0.28 μl/min/nmol) than those of human and cynomolgus monkey P450 2As (Table 3). Marmoset liver microsomes showed an apparent Km of 253 μM toward coumarin 7-hydroxylation, similar to marmoset P450 2A6. Kinetic analysis for 7-ethoxycoumarin O-deethylation indicated that the apparent Km and Vmax of marmoset P450 2A6 were 13 μM and 3.9 minutes−1, whereas the intrinsic clearance was 300 μl/min/nmol, suggesting marmoset P450 2A6 effectively catalyzed 7-ethoxycoumarin O-deethylation, approximately similar to human and cynomolgus monkey P450 2As. Moreover, marmoset liver microsomes showed an apparent Km of 16 μM toward 7-ethoxycoumarin O-deethylation, similar to marmoset P450 2A6. In contrast, marmoset P450 2A6 catalyzed phenacetin O-deethylation, unlike human P450 2A6 and cynomolgus monkey P450 2A24 and 2A26. The apparent Km (102 μM) of marmoset P450 2A6 toward phenacetin O-deethylation was higher than that of human P450 2A13 (33 μM) and cynomolgus monkey P450 2A23 (72 μM); although the intrinsic clearance (92 μl/min/nmol) was higher than that of cynomolgus monkey P450 2A23 (57 μl/min/nmol), it was lower than that of human P450 2A13 (291 μl/min/nmol). Marmoset P450 2A6 showed low testosterone hydroxylation activities in the multiple sites and catalyzed negligible rates of testosterone 7α-hydroxylation (Table 4). These results indicated that marmoset P450 2A6 catalyzed human P450 2A substrates with some differences in the substrate specificity, unlike the more correlated human and cynomolgus monkey P450 2As.
Kinetic parameters for oxidation activities of coumarin, 7-ethoxycoumarin, and phenacetin by marmoset P450 2A6 and liver microsomes (LM) and corresponding enzymes in cynomolgus monkeys (C) and humans (H)
Kinetic parameters were calculated by nonlinear regression analysis (mean ± S.E.).
Testosterone hydroxylation activities of recombinant marmoset P450 2A6
Activity was measured at a substrate concentration of 200 μM. Each value represents the mean of duplicate determinations.
Discussion
Common marmosets are potentially useful for preclinical studies in drug development. As we previously reported, similar to human livers, P450 1A-, 2A-, 2B-, 2C-, 2D-, 2E-, 2J-, and 3A-like genes are expressed in marmoset livers (Shimizu et al., 2014); however, these major players in drug metabolism have not been fully identified and characterized. In particular, P450 2A enzymes important for the metabolism of drugs and xenobiotics in humans, marmoset P450 2A enzyme, have not been identified to date. In this study, a novel P450 2A6 cDNA was isolated from marmoset livers based on the marmoset genome data. Phylogenetic analysis of P450 2A amino acid sequences indicated that marmoset P450 2A6 was closely clustered with human, cynomolgus monkey, and rhesus monkey P450 2As (Fig. 2). Marmoset P450 2A6 amino acids showed the highest sequence identity (93%) to human P450 2A13 among all the P450 2As analyzed.
Human lineage diverged from New World monkey lineages approximately 33 million years ago, and the P450 2ABFGST gene cluster diverged through gene duplications and inversions in each species during its evolution. The genome organization of P450 2ABFGST gene clusters does not correspond even between rats, macaques, and humans (Uno et al., 2011). Therefore, determining orthologous relationships between species is a difficult problem. Even marmoset P450 2A6 showed the highest sequence identity (93%) at amino acid level with human P450 2A13 and was phylogenetically much closer to human P450 2A13 than cynomolgus and rhesus monkey P450 2A23, indicating that marmoset was evolutionally closer with human than other nonprimate species. By sequence analysis of the primate P450 2A gene cluster, macaque P450 2A23 gene is hypothesized to be an apparent ortholog of the human P450 2A13 (Hoffman and Hu, 2007). Therefore, marmoset P450 2A6, macaque P450 2A23, and human P450 2A13 genes might have been generated as human P450 2A13-like gene of a primate ancestor during evolution. Further analysis of genomic sequences in the P450 2ABFGST gene cluster is needed to elucidate the orthologous relationship of primate P450 2A genes.
Marmoset P450 2A6 mRNA showed an abundant expression in the liver, but much lower expression in the lung (Fig. 3). In humans, P450 2A6 mRNA is predominantly expressed in the liver, whereas P450 2A13 mRNA is mainly expressed in the lung and respiratory tract, such as nasal mucosa (Ding et al., 1995; Koskela et al., 1999; Su et al., 2000). Human P450 2A6 is a major coumarin 7-hydroxylase in the liver (Yamazaki, et al., 1994). Human P450 2A13 is involved in metabolic activation of 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone in the lung and nasal mucosa (Wong et al., 2005; Zhang et al., 2007) and aflatoxin B1 in respiratory tract (He et al., 2006). Marmoset P450 2A6 might play an important role in metabolism of many toxicants and carcinogens, especially in the extrahepatic tissues (such as lung and nasal mucosa), as well as livers.
Marmoset liver microsomes did not substantially show coumarin 7-hydroxylation activity, unlike human and cynomolgus monkey liver microsomes (Table 3). The apparent Km for marmoset liver microsomes was about 50- and 120-fold higher than those of human and cynomolgus monkey liver microsomes, respectively, and the estimated intrinsic clearance was the lowest among the three species (Table 3), indicating marmoset was a poor metabolizer for coumarin 7-hydroxylation under the presented conditions. Additionally, marmoset P450 2A6 showed much higher Km and much lower intrinsic clearance as compared with human and cynomolgus monkey P450 2As (Table 3). These results were supported by the fact that marmosets reportedly excrete ≤5% the administered dose as urinary 7-hydroxycoumarin (Cohen, 1979; Lake et al., 1989), but humans excrete about 79% (Shilling et al., 1969). In contrast, marmoset liver microsomes showed lower Km and higher intrinsic clearance for phenacetin O-deethylation as compared with human and cynomolgus monkey liver microsomes (Table 3). Marmoset P450 2A6 also showed phenacetin O-deethylation activity, unlike human P450 2A6 (Table 3). Moreover, marmoset P450 2A6 does not have testosterone 7α-hydroxylation activity, unlike rat P450 2A enzyme (Table 4) (Yamazaki et al., 1994). The species differences in P450 2A-dependent drug metabolism need to be considered in drug development. Further investigation of marmoset P450 2A6 on metabolic activity using a variety of P450 substrates would help to better understand drug metabolism in marmosets and species differences from humans.
Although marmoset P450 2A6 and human P450 2A6 retained more than 86% amino acid sequence identity (Table 1), both enzymes showed substantial catalytic differences toward common P450 2A substrates, coumarin and phenacetin (Table 3). Especially, marmoset P450 2A6 showed marginal coumarin 7-hydroxylation, unlike human P450 2A6 (Table 3). Human P450 2A6 efficiently catalyzes coumarin 7-hydroxylation compared with P450 2A13, which differs by only 32 amino acids (von Weymarn and Murphy, 2003). Human P450 2A6 Val117 and Arg372 have been suggested to confer this difference (He et al., 2004). Marmoset P450 2A6 has Ala117, unlike human P450 2A6, but has Arg372 like human P450 2A6. These two substitutions might partly account for the impaired catalytic potential of P450 2A6 for coumarin 7-hydroxylation in marmosets compared with humans.
Phenacetin, metabolized by marmoset P450 2A6 (Table 3), was more efficiently metabolized by P450 2A13 than by P450 2A6 in humans (Fukami et al., 2007). Analysis by site-directed mutagenesis indicated that human P450 2A13 Ser208Ile, Ala213Ser, Phe300Ile, Ala301Gly, Met365Val, and Gly369Ser substitution decreased phenacetin affinity, resulting in the differences in phenacetin metabolism between human P450 2A6 and 2A13 (DeVore et al., 2008). In the deduced amino acid sequences, a total of 52 amino acid residues was different between marmoset and human P450 2A6, including Ser208, Ala213, Phe300, Ala301, Ile365, and Gly369 in marmoset P450 2A6 (Fig. 1). Interestingly, phenacetin was more efficiently metabolized by P450 2A23 than P450 2A24 and 2A26 in cynomolgus monkey. Although amino acid sequence identity among cynomolgus monkey P450 2As was high (>94%), P450 2A23 included differences of Ala213, Phe300, Met365, and Gly369 to those of P450 2A24 and 2A26. These residues of marmoset P450 2A6, relatively well-conserved with human P450 2A13 and cynomolgus monkey P450 2A23, might partly account for the catalytic efficiency of marmoset P450 2A6 in comparison with human P450 2A6 in phenacetin O-deethylation.
The genetic polymorphisms of P450s affect interindividual variability of drug metabolism and the risk of diseases (Sim et al., 2013). A variety of alleles in human P450 2A6 gene was found (see http://www.imm.ki.se/CYPalleles/). Defective P450 2A6 alleles such as P450 2A6*2 and *4 in Caucasian and Asian populations, found among those who smoke, reduced the risk of lung cancer (Fujieda et al., 2004; Rotunno et al., 2009). P450 2A6 deletion and duplication alleles were detected by analysis of copy number variants, and its analysis indicated that the duplication allele frequency was different among Asian, Caucasian, and Hispanic populations (Martis et al., 2013). Various primate species possess copy number variants, including human, bonobo, chimp, gorilla, orangutan, gibbon, macaque, baboon, and marmoset (Dumas et al., 2007). Therefore, genetic polymorphisms of marmoset P450 2A6 gene, if any, might account for the interanimal variability in P450 2A-dependent drug metabolism in the future.
In conclusion, marmoset P450 2A6 cDNA with a high sequence identity (91–94%) to human 2A6, 2A7, and 2A13 cDNA was identified in this study. Among the five tissue types analyzed, marmoset P450 2A6 mRNA was most abundantly expressed in the liver, and in the lung to a much lesser extent. Moreover, marmoset P450 2A6 marginally catalyzed coumarin 7-hydroxylation, but substantially catalyzed 7-ethoxycoumarin O-deethylation and phenacetin O-deethylation. These results indicated that marmoset P450 2A6 had functional characteristics differed from those of human and cynomolgus monkey P450 2As, partially in terms of different substrate specificity, indicating its importance as a model in P450 2A-dependent drug metabolism studies in marmosets and of further studies.
Acknowledgments
The authors thank Drs. Makiko Shimizu and Norie Murayama for technical help.
Authorship Contributions
Participated in research design: Uehara, Uno, Yamazaki.
Conducted experiments: Uehara, Uno.
Contributed new reagents or analytic tools: Inoue, Sasaki.
Performed data analysis: Uehara, Uno, Yamazaki.
Wrote or contributed to the writing of the manuscript: Uehara, Uno, Yamazaki.
Footnotes
- Received February 18, 2015.
- Accepted April 9, 2015.
S.U. and Y.U. equally contributed to this work.
This work was supported in part by Grant-in-Aid for Scientific Research and also resulted from “Construction of System for Spread of Primate Model Animals” under the Strategic Research Program for Brain Sciences of the Ministry of Education, Culture, Sports, Science, and Technology of Japan.
Abbreviations
- PCR
- polymerase chain reaction
- RT
- reverse transcription
- Copyright © 2015 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}