


Abstract
N-Alkylperfluorooctanesulfonamides have been used in a range of industrial and commercial applications. Perfluorooctanesulfonamide (FOSA) is a major metabolite of N-alkylperfluorooctanesulfonamides and has a long half-life in animals and in the environment and is biotransformed to FOSA N-glucuronide. The objective of this study was to identify and characterize the human and experimental animal liver UDP-glucuronosyltransferases (UGTs) that catalyze the N-glucuronidation of FOSA. The results showed that pooled human liver and rat liver microsomes had high N-glucuronidation activities. Expressed rat UGT1.1, UGT2B1, and UGT2B12 in HK293 cells catalyzed the N-glucuronidation of FOSA but at rates that were lower than those observed in rat liver microsomes. Of the 10 expressed human UGTs (1A1, 1A3, 1A4, 1A6, 1A9, 2B4, 2B7, 2B15, and 2B17) studied, only hUGT2B4 and hUGT2B7 catalyzed the N-glucuronidation of FOSA. The kinetics of N-glucuronidation of FOSA by rat liver microsomes and by hUGT2B4/7 was consistent with a single-enzyme Michaelis-Menten model, whereas human liver microsomes showed sigmoidal kinetics. These data show that rat liver UGT1.1, UGT2B1, and UGT2B12 catalyze the N-glucuronidation of FOSA, albeit at low rates, and that hUGT2B4 and hUGT2B7 catalyze the N-glucuronidation of FOSA.
N-Alkylperfluorooctanesulfonamides have been in commercial production since the 1950s (Banks et al., 1994). Because of their chemical stability compared with other chlorinated and brominated organochemicals, N-alkylperfluorooctanesulfonamides have been used in numerous industrial and commercial applications, including the manufacture of surfactants, lubricants, waxes, gloss-finish enhancers, adhesives, anticorrosion agents, stain repellents, fire-fighting foams, food-wrapper coatings, and insecticides (Vander Meer et al., 1986; MacKay, 1991). Perfluorooctanesulfonamide (FOSA) (Fig. 1) is a major metabolite of N-alkylperfluorooctanesulfonamides and has a long half-life in animals and in the environment (Manning et al., 1991; Grossman et al., 1992). FOSA is biotransformed to perfluorooctanesulfonic acid and FOSA N-glucuronide (Fig. 1) (Xu et al., 2004).
The UDP-glucuronosyltransferases (UGTs) catalyze the glucuronidation of carboxy, hydroxyl, amino, acidic carbon, or thiol groups of a variety of substrates to form hydrophilic glucuronides, which facilitate the excretion and detoxification of xenobiotics. UGTs constitute an enzyme superfamily, and the individual isoforms exhibit wide, but overlapping, substrate selectivities (Radominska-Pandya et al., 1999). More than 50 vertebrate UGTs have been identified (Tukey and Strassburg, 2000).
The N-glucuronidation of primary, secondary, and tertiary amines has been extensively studied (Green and Tephly, 1998), and the N-glucuronidation of amides, such as MaxiPost (D. Zhang et al., 2004), carbamazepine (Staines et al., 2004), and dulcin (Uesawa et al., 2004), has recently been reported. Although N-glucuronides of sulfonamides were found as human urinary metabolites when antibacterial agents, such as sulfadimethoxine and sulfamethomidine (Chiu and Huskey, 1998), and a specific cyclooxygenase-2 inhibitor valdecoxib (Yuan et al., 2002) were given, characterization of UGTs responsible for N-glucuronidation of sulfonamides has not been reported. Hence, the previous identification of the N-glucuronide of FOSA (Fig. 1) by rat liver microsomes (Xu et al., 2004) focused our attention on the identification of the UGTs that catalyze the N-glucuronidation of FOSA.
The objective of this study was to identify and characterize the rat and human liver UGTs that catalyzed N-glucuronidation of FOSA. The glucuronidation of FOSA was studied in rat, dog, monkey, and human liver microsomes, with human and rat cDNA-expressed UGTs, and by enzyme kinetic analyses.
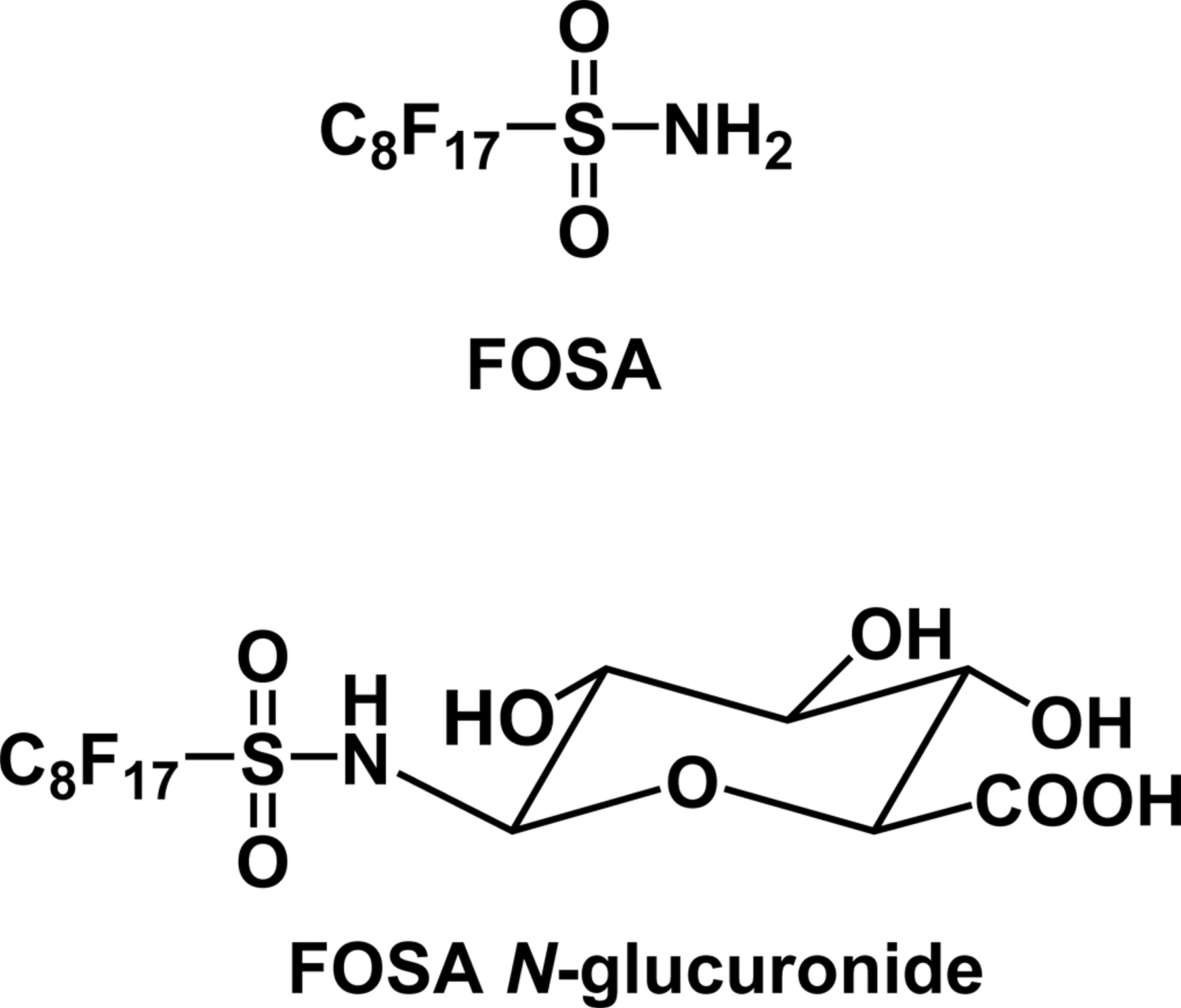
Chemical structures of FOSA and FOSA N-glucuronide.
Materials and Methods
Chemicals and Enzymes. FOSA and 3,3,4,4,5,5,6,6,7,7,8,8,8-tridecafluorooctanesulfonic acid (THPFOS) were supplied by the 3M Co. (St. Paul, MN). FOSA N-glucuronide was synthesized as described previously (Xu et al., 2004). UDP-glucuronic acid (UDPGA), saccharolactone, and alamethicin were purchased from Sigma-Aldrich, Inc. (St. Louis, MO). All the other chemicals were of high-performance liquid chromatography (HPLC) grade and were obtained from VWR International (Bristol, CT) unless specified otherwise.
BD Supersomes from baculovirus-infected Sf9 insect cells that express individual human cDNA for UGT1A1, UGT1A3, UGT1A4, UGT1A6, UGT1A9, UGT2B4, UGT2B7, UGT2B15, and UGT2B17 were purchased from BD Gentest (Woburn, MA). Pooled human liver microsomes (15 males and 10 females, lot 23) were also purchased from BD Gentest. Microsomal fractions from HK293 cells that express each individual rat UGT1.1, UGT2B1, and UGT2B12 were prepared as described previously (Green et al., 1995; King et al., 1997); these UGTs catalyzed the O-glucuronidation of chrysin, testosterone, and borneol, respectively. Rat liver microsomes were prepared as described earlier (Xu et al., 2004). Pooled male beagle dog liver microsomes and pooled male rhesus monkey liver microsomes were purchased from CellzDirect (Austin, TX). (Rat liver UGT1A6, UGT2B2, and UGT2B3 were not available for testing.)
Assay forN-Glucuronidation of FOSA. The complete incubation mixtures contained 0.5 mg/ml microsomal protein (rat, human, dog, monkey liver microsomes, or recombinant UGTs), various concentrations of FOSA (12.5–1000 μM), 5 mM MgCl2, 5 mM saccharolactone, 5 μg of alamethicin, and 5 mM UDPGA in a final volume of 200 μl of 100 mM Tris buffer (pH 7.4). Microsomal protein, Tris buffer, and alamethicin were mixed and placed on ice for 20 min. MgCl2, saccharolactone (aqueous solution), and FOSA (2.5% dimethyl sulfoxide stock solutions) were added, and the mixture was preincubated at 37°C for 5 min. Reactions were initiated by addition of UDPGA, and final organic solvent dimethyl sulfoxide concentration was 0.5% (v/v). Control incubation mixtures lacked UDPGA. The reaction mixtures were incubated for 60 min at 37°C. The reactions were quenched by addition of 200 μl of ice-cold acetonitrile that contained THPFOS, and the mixtures were filtered through 3M Empore protein precipitation filter plates. After dilution with 100 μl of water, 10 μl of the diluted filtrate was injected into the liquid chromatography/mass spectrometer (LC/MS) for analysis. The maximum FOSA incubation concentration was 1 mM because of its limited solubility. The incubation conditions for the formation of FOSA N-glucuronide in liver microsomes (four species) were optimized with respect to protein concentration and incubation time. The reaction rate was linear with protein concentration and incubation time up to 0.5 mg of protein/ml incubation mixture and 150 min, respectively.
LC/MS Analysis of FOSAN-Glucuronide. The LC/MS conditions were similar to those described previously (Xu et al., 2004). The mobile phase was 2 mM ammonium acetate in methanol (solvent A) and 2 mM ammonium acetate in water (solvent B). An Agilent HP1100 HPLC system (Agilent Technologies, Wilmington, DE) fitted with an analytical C18 column (2 × 150 mm, 2-μm particle size) (Waters, Milford, MA) was used. The column was held at ambient temperature and was eluted at a flow rate of 0.3 ml/min. The samples were eluted from the HPLC column with a stepped gradient: 0 min, 45% A; 10 min, 80% A; 12 min, 80% A; and 13 min, 45% A. The eluate was analyzed with a UV diode-array detector and by electrospray mass spectroscopy. The retention times of FOSA N-glucuronide and the internal standard THPFOS were 11.0 and 11.2 min, respectively. LC/MS analyses were performed with an Agilent LC/MSD ion-trap mass spectrometer (Agilent Technologies) with an electrospray interface operated in the negative-ion mode: dry temperature, 350°C; nebulizer, 40 psi; drying gas, 9 l/min; skim 1, –29.0 V; capillary exit, –30 V; and trap drive, 67.3. FOSA N-glucuronide and the internal standard THPFOS were detected in selected daughter-ion modes from MS/MS at unit resolution (FOSA N-glucuronide, m/z 674 → 483; THPFOS, m/z 427 → 407). The standard and quality control samples were prepared by spiking the synthetic FOSA N-glucuronide (0.003–10 μM in final concentration) into a series of incubation mixtures described above (without UDPGA). Standards and QC were processed in the same procedure as the incubation samples described in the N-glucuronidation assay. FOSA N-glucuronide was quantified by comparing peak area ratio of metabolite and THPFOS in incubation samples to a standard curve. The standard curve had a linearity response from 0.01 to 10 μM with correlation coefficients (r2) > 0.99. The lower limit of quantification of FOSA N-glucuronide was 0.01 μM.
Data Analysis. Data were obtained at least in triplicate. The Vmax and Km were derived from nonlinear regression analysis of the experimental data according to the Lineweaver-Burk equation for the typical Michaelis-Menten hyperbolic kinetics (GraphPad Prism 4, GraphPad Software, San Diego, CA) and Hill equation and Eadie-Hofstee plots for the sigmoidal kinetics. The enzyme kinetic model was chosen based on the diagnostic Eadie-Hofstee plots of the data. Goodness of fit to the model was determined by comparison of the sum of the squares of the residuals, the S.E. of the parameters, and coefficient of determination (R2). Kinetic data were reported as mean ± S.D. Intrinsic clearance (CLint) was calculated as Vmax/Km for typical Michaelis-Menten kinetics. For sigmoidal, cooperative kinetics, maximum clearance (CLmax) was calculated as Vmax · (n – 1)/(S50 · n (n – 1)1/n) to estimate the highest clearance (Houston and Kenworthy, 2000; Kaji and Kume, 2005).
Results
Kinetics of theN-Glucuronidation of FOSA in Liver Microsomes. Kinetic analysis of N-glucuronidation in liver microsomes was conducted in four different species: human, rat, dog, and monkey. The kinetics of the N-glucuronidation by rat liver microsomes was consistent with a single-enzyme Michaelis-Menten model. The apparent Km, Vmax, and Vmax/Km were 122 ± 9 μM, 355 ± 9 pmol/min/mg protein, and 2.91 μl/min/mg protein, respectively (mean ± S.D., n = 3) with substrate concentrations of 12.5 to 1000 μM (Fig. 2A). The N-glucuronidation of FOSA by human, dog, and monkey liver microsomes showed sigmoidal kinetics, as shown in a curvilinear Eadie-Hofstee plot (Fig. 2, B–D). The Hill equation was used to obtain the kinetic parameters listed in Table 1 with substrate concentrations of 25 to 1000 μM. With pooled human liver microsomes, the S50 (apparent Km), Vmax, and estimated CLmax were 143 ± 6 μM, 732 ± 18 pmol/min/mg protein, and 2.75 μl/min/mg protein, respectively (mean ± S.D., n = 3). With pooled dog liver microsomes, the S50, Vmax, and estimated CLmax were 195 ± 17 μM, 193 ± 12 pmol/min/mg protein, and 0.50 μl/min/mg protein, respectively. In pooled monkey liver microsomes, the corresponding S50, Vmax, and CLmax were 189 ± 13 μM, 255 ± 14 pmol/min/mg protein, and 0.70 μl/min/mg protein, respectively. The Hill coefficients of kinetic models for human, dog, and monkey were 3.19 ± 0.42, 2.46 ± 0.31, and 2.73 ± 0.32, respectively.
Kinetic parameters of FOSA N-glucuronide formation in liver microsomes from humans, rats, dogs, and monkeys and in recombinant hUGT2B4 and hUGT2B7
The kinetic parameters were derived from nonlinear regression analysis of the experimental data by fitting to a single-enzyme model or Hill equation. Each value represents best-fit values ± S.D. of triplicate points. Kinetic plots are presented in Fig. 2.
N-Glucuronidation of FOSA by Recombinant UGTs. Because most UGT isoforms exhibit distinct, but overlapping, substrate selectivities (Radominska-Pandya et al., 1999; Tukey and Strassburg, 2000) and because many UGT inhibitors are not isoform-selective (Grancharov et al., 2001), recombinant rat and human UGTs were used to identify the UGTs that catalyze the N-glucuronidation of FOSA. The commercially available recombinant human UGT isoforms expressed in baculovirus-infected insect cells were chosen to investigate the N-glucuronidation of FOSA. Of the 10 expressed human UGTs studied, only hUGT2B4 and hUGT2B7 catalyzed the N-glucuronidation of 500 μM FOSA at rates of 32.1 and 882 pmol/min/mg protein, respectively (Fig. 3A). Kinetic parameters for the N-glucuronidation by hUGT2B4 and hUGT2B7 were measured. As shown in Fig. 4, N-glucuronidation by hUGT2B4 and hUGT2B7 was fitted to the single-enzyme Michaelis-Menten kinetics. With hUGT2B4 and substrate concentrations of 100 to 1000 μM in hUGT2B4, the apparent Km, Vmax, and Vmax/Km were 847 ± 122 μM, 88.8 ± 7.6 pmol/min/mg protein, and 0.108 μl/min/mg protein, respectively (mean ± S.D., n = 3; Fig. 4A). With hUGT2B7, the apparent Km, Vmax, and Vmax/Km were 361 ± 32 μM, 1546 ± 71 pmol/min/mg protein, and 4.33 μl/min/mg protein, respectively (mean ± S.D., n = 3; Fig. 4B), with substrate concentrations of 25 to 750 μM.

Kinetics of FOSA N-glucuronide formation catalyzed by rat liver microsomes (A) with 12.5 to 1000 μM FOSA and by human (B), dog (C), and monkey (D) with 25 to 1000 μM FOSA. The inset in A is the Lineweaver-Burk plot of FOSA N-glucuronide formation used for the kinetic parameter measurement. Other insets in B through D show Eadie-Hofstee plots. Data are presented as mean ± S.D., n = 3 except Eadie-Hofstee plots, in which each datum was shown individually.
hUGT2B7 and hUGT2B4 were identified as the UGTs responsible for the N-glucuronidation of FOSA. Rat UGT2B1 shows high gene sequence similarity with human UGT2B7 (Tukey and Strassburg, 2000). Hence, the N-glucuronidation of FOSA was studied with rat UGT2B1 and subsequently with expressed rat UGT1.1 and UGT2B12. With 500 μM FOSA, expressed rat UGT1.1, UGT2B1, and UGT2B12 catalyzed the N-glucuronidation of FOSA at rates of 0.79 ± 0.23, 1.11 ± 0.43, and 0.82 ± 0.20 pmol/min/mg protein, respectively (Fig. 3B).
Discussion
In rats given 50 mg/kg N-ethylperfluorooctanesulfonamide p.o., the metabolically formed FOSA had a long elimination half-life (5 days) in blood and was present at a concentration of 25 μM for 2 days in the elimination phase (Manning et al., 1991; Grossman et al., 1992). N-Glucuronidation of FOSA could be a major metabolic pathway to eliminate FOSA, although the hydrolysis of FOSA, which occurs at a slow rate, may also contribute to its elimination. The metabolically formed FOSA N-glucuronide is excreted in bile and may be hydrolyzed to FOSA by intestinal β-glucuronidase. Therefore, continued enterohepatic circulation may contribute to the long half-life of FOSA.
Although sulfonamides are generally considered to be metabolically stable (Clapp, 1956; Becker et al., 1982), FOSA is biotransformed to perfluorooctanesulfonic acid and FOSA N-glucuronide (Xu et al., 2004). The objective of the present work was to study the N-glucuronidation of FOSA in pooled rat, dog, monkey, and human liver microsomes and with expressed rat and human liver UGTs. The kinetics of the N-glucuronidation of FOSA by rat liver microsomes was consistent with a single-enzyme model, indicating that a single rat UGT isoform was mainly responsible for catalyzing FOSA N-glucuronidation. Expressed rat UGT1.1, UGT2B1, and UGT2B12 catalyzed the N-glucuronidation of FOSA but at rates that were lower than those observed in rat liver microsomes. Because the specific activities of rat UGT1.1, UGT2B1, and UGT2B12 in rat liver microsomes have not been determined, the rates observed with expressed rat UGTs may be attributed to a dilution effect or, alternatively, another UGT isoform may contribute to FOSA N-glucuronidation in rat liver microsomes. Rat UGT2B1 has the highest gene sequence similarity with human UGT2B7 (Tukey and Strassburg, 2000), but the rate of N-glucuronidation of FOSA by UGT2B1 was lower than that observed with hUGT2B7.

N-Glucuronidation of FOSA (500 μM) catalyzed by expressed human (A) and rat (B) liver UGTs. Data are shown as mean ± S.D., n ≥ 3; N.D., not detected.

Lineweaver-Burk plots of FOSA N-glucuronide formation catalyzed by recombinant human UGT2B4 (A) with 100 to 1000 μM FOSA and by human UGT2B7 (B) with 25 to 750 μM FOSA. Data are shown as mean ± S.D., n = 3. The Vmax and Km were derived from nonlinear regression analysis of the experimental data by fitting to a single-enzyme model.
Chiu and Huskey (1998) reported significant differences in rates of the N-glucuronidation of sulfonamide drugs among various species. To explore the species differences in N-glucuronidation of FOSA, FOSA N-glucuronosyltransferase activities in pooled liver microsomes from human, dog, and monkey were investigated. The results showed that human liver microsomes had relatively high N-glucuronidation activities among the species studied (Table 1). The Vmax and intrinsic clearance (CLmax or Vmax/Km) of N-glucuronidation in human liver microsomes were 2- to 4-fold and 1- to 5-fold higher than those found in rat, dog, and monkey, respectively. Human and rat liver UGTs had similar Km values for FOSA, whereas the Km values in dog and monkey liver microsomes were higher. The low clearance in dog and monkey liver microsomes may indicate that N-glucuronidation of FOSA is not a major metabolic pathway in these species.
The observation of sigmoidal kinetics of N-glucuronidation in pooled human, dog, and monkey liver microsomes indicated positive cooperativities, which were identified by the diagnostic Eadie-Hofstee plots (Fig. 2). The atypical kinetic behavior of UGTs has been increasingly reported (Williams et al., 2002; Stone et al., 2003; Kaji and Kume, 2005), although most of the atypical kinetics identified in biotransformation enzymes are associated with CYP3A4 (Tang et al., 1999; Kenworthy et al., 2001; Egnell et al., 2003). For example, estradiol-3-glucuronidation showed homotropic activation in human liver microsomes (Williams et al., 2002). Morphine 3- and 6-glucuronidation by recombinant UGT2B7 expressed in HK293 cells exhibited negative cooperativity (Stone et al., 2003). Atypical enzymatic kinetics was also observed for naproxen glucuronidation in expressed hUGT1A9 (Bowalgaha et al., 2005). Although the mechanism of UGT homotropic effects remains unclear, the following explanations were proposed. As with CYP3A4 cooperativity, the observation of UGT atypical kinetics indicated that allosteric effector sites might exist or multiple substrates could bind to UGT active site simultaneously. Hill coefficients of UGT kinetics obtained from human, dog, and monkey suggested that the minimum number of binding sites on oligomeric UGT was 3, resulting in the substrate autoactivation. Alternatively, homotropic effects may be caused by dimerization of UGT active forms, which may act as cooperative substrate-binding multisubunit enzymes (Miners et al., 2004). The observation of high Km or S50 in those reported and in the present UGT homotropic kinetic study suggested that the observed UGT cooperativity might occur only in vitro systems. No evidence is available about the in vivo autoactivation of UGT. The FOSA concentrations used in the current study were much higher than the blood concentrations of FOSA in experimental animals given FOSA (Manning et al., 1991; Grossman et al., 1992). The concentrations of FOSA in human sera have been reported (Olsen et al., 2003, 2005). In Olsen et al. (2003), all the reported FOSA concentrations were below the limit of quantification (1 ng/ml), whereas in Olsen et al. (2005) only about 2% of the 645 measured values were above the lower limit of quantification (1–3.2 ng/ml).
Both hUGT2B4 and hUGT2B7 catalyzed the N-glucuronidation of FOSA and showed typical hyperbolic kinetics (Fig. 2). hUGT2B7's intrinsic clearance (Vmax/Km) was similar to the values found in pooled human liver microsomes, although both the Km and Vmax of hUGT2B7 were 2-fold higher than those of human liver microsomes. These observations indicated that hUGT2B7 may be the major hUGT that catalyzes the N-glucuronidation of FOSA. The variability in kinetic behavior between human liver microsomes and recombinant hUGT2B7 may attribute to their different enzyme components. Human liver microsomes contained both hUGT2B4 and hUGT2B7, which have different Km and Vmax for FOSA. The superposition of velocity curves for both UGT isoforms in human liver microsomes may show a nonhyperbolic curve that resembles sigmoidal behavior (Palmer, 1985). However, the kinetic curve in human liver microsomes did not fit a two-enzyme Michaelis-Menten equation. Another plausible explanation was that the properties of the native hUGT4/7 were altered during the expression in Sf9 insect cells and that a heterodimer of UGT does not exist in insect cells. This hypothesis needs to be tested in future studies, although a distinction between CYP3A4 expressed in insect cells versus native human liver microsomes was observed (Z. Zhang et al., 2004). hUGT2B4 and hUGT2B7 show 89% similarity in their gene sequences (Tukey and Strassburg, 2000), but the intrinsic clearance of N-glucuronidation of FOSA by hUGT2B7 was about 40-fold higher than that of hUGT2B4. Previous studies indicate that hUGT2B4 has the same substrate selectivity as UGT2B7 but with markedly lower catalytic activity (Jin et al., 1997; Miners et al., 2004). The present results are consistent with these findings.
UGT2B7 catalyzes the glucuronidation of a variety of drugs and chemicals (King et al., 2000), including morphine (Coffman et al., 1997), diclofenac (King et al., 2001), zidovudine (Barbier et al., 2000), and hyodeoxycholic acid (Ritter et al., 1992). It was recently reported that UGT2B7 catalyzes the N-glucuronidation of the amide nitrogen of MaxiPost (D. Zhang et al., 2004) and carbamazepine (Staines et al., 2004). The present study shows that the N-glucuronidation of FOSA provides another novel amide substrate for UGT2B7. UGT2B7 is widely distributed in human tissues (King et al., 1999) and therefore may play a major role in the conjugation and elimination of FOSA in both rats and humans.
Acknowledgments
We thank Gloria Kwei, Andreas Harsch, Sujal Vilas Deshmukh, and John Kevin Leach for helpful suggestions and advice.
Footnotes
-
This research was supported in part by the 3M Company and by Merck & Co., Inc.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.106.009399.
-
ABBREVIATIONS: FOSA, perfluorooctanesulfonamide; UGT, UDP-glucuronosyltransferase; THPFOS, 3,3,4,4,5,5,6,6,7,7,8,8,8-tridecafluorooctanesulfonic acid; UDPGA, UDP-glucuronic acid; HPLC, high-performance liquid chromatography; LC/MS, liquid chromatography/mass spectrometry.
-
↵1 Current affiliation: Amylin Pharmaceuticals Inc., San Diego, California.
- Received February 10, 2006.
- Accepted May 19, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}