


Abstract
In the present study, we attempted to identify the membrane permeation process(es) primarily involved in the molecular-weight-dependent biliary excretion of β-lactam antibiotics. A search of the literature indicated that the molecular weight threshold operates mainly in the transport process across bile canalicular membranes. We confirmed that biliary clearance of the model biliary-excretion-type cephalosporin cefoperazone was reduced to 10% of the control in Eisai hyperbilirubinemic rats, which are genetically deficient in multidrug resistance-associated protein (Mrp) 2, indicating that Mrp2 plays a major role as an efflux transporter on the canalicular membranes. ATP-dependent uptake of several cephalosporins including cefoperazone, cefbuperazone, cefpiramide, and ceftriaxone, all of which are mainly excreted into bile, was confirmed in membrane vesicles from Sf9 cells transfected with rat Mrp2. Both the inhibitory potency of the cephalosporins for Mrp2-mediated transport and the uptake of cephalosporins by Mrp2-expressing vesicles were molecular weight-dependent, suggesting that Mrp2 is one of the major transporters involved in molecular weight-dependent biliary excretion. An uptake study in membrane vesicles of Sf9 cells transfected with breast cancer resistance protein (Bcrp) revealed that Bcrp accepts cefoperazone, cefbuperazone, cefpiramide, cefotetan, ceftriaxone, cefotiam, cefamandole, and cefazolin as substrates, and Bcrp-mediated transport was also molecular weight-dependent, suggesting that Bcrp also contributes to molecular weight-dependent biliary excretion of β-lactam antibiotics in rats.
Biliary excretion is one of the major elimination pathways for metabolites of both endogenous compounds and xenobiotics, including therapeutic agents. Biliary excretion is highly dependent on molecular weight, and a threshold of molecular weight is observed for the biliary excretion of organic anions: compounds with molecular weights higher than 325 ± 50 and 500 ± 50 are preferentially excreted into the bile in rats and humans, respectively (Hirom et al., 1972; Levine, 1978). Biliary excretion results from concentrative transport processes across both sinusoidal and canalicular membranes, and various types of transport systems have been characterized on both membranes. However, the mechanism(s) underlying the observed molecular weight dependence remains to be fully clarified.
At least some of the mechanisms involved in the transport of organic anions across sinusoidal and canalicular membranes have recently been identified at the molecular level. On sinusoidal membranes, members of the organic anion-transporting polypeptide (OATP/SLCO) family are expressed and are thought to be involved in sinusoidal uptake of organic anions (Tamai et al., 2000; Kim, 2003; Chandra and Brouwer, 2004; Hagenbuch and Meier, 2004; Shitara et al., 2005): Among the SLCO family members, OATP-B (SLCO2B1), OATP-C (SLCO1B1), and OATP-8 (SLCO1B3) are sinusoidal membrane transporters in humans, whereas those in rats are Oatp1 (Slco1A1), Oatp2 (Slco1a4), and Oatp4 (Slco1b2) (Hagenbuch and Meier 2004; Shitara et al., 2005). The effects of genetic polymorphisms on drug disposition, as recently reported for various therapeutic agents in humans, strongly suggest a predominant role of OATP-C in hepatic handling of organic anions (Niemi et al., 2005; Ieiri et al., 2007). Sinusoidal efflux transporters have also been identified, and sodium/phosphate transporter (NPT) 1 (Slc17A1), multidrug resistance-associated protein (MRP) 3 (ABCC3), and MRP4 (ABCC4) have been proposed to be involved in the efflux of therapeutic agents and/or their metabolites (Yabuuchi et al., 1998; Uchino et al., 2000; Manautou et al., 2005; Zamek-Gliszczynski et al., 2006a; van de Wetering et al., 2007). On the other hand, several efflux transport systems have been identified in bile canalicular membranes, and a fundamental role of MRP2 (ABCC2) has been suggested, particularly in rats (Chandra and Brouwer, 2004; Nies and Keppler, 2007). Besides MRP2, breast cancer resistance protein (BCRP/ABCG2) and bile salt export pump (BSEP/ABCB11) are also proposed to be involved in biliary excretion of organic anions (Trauner and Boyer, 2003; Chandra and Brouwer, 2004).
To characterize the mechanisms involved in the molecular weight threshold, it is important to use appropriate model compounds. In the present study, we focused on β-lactam antibiotics. Biliary excretion of cephalosporins is highly dependent on molecular weight in rats: less than 15% of the dose is excreted into the bile for cephalosporins with a molecular weight of less than 450, but those with a molecular weight of more than 450 exhibit 15 to 100% recovery in bile (Wright and Line, 1980). In addition, their elimination pathway is mainly excretion into bile and/or urine with minimal metabolism in the body (Tsuji et al., 1983; Tsuji, 2006). Therefore, they may be suitable as model compounds for examining the determinants of the molecular weight threshold. Transport mechanism(s) for β-lactam antibiotics in the liver have already been studied using isolated hepatocytes and membrane vesicles, and the recognition specificity for various β-lactam compounds of the transporters localized on both sinusoidal and canalicular membranes has been clarified to be a major determinant of efficiency of biliary excretion (Terasaki et al., 1986; Tamai et al., 1987, 1988, 1990; Tsuji et al., 1990). In addition, molecular mechanism(s) involved in their permeation across both membranes have been identified, and those include OATP-B, OATP-C, Oatp2, and NPT1 across the sinusoidal membranes (Yabuuchi et al., 1998; Tamai et al., 2000; Uchino et al., 2000; Nakakariya et al., 2008) and Mrp2 across the canalicular membrane (Sathirakul et al., 1993; Muraoka et al., 1995). However, there has been only minimal systematic analysis of the molecular mechanisms involved in biliary excretion of β-lactams. Because biliary excretion involves permeation processes through various membranes, including sinusoidal and canalicular membranes, we have focused here on the β-lactam transport mechanism(s) localized on canalicular membranes. We have sought to identify the membrane permeation process primarily involved in biliary excretion of cephalosporin antibiotics by means of literature search, a biliary excretion study in Eisai hyperbilirubinemic rats (EHBRs), which are genetically deficient in Mrp2, and transport studies using membrane vesicles expressing canalicular transporters.
Materials and Methods
Materials. Cefoperazone sodium salt, ceftriaxone sodium salt, cefmetazole sodium salt, cefamandole sodium salt, cefazolin sodium salt, cephalexin, and ethacrynic acid were purchased from Sigma-Aldrich Japan K.K. (Tokyo, Japan). Bromosulfophthalein (sodium salt), cyclosporin A, and S-(2,4-dinitrophenyl)-glutathione were from Wako Pure Chemical Industries (Osaka, Japan). Cefbuperazone, cefpiramide sodium, cefotetan, and cefotiam hydrochloride were gifts from Toyama Chemical Co., Ltd. (Tokyo, Japan), Dainippon Sumitomo Pharma Co., Ltd. (Osaka, Japan), Astellas Pharma Inc. (Osaka, Japan), and Taisyo Toyama Pharmaceutical Co., Ltd (Tokyo, Japan), respectively. [3H]Estradiol-17β-d-glucuronide (E217βG) (40.5 Ci/mmol, 97%) was purchased from PerkinElmer Life and Analytical Sciences (Boston, MA). Membrane vesicles prepared from Sf9 cells expressing rat Mrp2 (rMrp2), rat Bcrp (rBcrp). or human MRP2 (hMRP2) were supplied by Genomembrane, Inc. (Yokohama, Japan). All other reagents were commercial products of reagent grade.
Animals. Male Sprague-Dawley rats (SDRs) (7–9 weeks old) were purchased from Sankyo Labo Service Corporation, Inc. (Toyama, Japan). EHBRs (7–9 weeks old) were purchased from Sankyo Labo Service Corporation, Inc. or supplied by Eisai Co., Ltd. (Tokyo, Japan). This study was carried out in accordance with the Guide for the Care and Use of Laboratory Animals in Takara-machi Campus of Kanazawa University.
Biliary and Urinary Excretion of Cefoperazone and Cephalexin in SDRs and EHBRs. Under pentobarbital anesthesia (50 mg/kg), the femoral vein was cannulated with polyethylene catheters (SP31; Natsume, Tokyo) for the injection of cefoperazone and cephalexin. The bile duct and bladders were similarly cannulated for the collection of bile and urine, respectively. After the animals had recovered from anesthesia, cefoperazone or cephalexin (20 mg/kg dissolved in saline) was intravenously injected, and blood samples were collected from the jugular vein and immediately centrifuged to obtain plasma. Plasma, bile, and urine samples were deproteinized with 2 volumes of acetonitrile (for cefoperazone) or methanol (for cephalexin), followed by centrifugation. A 20-μl aliquot of the supernatant was subjected to LC-UV analysis using a Tosoh ODS 80 TM column (4.6 × 150 mm; Tosoh Co., Tokyo, Japan). For cefoperazone the mobile phase consisted of acetonitrile-water containing 10 mM ammonium acetate and 10 mM tetra-N-butylammonium bromide (24:76, v/v), the flow rate was 1.5 ml/min, and the detection wavelength was 270 nm. For cephalexin, the mobile phase consisted of methanol-water containing 10 mM ammonium acetate (20:80, v/v), the flow rate was 1.0 ml/min, and the detection wavelength was 260 nm.
Infusion Study for Cefoperazone in SDRs. Under pentobarbital anesthesia, infusion of cefoperazone was performed at 3.22 mg/h via the jugular vein in SDRs. The plasma, liver, and kidney were obtained at 1 h after the start of infusion to determine the tissue/plasma concentration ratio. The tissue sample was homogenized with an equal volume of water, then 4 volumes of acetonitrile was added, and the mixture was centrifuged. A 50-μl aliquot of the supernatant was subjected to LC-UV analysis as described above.
Calculation of Pharmacokinetic Parameters. The area under the curve (AUC) and the area under the first-moment curve (AUMC) were calculated from the plasma concentration-time profile with extrapolation to infinity by application of the trapezoidal rule. Total clearance (CLtotal) was determined as dose/AUC. The mean residence time was calculated as AUMC/AUC. The biliary clearance with regard to plasma concentration (CLbile, plasma) was calculated as Xbile/AUC, where Xbile represents the amount excreted into the bile. Statistical analysis was performed with Student's t test. The criterion of significance was taken to be p < 0.05.
In the analysis of literature information, we calculated CLbile, plasma as well as the biliary clearance with regard to hepatic concentration (CLbile, liver) where possible to focus on membrane permeation across the bile canalicular membrane. If the time profile of hepatic concentration was reported, the CLbile, liver was calculated as 
If the hepatic concentration was available only at a few sampling points, the AUCliver cannot be accurately estimated, and, therefore, the CLbile, liver was approximately estimated by the following equation:  where AUC is area under the curve for plasma concentration, and Kp, liver is the liver/plasma concentration ratio. Theoretically, eq. 2 is correct at steady state but would not be accurate after a bolus administration because the Kp, liver can change time dependently. In the present study, therefore, the Kp, liver was estimated as the mean value of liver/plasma concentration at all the sampling points available in the literature.
where AUC is area under the curve for plasma concentration, and Kp, liver is the liver/plasma concentration ratio. Theoretically, eq. 2 is correct at steady state but would not be accurate after a bolus administration because the Kp, liver can change time dependently. In the present study, therefore, the Kp, liver was estimated as the mean value of liver/plasma concentration at all the sampling points available in the literature.
Urinary secretion was also normalized by renal concentration, and renal secretion clearance with regard to renal concentration (CLurine, kidney) was calculated by applying the following equation, if the time profile of the renal concentration was available:  where fp is plasma unbound fraction, and GFR is glomerular filtration rate, which was assumed to be 5.24 ml/min/kg in rats (Davies and Morris, 1993). As for liver, if the renal concentration was available only at a few sampling points, the CLurine, kidney was approximately estimated as follows:
where fp is plasma unbound fraction, and GFR is glomerular filtration rate, which was assumed to be 5.24 ml/min/kg in rats (Davies and Morris, 1993). As for liver, if the renal concentration was available only at a few sampling points, the CLurine, kidney was approximately estimated as follows:  where Kp, kidney is the kidney/plasma concentration ratio and was estimated as the mean value of kidney/plasma concentration at all the sampling points available in the literature. In the present study, we assumed no tubular reabsorption of cephalosporins.
where Kp, kidney is the kidney/plasma concentration ratio and was estimated as the mean value of kidney/plasma concentration at all the sampling points available in the literature. In the present study, we assumed no tubular reabsorption of cephalosporins.
Transport Studies with Membrane Vesicles Expressing MRP2 and BCRP. The transport studies were performed using a rapid filtration technique according to the manufacturer's protocol with a minor modification. In brief, 40 μl of transport medium (50 mM MOPS-Tris (pH 7.0), 70 mM KCl, 7.5 mM MgCl2, and 2 mM glutathione), containing membrane vesicles (50–100 μg of protein) and test compounds, was preincubated at 37°C for 5 min and then rapidly mixed with the reaction mixture containing 4 mM ATP or AMP with an ATP-regenerating system (10 mM creatine phosphate and 100 μg/μl creatine phosphokinase). The transport reaction was terminated by the addition of 1 ml of ice-cold buffer containing 40 mM MOPS-Tris (pH 7.0) and 70 mM KCl. The stopped reaction mixture was then filtered through a 0.45-μm HAWP filter (Millipore Corporation, Billerica, MA) and washed four times with 5 ml of stop solution.
For determination of cephalosporin antibiotics trapped on the membrane filters, the filters were dried and cut into small pieces. The drug was extracted with 1 ml of water containing internal standard (cephradine) for 20 min and then centrifuged, and 0.8 ml of the sample was loaded onto a Strata-X tube (Phenomenex, Torrance, CA) preconditioned with 1 ml of methanol and 2 ml of water. The drug was eluted from the column with 1 ml of methanol. The eluate was evaporated to dryness at 45°C under a stream of nitrogen, and the residue was redissolved in 40 μl of mobile phase. The concentration of cephalosporin was measured by with a LC-tandem mass spectrometry system equipped with a constant flow pump (1200 series G1312A; Agilent Technologies, Tokyo, Japan), an automatic sample injector (G1367B; Agilent Technologies), a column oven (G1316A; Agilent Technologies), and a tandem mass spectrometer (API 3200; Applied Biosystems, Tokyo, Japan). The analytical column was COSMOSIL 5C18-AR-II (2.0 mm × 150 mm; Nacalai Tesque, Kyoto, Japan). Mobile phase A was 0.01 M ammonium formate and mobile phase B was 100% methanol. The gradient elution time program was set as follows: 0 to 4 min, B5to 35%; 4 to 15 min, B, 35%; 15.0 to 15.1 min, B, 35 to 5%; and 15.1 to 25 min, B, 5%. Mass spectrometric detection was done from 9 to 15 min in the positive ion mode based on the transition of mass/charge ratio (m/z): 646 to 143, 628 to 143, 613 to 257, 555 to 125, 526 to 174, 472 to 215, 463 to 158, 455 to 323, 350 to 176, and 348 to 158 for cefoperazone, cefbuperazone, cefpiramide, ceftriaxone, cefotiam, cefmetazole, cefamandole, cefazolin, cephradine, and cephalexin, respectively. Cefotetan was detected in the negative ion mode at m/z 574. The flow rate was 0.2 ml/min. The recovery of all the cephalosporins examined from the membrane filters was more than 80%. For the determination of inhibitory effects of various compounds on [3H]E217βG uptake, membrane vesicles were preincubated with [3H]E217βG in the presence or absence of an inhibitor at 37°C for 5 min, followed by the start of reaction by rapid mixture with a solution containing ATP or AMP with an ATP-regenerating system. Radioactivity of [3H]E217βG associated with the filter was determined in a liquid scintillation counter LSC-5100 (Aloka, Tokyo, Japan) with Clearsol I (Nacalai Tesque Inc.) as the scintillation fluid.
Uptake of a test compound by the membrane vesicles was normalized by the medium concentration to give the distribution volume with dimensions of microliters per milligram of protein. The MRP2-mediated uptake was obtained by subtraction of the uptake by control vesicles from that by MRP2-expressing vesicles and fitted to the Michaelis-Menten equation.
Concentration-dependent inhibition of Mrp2-dependent uptake of [3H]E217βGby β-lactam antibiotics was fitted to either of the following equations: 
 where R is the uptake normalized by the control (without inhibitor) value, I is the inhibitor concentration, IC50 is the inhibition constant, and r is the contribution ratio of the higher affinity component to the overall uptake of [3H]E217βG. These fittings were performed by an iterative nonlinear least-squares method using the MULTI program (Yamaoka et al., 1981), and the algorithm used for the fitting was the damping Gauss-Newton method.
where R is the uptake normalized by the control (without inhibitor) value, I is the inhibitor concentration, IC50 is the inhibition constant, and r is the contribution ratio of the higher affinity component to the overall uptake of [3H]E217βG. These fittings were performed by an iterative nonlinear least-squares method using the MULTI program (Yamaoka et al., 1981), and the algorithm used for the fitting was the damping Gauss-Newton method.
Results
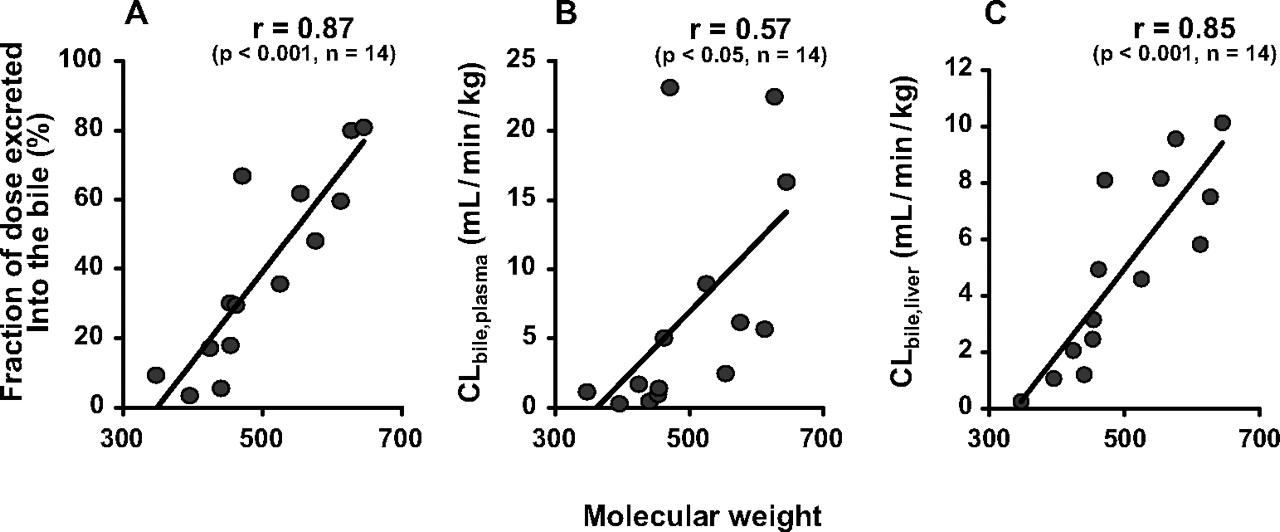
Literature Search for Information on Biliary Excretion of β-Lactam Antibiotics. To analyze the molecular weight dependence in biliary excretion of β-lactam antibiotics, we first searched the literature on biliary excretion of the antibiotics in rats (Table 1). We focused in the present study on cephalosporins for the following reasons. First, they are minimally metabolized in the body, so the influence of metabolism on distribution can be neglected. Second, they are anionic at neutral pH, which is important because a molecular weight threshold is known to exist for organic anions (Hirom et al., 1972; Levine, 1978). Cephalexin also has a cationic moiety but was selected as a urinary excretion-type antibiotic. A correlation between the molecular weight and fraction of dose recovered in the bile has been reported (Hirom et al., 1972; Levine, 1978). Therefore, information on the fraction of dose recovered in the bile was collected from the literature (Table 1) and plotted against molecular weight for cephalosporins. The results confirmed the molecular weight dependence of biliary excretion of these compounds, because the fraction of dose recovered in the bile was well correlated with molecular weight (Fig. 1A). According to pharmacokinetic theory, the fraction of dose would be influenced by excretion into both bile and urine. Because we wished to focus on biliary excretion to identify the membrane permeation process primarily involved in the molecular weight threshold, biliary clearances with regard to plasma and hepatic concentrations (CLbile, plasma and CLbile, liver, respectively) were also collected from the literature. There was a significant correlation with molecular weight for both clearances, but it was higher for CLbile, liver than for CLbile, plasma (Fig. 1, B and C). Thus, molecular weight dependence is observed in the excretion process of cephalosporins across the bile canalicular membranes. On the other hand, no obvious relationship was observed between CLurine, kidney and molecular weight of cephalosporins (Table 1).
Pharmacokinetic parameters of β-lactam antibiotics in rats

Molecular weight dependence of fraction of dose excreted into the bile (A), CLbile, plasma (B), and CLbile, liver (C) for β-lactam antibiotics in rats. Pharmacokinetic parameters for each compound were obtained from the literature, as shown in Table 1.
Mrp2 Is Primarily Involved in Biliary Excretion of Cefoperazone in Rats. To analyze the molecular mechanism(s) underlying permeation across the bile canalicular membrane, biliary excretion of cefoperazone, which has the highest molecular weight and is most extensively excreted into the bile among the cephalosporins (Table 1), was compared in SDRs and EHBRs. Cephalexin, which has the lowest molecular weight and is excreted mainly into the urine (Table 1), was also examined as a control. Plasma disappearance of cefoperazone was delayed in EHBRs compared with SDRs, whereas that of cephalexin was similar in the two strains (Fig. 2, A and B). The CLtotal for cefoperazone was significantly lower and the mean residence time was higher in EHBRs than in SDRs, whereas the same parameters for cephalexin exhibited little difference between the two strains (Table 2). Approximately 80% of cefoperazone administered i.v. was recovered in bile in SDRs, whereas the biliary excretion of cefoperazone in EHBRs was greatly reduced (∼30% of the dose was recovered into the bile), and ∼70% was recovered in urine in EHBRs (Fig. 2, C and E). Biliary excretion of cephalexin was much smaller than that of cefoperazone, but the biliary excretion of cephalexin was also lower in EHBRs than SDRs (9.29 ± 1.1 and 1.47 ± 0.21% of the dose was recovered in the bile over 24 h in SDRs and EHBRs, respectively) (Fig. 1D). The CLbile, plasma in EHBRs was only 10% of that in SDRs for both cefoperazone and cephalexin (Table 2).

Mrp2 is primarily involved in biliary excretion of cefoperazone in rats. Cefoperazone (A, C, and E) or cephalexin (B, D, and F) was injected i.v. (20 mg/kg) in SDRs (○) or EHBRs (•), and the time profile for plasma concentration (A and B), biliary (C and D), and urinary (E and F) excretion was measured. Each point represents the mean ± S.E.M. of three rats. When error bars are not shown, they are smaller than the symbols.
Pharmacokinetic parameters of cefoperazone and cephalexin in SDR and EHBRs
Primary Active Transport of Cephalosporins by Mrp2. We next attempted to examine whether cefoperazone and other cephalosporins, which are substantially excreted into the bile, are transported by rMrp2. For this purpose, membrane vesicles prepared from Sf9 cells transfected with rMrp2 gene were used for an uptake study. Uptake of cephalosporins by the rMrp2-expressing vesicles was first screened as a cassette dosing for 10 compounds, 9 compounds with relatively high molecular weight and cephalexin as a control (Table 1; data not shown). Uptake of cefoperazone, cefpiramide, and ceftriaxone by rMrp2-expressing vesicles increased almost linearly up to 5 min, whereas that by control vesicles and that in the presence of AMP did not (data not shown). Therefore, uptake of each compound was then separately measured for 5 min to compare the absolute value for the uptake among compounds (Fig. 3). Uptake of cefoperazone, cefbuperazone, cefpiramide, and ceftriaxone by rMrp2-expressing vesicles was significantly higher than that by control vesicles (Fig. 3A), suggesting that these compounds are substrates of rMrp2. The rMrp2-mediated uptake, assessed by subtraction of the uptake by control vesicles from that by rMrp2-expressing vesicles, exhibited molecular weight dependence, although the molecular weight alone could not explain the uptake values (Fig. 4A). The uptake of cefoperazone in the presence of ATP was higher than that in the presence of AMP (Fig. 3A) and exhibited saturation (Fig. 5) in both rMrp2-expressing and control vesicles. Kinetic parameters were determined for rMrp2-mediated uptake of cefoperazone in the presence of ATP, with Km and Vmax being 70.9 ± 58.1 μM and 195 ± 116 pmol/mg of protein/min, respectively.
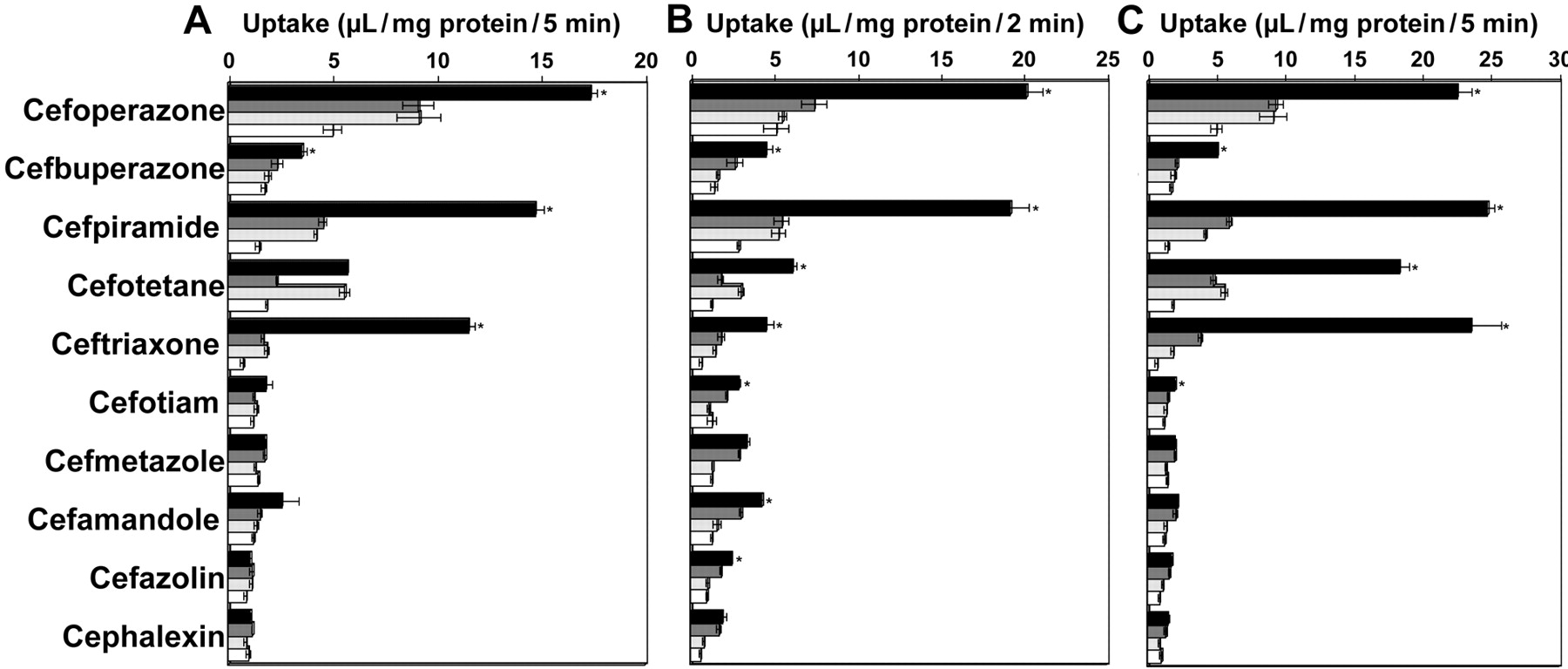
Uptake of β-lactam antibiotics by membrane vesicles expressing rMrp2 (A), rBcrp (B), or hMRP2 (C). The membrane vesicles were separately incubated with 20 μM of each compound dissolved in medium containing 4 mM ATP or AMP at 37°C. Black and gray columns represent the uptake by the vesicles expressing each transporter in the presence of ATP and AMP, respectively, whereas light gray and white columns represent the uptake in control vesicles in the presence of ATP and AMP, respectively. Each datum shows the mean ± S.E.M. (n = 3). When error bars are not shown, they are smaller than the symbols. *, significant difference from the control vesicles.
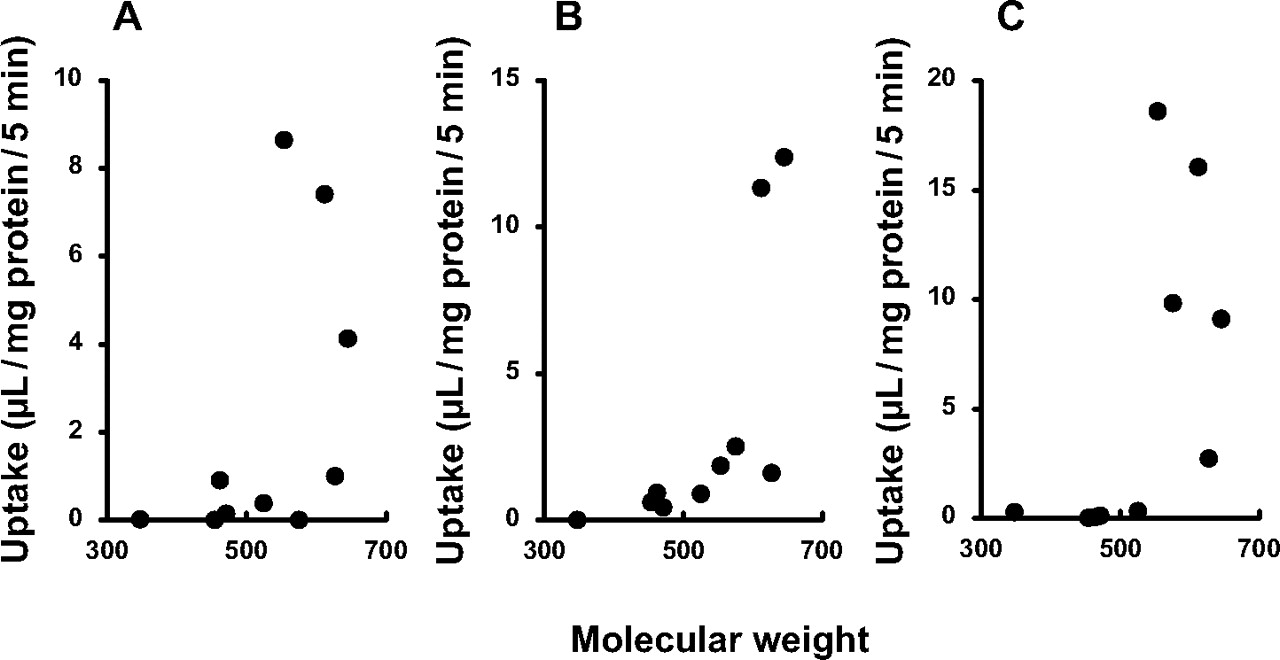
Molecular weight dependence of the uptake of β-lactam antibiotics by membrane vesicles expressing rMrp2 (A), rBcrp (B), or hMRP2 (C). Uptake data shown in Fig. 4 were used to calculate uptake mediated by each transporter by subtracting the uptake in the presence of ATP by control vesicles from that in each transporter-expressing vesicle. The values obtained were plotted against molecular weight.

Kinetic analysis of cefoperazone uptake by rMrp2-expressing membrane vesicles. Uptake of cefoperazone (1–10,000 μM) for 5 min was determined at 37°C in the presence of ATP. Uptake in rMrp2-expressing (•) and control (○) vesicles, respectively. Each point and vertical bar represents the mean ± S.E.M. (n = 3–5). The solid lines are those fitted by using nonlinear least-squares analysis, whereas the broken lines are those fitted to saturable transport by rMrp2, obtained by subtracting the uptake by control vesicles from that by rMrp2-expressing vesicles.
To obtain further evidence of recognition of cephalosporins by rMrp2, inhibition of the uptake of [3H]E217βG by rMrp2-expressing vesicles was examined. The rMrp2-mediated [3H]E217βG uptake was inhibited by cefoperazone, cefpiramide, ceftriaxone, cefotiam, cefmetazole, cefazolin, and cephalexin in a concentration-dependent manner (Fig. 6A). The IC50 values were 199 ± 23 μM, 368 ± 31 μM, 3.26 ± 0.53 mM, 1.41 ± 0.15 mM, 61.6 ± 37.2 mM, and 15.6 ± 6.0 mM for cefpiramide, ceftriaxone, cefotiam, cefmetazole, cefazolin, and cephalexin, respectively. The inhibition profile for cefoperazone was biphasic (Fig. 6A), yielding IC50, high and IC50, low values of 6.66 ± 3.23 μM and 3.88 ± 1.32 mM, respectively.

Inhibition of Mrp2-mediated uptake of E217βGby β-lactam antibiotics. A, uptake of [3H]E217βG (0.5 μM) for 5 min was determined at 37°C in the presence of ATP and various concentrations of cefoperazone (black circle), cefpiramide (black triangle), ceftriaxone (gray diamond), cefotiam (gray circle), cefmetazole (gray triangle), cefazolin (white circle), or cephalexin (white triangle). Each symbol represents rMrp2-mediated transport determined by subtracting the uptake by control vesicles from that by rMrp2-expressing vesicles. Each point and vertical bar represents mean ± S.E.M. (n = 3–6). The solid lines and a broken line are those fitted on the assumption of one or two transport components for [3H]E217βG transport, respectively. B, uptake of [3H]E217βG (10 μM) for 3 min was determined at 37°C in the presence of ATP and each unlabeled compound (200 μM, except for cyclosporin A, which was used at 20 μM). Rat (▪), dog ( ), and human (□) MRP2-mediated transports were determined by subtracting the uptake by control vesicles from that by MRP2-expressing vesicles, respectively. Each point and vertical bar represents mean ± S.E.M. (n = 3–4).
), and human (□) MRP2-mediated transports were determined by subtracting the uptake by control vesicles from that by MRP2-expressing vesicles, respectively. Each point and vertical bar represents mean ± S.E.M. (n = 3–4).
Transport of Cephalosporins by rBcrp and hMRP2. The biliary excretion of cefoperazone observed in EHBRs implied that there is a contribution of some transporter(s) other than Mrp2 to the excretion (Fig. 2C). Therefore, we also examined possible transport of cephalosporins by rBcrp, using membrane vesicles prepared from Sf9 cells transfected with the rBcrp gene. Uptake of cefoperazone, cefbuperazone, cefpiramide, cefotetan, ceftriaxone, cefotiam, cefamandole, and cefazolin by rBcrp-expressing vesicles was significantly higher than that by control vesicles (Fig. 3B), suggesting that these compounds are substrates of rBcrp. The rBcrp-mediated uptake, assessed by the subtraction of the uptake by control vesicles from that by rBcrp-expressing vesicles, tended to be higher for high molecular weight compounds (Fig. 4B).
The uptake of cephalosporins by hMRP2-expressing vesicles was also examined, and uptake of cefoperazone, cefbuperazone, cefpiramide, cefotetan, ceftriaxone, and cefotiam was significantly higher than that in control vesicles (Fig. 3C). The hMRP2-mediated uptake tended to be higher for high molecular weight compounds such as cefoperazone, cefpiramide, cefotetan, and ceftriaxone (Figs. 3C and 4C). Recovery of cefoperazone, cefpiramide, and ceftriaxone in the urine after intravenous administration in humans was less than that of the other compounds, according to the literature (Table 3), implying that these compounds are excreted into the bile in humans.
Pharmacokinetic parameters of β-lactam antibiotics in humans
The recognition of cefoperazone by hMRP2 was also confirmed by means of a [3H]E217βG uptake inhibition study: cefoperazone inhibited the uptake of [3H]E217βG by human MRP2 and dog MRP2 (Fig. 6B). The inhibition was also observed by bromosulfophthalein for the [3H]E217βG uptake by rat, dog, and human MRP2 (Fig. 6B).
Discussion
The mechanism(s) underlying the molecular weight threshold in biliary excretion of organic anions (Hirom et al., 1972; Levine, 1978) remains to be established, although xenobiotic transporters are likely to be involved. If recognition by such transporters plays an important role in determining the molecular weight threshold, permeability clearance across the basolateral and/or luminal membranes in the liver and/or kidney should correlate with either the fraction of dose recovered into the bile or the molecular weight. In the present study, therefore, we first searched the literature to collect information on the permeability clearance of β-lactam antibiotics and its possible relation to molecular weight. According to the literature, CLbile, liver, which represents permeability clearance across the bile canalicular membranes, was correlated with molecular weight (Fig. 1C), indicating that permeability across this membrane plays a key role in the molecular weight dependence observed in the biliary excretion of cephalosporins (Fig. 1A). This finding was consistent with our previous observation that affinity for the transport mechanism across bile canalicular membranes determines the biliary clearance of cephalosporin antibiotics (Tamai et al., 1990).
The transport mechanism for cephalosporins across the bile canalicular membranes in rats was further analyzed using EHBRs in the present study. Cefoperazone was excreted mainly into the bile after i.v. injection in SDRs, but the biliary excretion was greatly reduced in EHBRs (Fig. 2C). Cephalexin is mainly recovered in the urine (Fig, 2F), and its biliary excretion was quite minor but again was lower in EHBRs than in SDRs (Fig. 2D). These results suggest that Mrp2 is predominantly responsible for the transport of cephalosporins across bile canalicular membranes. This hypothesis was further supported by the observation of rMrp2-mediated transport of several cephalosporins (Fig. 3) and by the concentration-dependent inhibition of rMrp2-mediated transport of [3H]E217βG by the cephalosporins (Fig. 6A). The rMrp2-mediated transport tended to be higher for the higher molecular weight compounds (Fig. 4A), indicating that Mrp2 is involved in the molecular weight dependence of the biliary excretion of cephalosporin antibiotics. In addition, the IC50 values obtained in the inhibition study (Fig. 6A) tended to be lower for the higher molecular weight compounds (see Results). Thus, the affinity of the cephalosporins for rMrp2 showed molecular weight dependence.
The rMrp2-mediated transport of cefoperazone exhibited saturation (Fig. 5), and cefoperazone inhibited [3H]E217βG transport (Fig. 6). The present study has thus provided the first evidence that rMrp2 accepts cefoperazone and other several cephalosporins, including cefbuperazone, cefpiramide, and ceftriaxone, as substrates. In addition, transport studies using hMRP2-expressing vesicles indicated that these compounds are also substrates of hMRP2 (Fig. 3C). A comparison of the transport activity for cephalosporins between rats and humans did not reveal any marked species difference in substrate recognition specificity (Fig. 3, A and C), and the rMrp2- and hMRP2-mediated transport activities both exhibited similar molecular weight dependence: cephalosporins with molecular weight higher than 500 were most effectively transported (Fig. 4, A and 4). However, uptake of cefotetan by rMrp2-expressing vesicles was similar to that in control vesicles, whereas uptake by hMRP2-expressing vesicles was much higher than the control value (Fig. 3, A and C). In addition, the inhibition profiles exhibited species differences in the inhibition potential of some compounds among rat, dog, and human MRP2 (Fig. 6B). Therefore, further study of the species difference in MRP2-mediated transport of cephalosporins seems to be necessary, as previously noted for the transport of other substrates (Ninomiya et al., 2005).
The biphasic inhibition curve of cefoperazone described in Fig. 6A indicates that cefoperazone may inhibit two binding sites that mediate [3H]E217βG transport. It has already been demonstrated that rMrp2 has two sites for [3H]E217βG transport, supporting our data (Ninomiya et al., 2005). Additionally, the presence of multiple recognition sites on rMrp2 has been suggested for other substrates (Ito et al., 2001). The remarkable difference between IC50, high and IC50, low (6.66 μM and 3.88 mM) suggests that cefoperazone has different affinity for the two binding sites for [3H]E217βG transport. On the other hand, the saturation curve of cefoperazone transport was monophasic (Fig. 5), and the Km value (70.9 μM) for cefoperazone transport was different from IC50, high and IC50, low. This finding may suggest that cefoperazone is transported by site other than the two binding sites for [3H]E217βG transport, and further studies are necessary to identify the binding site for each compound.
The major elimination organ for therapeutic agents can influence their pharmacological and/or toxicological properties in patients. For example, drugs that are predominantly eliminated via urinary excretion may exhibit severe adverse effects in patients with renal insufficiency owing to reduced systemic clearance and consequent higher exposure in plasma and peripheral organs. Therefore, it is important to estimate the contribution ratios of biliary and renal clearances to the overall systemic elimination in patients at an early stage during drug development. The present findings support the idea that MRP2-expressing vesicles would be useful as a screening system to estimate the transport efficiency of organic anions across the bile canalicular membranes. It is noteworthy that cefoperazone, cefpiramide, and ceftriaxone exhibited higher hMRP2-mediated uptake (Fig. 3C), and their urinary recoveries were relatively smaller in humans (Table 3). On the other hand, hMRP2 mediated the transport of cefotetan (Fig. 3C), which is mainly excreted into the urine (Table 3). Unique characteristics of cefotetan transport were also observed in rats: the rMrp2-mediated transport was minimal (Fig. 3A), but the CLbile, liver of cefotetan was close to that for other biliary excretion-type compounds, such as cefoperazone (Table 1). Thus, MRP2 may not be the only transporter involved in the biliary excretion of all of the cephalosporins. Indeed, cephalosporins are known to be recognized as substrates by various types of transporters other than MRP2, including organic anion transporters 1 and 3 (Takeda et al., 1999; Ueo et al., 2005), organic cation/carnitine transporter 2 (Ganapathy et al., 2000), oligopeptide transporters 1 and 2 (Sai et al., 1996; Ganapathy et al., 1997; Shen et al., 2007), and MRP4 (Ci et al., 2007). Possible involvement of these transporters in the elimination of at least some cephalosporins from the systemic circulation should also be considered.
Cefoperazone was recovered in the bile in EHBRs (Fig. 2C), indicating that some transporter(s) other than Mrp2 is involved in its transport across the bile canalicular membranes. The present study has newly identified cefoperazone, cefbuperazone, cefpiramide, cefotetan, ceftriaxone, cefotiam, cefamandole, and cefazolin as rBcrp substrates (Fig. 3B). In addition, the absolute values for the uptake by rBcrp (expressed as microliters per milligram of protein per minute) were much higher than those for the uptake by rMrp2 in Sf9 membrane vesicles (Fig. 3, A and B), although such difference in the uptake by Sf9 vesicles between rBcrp and rMrp2 does not necessarily represent relative contribution of each transporter in vivo. Interestingly, transport by rBcrp also tended to be higher for higher molecular weight compounds (Fig. 4B). It was recently reported that Bcrp is involved mainly in biliary excretion of conjugated metabolites in mice (Zamek-Gliszczynski et al., 2006b). Thus, this transporter may be another contributor to the phenomenon of molecular weight-dependent excretion. However, the uptake of cefbuperazone by rMrp2- and rBcrp-expressing membrane vesicles was not so remarkable compared with that of cefoperazone and cefpiramide (Fig. 3). Nevertheless, CLbile, liver values of these three compounds are relatively higher, and substantial amounts (60–80% of dose) are excreted into the bile in rats in vivo (Table 1). This means that Mrp2 and Bcrp cannot entirely explain the molecular weight-dependent biliary excretion of cephalosporins. Possible involvement of other transporters on bile canalicular membranes, such as P-glycoprotein and/or bile salt export pump, should also be considered to understand molecular mechanism for the biliary excretion of all the cephalosporins.
Acknowledgments
We thank Toshiyuki Takeuchi and Lica Ishida for performing in vivo studies and technical assistance, respectively. We also thank Dr. Tatsuya Higashi for fruitful discussions regarding quantification with LC-tandem mass spectrometry.
Footnotes
-
This study was supported in part by a grant-in-aid for scientific research provided by the Ministry of Education, Science and Culture of Japan.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.107.019125.
-
ABBREVIATIONS: OATP/Oatp, organic anion-transporting polypeptide; NTP, sodium/phosphate transporter; MRP/Mrp, multidrug resistance-associated protein; BCRP/Brcp, breast cancer resistance protein; EHBR, Eisai hyperbilirubinemic rat; E217βG, estradiol-17β-d-glucuronide; r, rat; h, human; SDR, Sprague-Dawley rat; LC, liquid chromatography; AUC, area under the curve; AUCM, area under the first-moment curve; MOPS, 4-morpholinepropanesulfonic acid.
- Received November 8, 2007.
- Accepted March 12, 2008.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}