


Abstract
Studies were designed to quantitatively assess the mRNA expression of 1) 10 cytochrome P450 (P450) enzymes in human cornea, iris-ciliary body (ICB), and retina/choroid relative to their levels in the liver, and of 2) 21 drug transporters in these tissues relative to their levels in human small intestine, liver, or kidney. Potential species differences in mRNA expression of PEPT1, PEPT2, and MDR1 were also assessed in these ocular tissues from rabbit, dog, monkey, and human. P450 expression was either absent or marginal in human cornea, ICB, and retina/choroid, suggesting a limited role for P450-mediated metabolism in ocular drug disposition. In contrast, among 21 key drug efflux and uptake transporters, many exhibited relative expression levels in ocular tissues comparable with those observed in small intestine, liver, or kidney. This robust ocular transporter presence strongly suggests a significant role that transporters may play in ocular barrier function and ocular pharmacokinetics. The highly expressed efflux transporter MRP1 and uptake transporters PEPT2, OCT1, OCTN1, and OCTN2 may be particularly important in absorption, distribution, and clearance of their drug substrates in the eye. Evidence of cross-species ocular transporter expression differences noted in these studies supports the conclusion that transporter expression variability, along with anatomic and physiological differences, should be taken into consideration to better understand animal ocular pharmacokinetic and pharmacodynamic data and the scalability to human for ocular drugs.
As the vital organ of vision, the eye is equipped with sophisticated anatomic structures to maintain a highly regulated and confined environment for its function. This includes various ocular barriers, i.e., cornea epithelium, blood-aqueous barrier (BAB), and blood-retina barrier (BRB), that govern the entry or exit of nutrients and xenobiotics to ocular tissues in either the anterior or posterior part of the eye (Hornof et al., 2005). For topically administered drugs, the cornea forms the primary rate-limiting permeability barrier to compound absorption into the anterior chamber of the eye. Additional barriers to reaching target sites in the anterior chambers include drug elimination through aqueous humor outflow and entering the uveal blood circulation via the BAB. For pharmaceuticals delivered directly to the back of the eye, their pharmacological effect and duration are not only controlled by the formulation, physicochemical properties, or device used but also by their interaction with the retina, which determines drug clearance into the systemic circulation. Conversely, BAB and especially BRB also restrict the entry of drugs and metabolites from systemic circulation into the eye.
Unlike the extensive body of knowledge that exists for drug transporters and metabolizing enzymes found in major drug disposition organs, that is, small intestine, liver, and kidney, knowledge about the expression and function of these systems in ocular tissues is very limited despite the potentially important role they may play in ocular drug disposition (Duvvuri et al., 2004; Mannermaa et al., 2006). Furthermore, available expression and function data have been largely limited to lower species and cell lines. The studies described here, by using real-time quantitative polymerase chain reaction (PCR) methods, were primarily aimed at providing the first systematic profiling of mRNA expression levels of key cytochromes P450 (P450s) and drug transporters that may be essential constituents of ocular barriers in such ocular tissues as cornea, iris-ciliary body (ICB) (for BAB), and retina/choroid (for BRB). In addition, the potential cross-species differences for PEPT1, PEPT2, and MDR1 mRNA expression in rabbit, dog, monkey, and human ocular tissues were also studied. The selection of these transporters for cross-species investigation was motivated by the fact that their functional or molecular presence in the eye has been either shown or suggested in the literature (Anand and Mitra, 2002; Dey et al., 2003; Ocheltree et al., 2003) and that they are well documented drug transporters that have been cloned from all four species intended for comparison.
Materials and Methods
Human and Animal Ocular Tissues. Human whole globes, enucleated within 6 h of death from human donors (age range 60–93 years old) were obtained from the San Diego Eye Bank. On arrival, the eyes were immediately dissected, and the following tissues were collected: cornea, ICB, and retina/choroid. The tissues were snap-frozen in liquid nitrogen and stored at -80°C until processing. The cornea, ICB, and choroid/retina pooled from two or three individuals were used for this study. All the tissues were from donors free of ocular disease and other systemic complications and were processed in accordance with the guidelines outlined in the Declaration of Helsinki.
Female Dutch Belted rabbits (1.5–2.0 kg) from Myrtle's Rabbitry (Thompson Station, TN), beagle dogs (male and female, weighing 9–13 kg) from Marshall Farms (North Rose, NY), and cynomolgus monkeys from BRF (a subsidiary of Charles River Laboratories, Wilmington, MA) were used in this study. Before tissue collection, all the animals underwent physical and ocular examinations and passed with no abnormal findings. The animals were euthanized i.v. with an overdose of sodium pentobarbital (Buethanasia, Butler Corp., Fredericksburg, VA), and the eyes were enucleated immediately. The cornea, ICB, and retina/choroid were then quickly dissected from both eyes of all the species. Samples were snap-frozen in liquid nitrogen and stored at -80°C until processing. The corneas, ICB, and retina/choroid pooled from two or three animals were used for this study. All the animal-related work was conducted under a Pfizer Inc. (New York, NY) Institutional Animal Care and Use Committee–approved protocol in compliance with Animal Welfare Act regulations and the Guide for the Care and Use of Laboratory Animals.
RNA Extraction and cDNA Synthesis. The isolation of RNA was carried out using TRIzol (Invitrogen, Carlsbad, CA) and/or RNeasy kit (Qiagen, Valencia, CA) performed according to manufacturer's protocols. Total RNA from the cornea was extracted with TRIzol after homogenization with an Ultra-Turrax T8 (IKA, Wilmington, NC). The isolated RNA was then purified via the Qiagen RNeasy kit. Total RNA from other ocular tissues (ICB and retina/choroid) was extracted with the Qiagen RNeasy kit after homogenization with a FastPrep F120 instrument (Qbiogene, Irvine, CA). The isolated RNAs were treated with DNase (DNA-free kit, Ambion, Huntington, UK) to digest genomic DNA. The integrity of RNA was visually assessed by conventional agarose gel electrophoresis, and the concentration was measured by a NanoDrop spectrophotometer (Thermal Fisher Scientific, Wilmington, DE). All the nonocular RNA including small intestine, liver, and kidney were obtained from BioChain Institute (Hayward, CA). Human RNA was obtained from a five-person pool, and rabbit, dog, and monkey RNA were from one animal each. The RNA from all the samples was reverse-transcribed into cDNA using the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA) according to manufacturer's protocol. Parallel control reactions without the addition of reverse transcriptase were performed for each sample.
Real-Time Quantitative PCR. Real-time PCR was performed in 96-well plates using the iCycler iQ system (Bio-Rad, Hercules, CA). Primer and probes were either purchased from Applied Biosystems or designed based on the sequence entries in the GenBank and synthesized from Integrated DNA Technologies (Coralville, IA) (see Tables 1 and 2). The self-designed probes were labeled with a reporter dye (5-carboxyfluorescein) on the 5′-end and quencher dye (BHQ-1) on the 3′-end. The sequences of the designed probes and primers are as follows: rabbit MDR1 [sense primer: TGAACGTGAGGTATCTGCGGGAAA; probe: TGGTGAGTCAGGAACCTGTGTTGTTCGCCA; antisense primer: ATAGCGGACGTTCTCAGCGATTGT; 89 base pairs (bp)], rabbit PEPT1 (sense primer: AGTGGTGAAGTGCATCTGCTTTGC; probe: AGGCACCGCAGTAAGCAGTTTCCCAAGA; antisense primer: AAGCCGCTCGTCGTATTTCTCCTT; 115 bp); rabbit PEPT2 (sense primer: TGAAATCTGTACTCCAGGCAGCGT; probe: TGGGAACATCATCGTGCTTGTTGTGGCACA; antisense primer: AATTCAGCCCACTGTACCAGACCA; 103 bp); rabbit glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (sense primer: TCATTTGAAGGGCGGAGCCAAA; probe: CCATGTTTGTGATGGGCGTGAACCACGAGA; antisense primer: GCAGGATGCGTTGCTGACAATCTT; 127 bp); and dog GAPDH (sense primer: AGTCAAGGCTGAGAACGGGAAACT; probe: TTCCAGGAGCGAGATCCCGCCAACATCAAA; antisense primer: TCCACAACATACTCAGCACCAGCA; 114 bp). Each PCR reaction was run in a total volume of 20 μl, containing cDNA transcribed from 10 ng of total RNA, TaqMan Universal PCR Master Mix, and TaqMan Gene Expression Assay mixes containing specific primers and probes. The concentrations for self-designed primers and probes were 900 and 250 nM, respectively. The PCR was run at 50°C for 2 min, 95°C for 10 min, followed by 40 cycles of 95°C for 15 s and 60°C for 1 min. Triplicate PCR reactions were performed for each sample. The setting of the threshold and baseline was performed according to the manufacturer's instructions.
List of human genes studied in the analysis The table summarizes the systematic gene nomenclature according to the Human Genome Organization consensus, common gene symbols, and Applied Biosystems (ABI) assay I.D. The shaded coding illustrates the rank order of the expression levels among the tissues studied. (Fields marked by “X” indicate absent expression; white fields indicate very low expression; darker shades indicate higher expression). For the P450s, the expression level is normalized by liver level, which is assigned as 100 and indicated by the dark shade. For transporters, the expression level is normalized by the highest among liver, small intestine, or kidney, which is assigned as 100 and indicated by the dark shade.
List of reference and animal genes studied in the analysis The table summarizes the gene name, sequence accession I.D., and ABI assay I.D. For those not available from ABI, probes and primers were designed, and their sequences are listed under Materials and Methods.
Relative Expression Analysis. Quantification of the gene expression was performed using the relative standard curve method. The standard curve was generated by the mixtures of cDNA from the tissues of interest and serially diluted by a 1:4 ratio. A six-point standard curve was made for each gene. The amplification efficiencies were close to 1 for all the genes. Four human housekeeping genes, including β-actin, GAPDH, 18S, and peptidylprolyl isomerase A, were evaluated as potential internal reference genes. It was decided to exclude 18S because it had a much lower Ct value (the Ct value is the cycle time when fluorescence is higher than a defined threshold level) and much larger variance among the tissues studies. The geometric mean of β-actin, GAPDH, and peptidylprolyl isomerase A was used as the normalization factor for the relative quantification of each human gene. To study species difference with respect to the relative expression of PEPT1, PEPT2, and MDR1 mRNA in rabbit, dog, monkey, and human ocular tissues, GAPDH from each species was used as the normalization factor.
Results
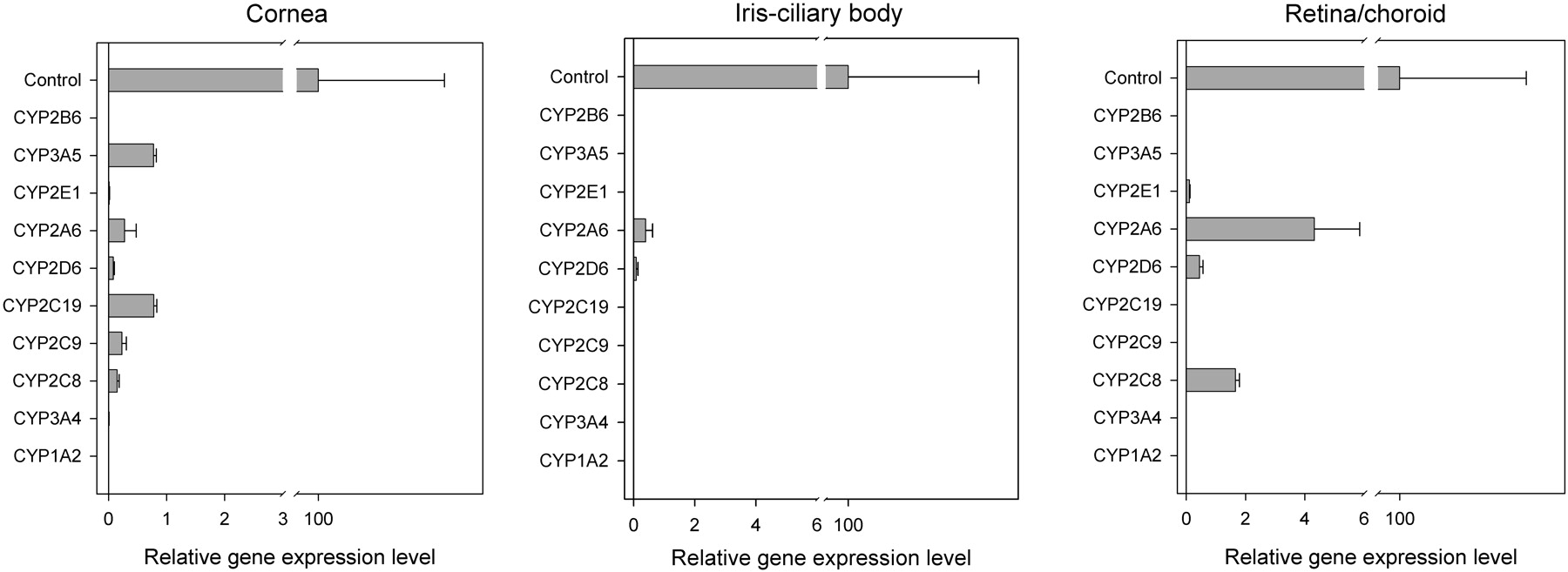
P450 mRNA Expression in Human Ocular Tissues. Each enzyme expression in human ocular tissues was normalized to the corresponding hepatic level. CYP1A2 and CYP2B6 were absent in all the ocular tissues tested. CYP2C9, CYP2C19, and CYP3A5 were absent in both ICB and retina/choroid. CYP3A4 is absent in retina/choroid, and CYP2C8 was absent in ICB. In cornea and ICB, the mRNA level of each detectable P450 gene was less than 1% of the hepatic level. In retina/choroid, the mRNA levels of CYP2D6 and CYP2E1 were also less than 1% of the hepatic level; the mRNA expression levels of CYP2A6 and CYP2C8 were about 4 and 2% of their corresponding liver levels, respectively (Fig. 1).
ATP-Binding Cassette Efflux Transporter mRNA Expression in Human Ocular Tissues. The ascending order of the MDR1 expression in the tissues tested was as follows: cornea < retina/choroid < ICB < liver < kidney < small intestine. The rank order of breast cancer resistance protein (BCRP) expression was small intestine > liver > retina/choroid ≈ ICB ≈ kidney > cornea. The expression of multidrug resistance protein (MRP) 1 was higher in ICB and retina/choroid compared with that observed in small intestine, kidney, and liver with the rank order of ICB > retina/choroid > small intestine > kidney > cornea > liver. The ascending order of MRP2 expression was cornea ≈ ICB < retina/choroid < kidney < small intestine < liver. The rank order of MRP3 expression was liver > kidney ≈ small intestine > retina/choroid > cornea > ICB (Fig. 2).
Uptake Transporter mRNA Expression in Human Ocular Tissues.Peptide transporters. PEPT1 expression was detected at low levels in ICB and cornea but was absent in retina/choroid. PEPT2 expression in both retina/choroid and ICB was comparable with the level in kidney, which was higher than the levels in liver and small intestine. PEPT2 was also expressed in the cornea at a level similar to that observed in the small intestine and liver. Human peptide transporter 1 expression in ocular tissues was very low (Fig. 3A).
Organic anion transporting peptides (solute carrier 21). Organic anion transporting peptide (OATP) 1A2 expression in retina/choroid was roughly 5-fold lower than the level found in the kidney. OATP1B1 and OATP1B3 expression was absent in ocular tissues tested. OATP2B1 was expressed in all the ocular tissue tested, with the highest level observed in ICB, which was roughly 16% of the hepatic level. Its expression levels in retina/choroid and cornea were 6 and 1% of the hepatic level, respectively (Fig. 3B).
Organic cation/anion transporters (solute carrier 22). OCT1 was highly expressed in ocular tissues, especially in retina/choroid, where its level was higher than those expressed in small intestine, kidney, and liver. OCT2 was detected in the ICB at about 8% of the level in the kidney but absent in the cornea and retina/choroid. OCT3 expression levels in the retina/choroid, ICB, and cornea were about 0.3, 2, and 5% of the hepatic level, respectively (Fig. 3C).
OCTN1 was highly expressed in ocular tissues, especially in ICB, where its expression was higher than that found in the small intestine and kidney. OCTN2 was also expressed in all the ocular tissues tested. Its expression level in cornea, ICB, and retina/choroid was about 6, 52, and 59% of the level in the kidney, respectively (Fig. 3C).
None of the organic anion transporters (OATs) tested was detected in the cornea. OAT1 expression in the ICB was about 5% of the level in the kidney. OAT2 was only detected in retina/choroid at a very low level of less than 0.1% of the hepatic level. OAT3 expression level in retina/choroid and ICB was about 2 and 14% of the level in the kidney, respectively (Fig. 3C).
Bile salt cotransporter. ASBT mRNA was absent in all the ocular tissues tested (Fig. 3D).
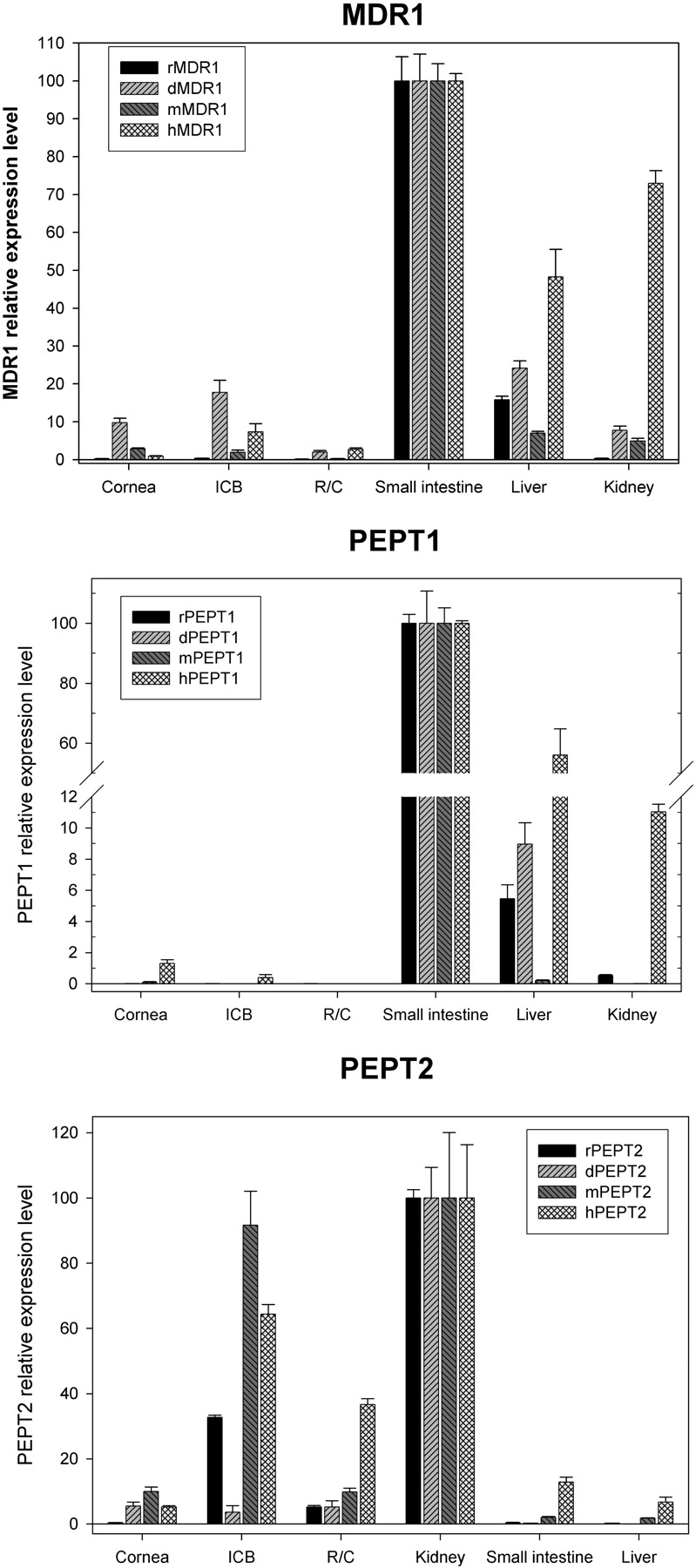
Transporter mRNA Expression Comparison between Rabbit, Dog, Monkey, and Human Ocular Tissues. With respect to transporter mRNA expression in nonocular human tissues, the small intestine had the highest expression level of MDR1 and PEPT1 (Figs. 2 and 3A); the kidney had the highest expression level of PEPT2 (Fig. 3A). This expression pattern was consistent across species (Fig. 4). To facilitate cross-species comparison, the relative level of MDR1 or PEPT1 in each ocular tissue was normalized by the level in small intestine of the corresponding species, whereas the relative level of PEPT2 in each ocular tissue was normalized by the kidney level of the corresponding species.
MDR1 mRNA was detected in all the tested ocular tissues across species. In cornea, the dog seemed to have the highest level, which was 10% of the level in dog small intestine. Human and rabbit cornea appeared to have low levels of MDR1 with less than 1% of the level in the small intestine. In ICB, rabbits seemed to have the lowest level and dog the highest level. MDR1 mRNA expression in rabbit, dog, monkey, and human ICB was 0.4, 18, 2, and 7% of the level in the small intestine, respectively. In retina/choroid, MDR1 levels in rabbit and monkey tissues were similarly low, less than 0.3% of the level in the small intestine; MDR1 levels in dog and human retina/choroid were similar and about 2% of the level in the small intestine (Fig. 4A).
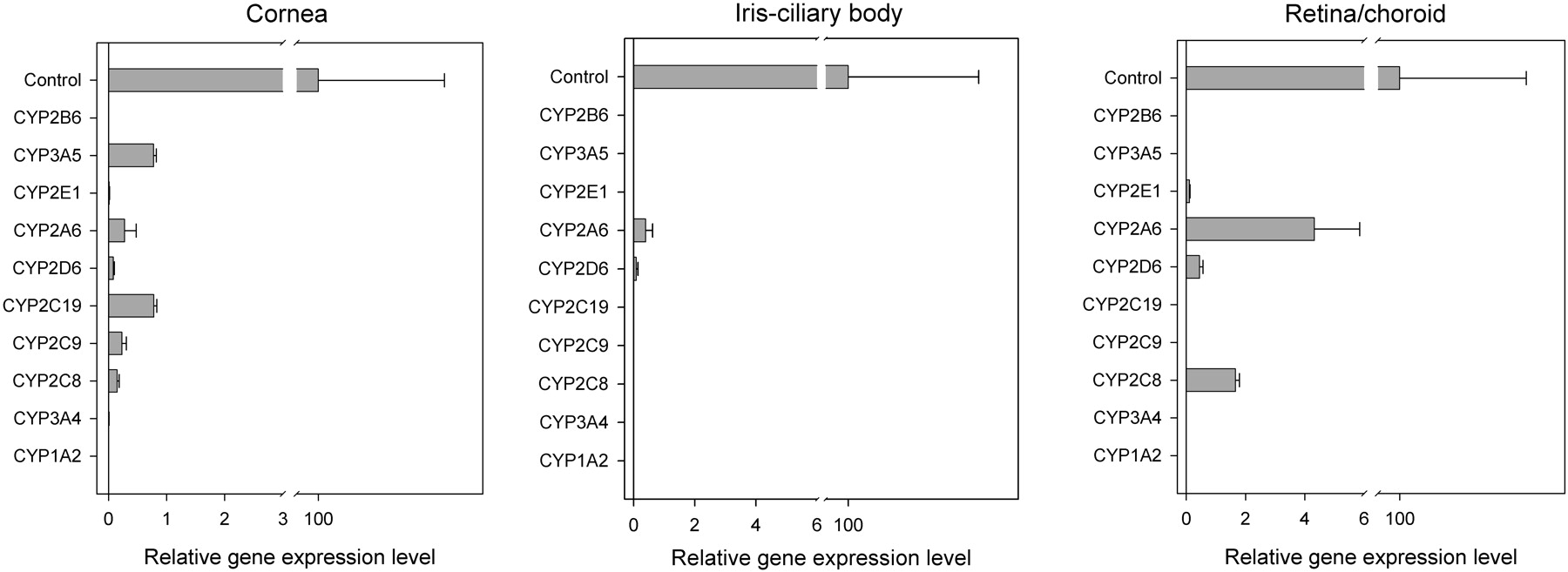
Relative mRNA expression levels of P450 enzymes in human cornea, ICB, and retina/choroid. The expression level of each gene is normalized by its level in the liver, which is named as “Control” and assigned as 100. Data are presented as the mean of triplicates with the S.D. as the error bar. The error bar in the “Control” is the mean value of all the S.D. obtained from the analysis on each gene.

Relative gene expression levels of ATP-binding cassette transporters in human cornea, ICB, retina/choroid, small intestine, liver, and kidney: MDR1, MRP1, MRP2, MRP3, and BCRP. Data are presented as the mean of triplicates with the S.D. as the error bar.
PEPT1 mRNA was detected at low levels in ocular tissues tested from rabbit, dog, monkey, and human. It was not detected in the dog ICB and human retina/choroid. In the cornea, humans seemed to have the highest PEPT1 level, which was about 1.5% of the level in human small intestine (Fig. 4B).
PEPT2 mRNA was detected in all the tested ocular tissues across species. In the cornea, rabbit seemed to have the lowest level, which was less than 0.5% of the level in rabbit kidney. Dog, monkey, and human cornea had higher levels, ranging from 5 to 10% of the kidney level. In ICB, dog displayed the lowest level of PEPT2, which was 3.6% of the level in dog kidney. PEPT2 levels in rabbit, monkey, and human ICB were higher at 32, 92, and 64% of the level in the kidney, respectively. In the retina/choroid, the highest levels were observed in the human (37% of the level in the kidney), whereas rabbit, dog, and monkey had lower levels, ranging from 5 to 10% of the level found in the kidney.

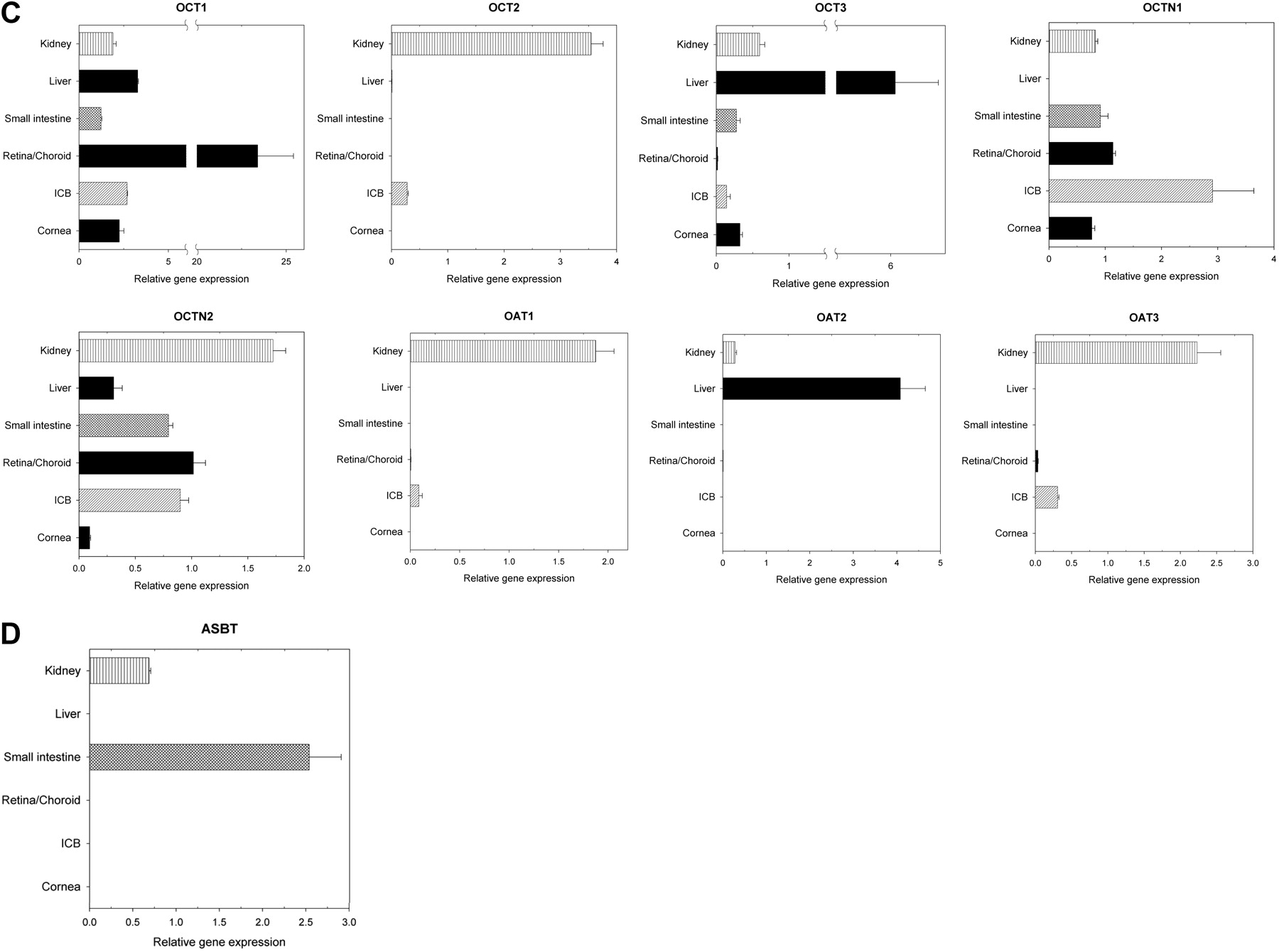
Relative gene expression levels of solute carrier (SLC) transporters in human cornea, ICB, retina/choroid, small intestine, liver, and kidney. A, peptide transporters; B, OATPs (SLC21). C, organic cation/anion transporters (SLC22); D, bile salts cotransporter. Data are presented as the mean of triplicates with the S.D. as the error bar.
Discussion
Unlike the liver, small intestine, and kidney, the mammalian eye is seldom considered as a major disposition organ after drug dosing. Yet, for ophthalmic drugs, it is important to understand ocular drug disposition as the drug or metabolite concentration in the ocular tissues often dictates its pharmacodynamic effect. Ocular tissues evidently display metabolizing activities toward xenobiotic and endogenous substrates (Duvvuri et al., 2004). It is also recognized that drug transporter activity is an essential component for ocular barrier function (Mannermaa et al., 2006). However, information on the expression profile of drug-metabolizing enzymes and transporters in the human eye is largely absent. In the presented studies, quantitative real-time PCR methods were used to obtain the relative expression profiles of the P450 enzymes and various transporter systems in the human eye. The expression of three selected transporters was also compared across species. To facilitate data interpretation, each P450 enzyme expression in the ocular tissues was presented as the relative level after being normalized by its level in the liver, which is the major metabolizing organ. Similarly, each transporter expression level in ocular tissues was compared and rank-ordered with its level in the liver, small intestine, and kidney, all of which are organs of rich drug transporter expression. These studies compared the relative mRNA abundance of a given enzyme or transporter in different tissue types but did not compare mRNA expression levels between different enzymes or transporters. In addition, the relative expression level of a given enzyme or transporter in humans was qualitatively described as high, moderate, low, very low, or absent. It should also be noted that a limitation of using mRNA level as the surrogate for gene expression is that mRNA levels are not necessarily correlated with the functional protein expression, which may be a better indicator for the functional role of these metabolizing enzymes and transporters in ocular drug disposition. Finally, the human ocular tissues studied here are free from ocular diseases but are from aged donors. As most ocular diseases occur in the elderly population, the expression data in this population may be most relevant in understanding drug disposition in the eye. Nevertheless, the effect of disease status and age on the expression of metabolizing enzymes and transporters is beyond the scope of this study.
P450 mRNA Expression in Human Ocular Tissues. The mRNA expression of the 10 studied P450 enzymes in the eye was much less than their levels in the liver and in some instances was absent in ocular tissues. This limited mRNA expression suggests that P450-mediated metabolism may contribute, but perhaps not to a significant degree, to the overall metabolism capacity of the eye. These data are consistent with a very low turnover rate when P450 probe substrates were incubated in various ocular tissue homogenates (data not shown). Although not profiled in this article, other metabolizing enzymes including esterases, peptidases, and ketone reductases are known to have functional expression in various ocular tissues across species (Lee et al., 1988; Mannermaa et al., 2006). When it comes to molecular design of ophthalmic drug, the focus should be on the specificity and capacity of these non-P450 enzymes. One of the examples is the prodrug approach that successfully exploits esterases in ocular drug design (Shirasaki, 2007).
Drug Transporter mRNA Expression in Human Ocular Tissues. This article reports the relative expression profiles of 21 drug transporters in human cornea, ICB, retina/choroid, liver, kidney, and small intestine. This is an extensive but by no means complete list of important drug transporters. The rank-order expression levels of these transporters in the liver, kidney, and small intestine in this study largely conform to previously reported data using a similar real-time PCR approach (Hilgendorf et al., 2007). This interlab consistency further validates using real-time PCR as a viable method to relatively quantify mRNA expression levels in different tissue types despite the potential differences associated with this methodology, including tissue resource, mRNA extraction method, PCR conditions, selection of housekeeping genes for normalization (Vandesompele et al., 2002), et al.

Relative gene expression levels of transporter genes in cornea, ICB, retina/choroid of rabbit, dog, monkey, and human. MDR1, normalized by the level in the small intestine of the same species; PEPT1, normalized by the level in the small intestine of the same species; PEPT2, normalized by the level in the kidney of the same species. Data are presented as the mean of triplicates with the S.D. as the error bar.
Cornea. Considering that the cornea is in direct contact with the external environment, it is possible for this tissue to express efflux transporter as a part of its protective mechanism. There are conflicting reports in the literature regarding ATP-binding cassette efflux transporter expression in human cornea. For example, one study showed there was no detectable MDR1 mRNA expression in human cornea (Becker et al., 2007), whereas another reported the mRNA expression of MDR1 in human cornea, as well as P-glycoprotein expression by Western blot in a human corneal cell line (Dey et al., 2003). In the current study, MDR1 mRNA expression was found to be very low, which may help explain the literature inconsistency as many variables in real-time PCR protocols (including cycle number and intersubject variability of source mRNA) could lead to conflicting results, especially for genes with low copy numbers. Similar to MDR1, both BCRP and MRP2 also showed very low expression levels in the human cornea relative to their levels in the liver or small intestine, whereas there were moderate MRP1 and low MRP3 expression levels in the human cornea. Because these major drug efflux transporters tend to have overlapping or complementary substrate specificity (Food and Drug Administration, http://www.fda.gov/cder/drug/drug-Interactions/tableSubstrates.htm#major), MRP3 and especially MRP1 in human cornea may be the primary efflux transporters to offer potential barrier protection despite the low abundance of MDR1, MRP2, and BCRP in human cornea. These data suggest that designing around MRP1 efflux may be a key consideration to enhance ocular penetration if drug efflux is a concern for corneal permeability. Besides the presence of efflux transporters, human cornea also has low to moderate expression of PEPT1, PEPT2, and some organic cation/anion transporters (OCT1, OCT3, OCTN1, and OCTN2). Contrary to efflux mechanism, these uptake transporters offer potential absorptive transport pathways for topically dosed drug molecules. For example, the conversion of antiviral drugs acyclovir and ganciclovir into their L-valyl esters significantly improved their corneal permeability, presumably by targeting the dipeptide transporters expressed in the cornea (Anand and Mitra, 2002). The data presented here suggest, by their presence, both PEPT1 and PEPT2 may participate in this absorptive mechanism for their drug substrates after topical dosing to the cornea.
ICB. ICB forms the BAB. Its barrier function may not be as complete as the BRB because the content exchange between plasma and aqueous humor is less restricted. This study reveals that numerous efflux and uptake transporters are expressed in ICB: MRP1 and OCTN1 mRNA levels are comparable with those levels found in small intestine and kidney; PEPT2 level is comparable with that in the kidney; other transporters including OCT1, OCTN2, and OAT3 are also substantially expressed in this tissue. The presence of these efflux and uptake transporters may provide an additional active elimination pathway for drug removal from aqueous humor to systemic circulation or vice versa, which works in concert with the less restricted passive pathway and aqueous humor turnover to eliminate drugs from the anterior chamber (Hornof et al., 2005). This may explain why some large and lipophilic drugs have clearance values exceeding the rate of aqueous humor turnover (Schoenwald, 2003).
Retina/Choroid. The retina is composed of neural retina and retinal pigment epithelium cells, which are adjacent to the choroid. The BRB includes the endothelial cells of retinal blood vessels as the inner BRB and the retinal pigment epithelial cells as the outer BRB. The BRB prevents plasma contents including drug molecules from entering into the retina. The retina is in direct contact with vitreous humor; thus, retinal drug concentration and elimination half-life for drug molecules administered into the vitreous humor are partially determined by retinal penetration, which can be governed by both passive and active processes. As summarized in a recent review article (Mannermaa et al., 2006), there is rich transporter expression in the BRB, but most studies were conducted in preclinical species and immortalized cell lines. This study offers a more complete profiling of transporter expression in human retina with a few noteworthy findings. All five efflux transporters tested are expressed in human retina, especially MRP1, which has a high relative expression level. This observation is consistent with the previous study showing both mRNA and protein expression of MRP1 in both ARPE-19 and primary human retinal pigment epithelia cells (Aukunuru et al., 2001). PEPT1 mRNA could not be detected in human retina; however, there is moderate PEPT2 expression in this tissue, which suggests that PEPT2 may be the dominant oligopeptide transporter in human BRB, similar to the role of PEPT2 in the blood-brain barrier (Shen et al., 2007). OCT1 seems to have the highest relative expression level in retina compared with all the other tissues tested. OCT1 expression in ocular tissue has not been reported up to now. As many drug molecules are substrates or inhibitors of human OCT1, the functional implication of a highly expressed OCT1 in human retina remains to be further defined for ophthalmic drugs. Besides OCT1, other organic anion and cation transporters are also expressed in human retina (Fig. 3C). Further investigation of drug transporter expression regarding location, orientation, and transport capacity in both inner and outer BRBs will be essential to understand the role of drug transporters in retinal disposition and how drug-transporter interactions may be able to influence ophthalmic drug design.
Species Differences in Ocular Transporter Expression. The ocular mRNA expression of MDR1, PEPT1, and PEPT2 was normalized across species and presented as relative to levels in the small intestine (MDR1 and PEPT1) or kidney (PEPT2). Potential species differences were observed for the mRNA expression of these three transporters in ocular tissues. The implications of these observed differences are illustrated by the results found in the cornea. In the cornea, dog MDR1 mRNA expression is about 10% of its expression in dog small intestine, and human MDR1 mRNA level is about 1% of its expression in human small intestine. These data suggest that the corneal MDR1 expression, as well as the MDR1-mediated efflux potential, may be different between dogs and humans. PEPT1 mRNA level in rabbit cornea is less than 0.01% of that found in rabbit small intestine, much lower than the relative level (1.3%) of PEPT1 in human cornea normalized by the level in human small intestine. Similarly, PEPT2 mRNA level in rabbit cornea is about 0.4% of its level in rabbit kidney, much lower than the relative level (5.2%) of human corneal PEPT2 normalized by human kidney PEPT2 level. These data suggest that corneal PEPT1 and PEPT2 expression, as well as the PEPT1/PEPT2-mediated absorptive uptake, may be different between rabbits and humans. These potential differences remain to be further investigated by additional protein expression data and functional comparison using probe transporter substrates. This transporter expression variability in the eye, when better understood, may help us understand the potential limitations of different animal models with respect to ocular pharmacokinetics and perhaps help refine animal pharmacokinetic/pharmacodynamic information for better human dose prediction.
In conclusion, this study represents the most comprehensive analysis of P450 enzymes and drug transporter expression in human ocular tissues to date. It provides the molecular evidence for the potentially limited role of P450-mediated metabolism in the eye and the potentially important role of certain drug transporters in ocular drug disposition. In addition, this report shows the importance of studying the species difference of drug transporters in the eye. Future efforts will further investigate the role of drug-metabolizing enzymes and transporters in ocular drug disposition, which may provide guidance on how to optimize drug disposition properties in ophthalmic drug design.
Acknowledgments
We thank Chunze Li and Deepak Davie for insightful scientific discussions, Mike Zientek for critical reading of the manuscript, and Terri Vandegiessen, Yanfang Liu, and Helen Cho for technical assistance.
Footnotes
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.108.021121.
-
ABBREVIATIONS: BAB, blood-aqueous barrier; BRB, blood-retina barrier; PCR, polymerase chain reaction; P450, cytochrome P450; ICB, iris-ciliary body; MDR, multidrug resistance; bp, base pair; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; BCRP, breast cancer resistance protein; MRP, multidrug resistance protein; OATP, organic anion transporting peptide; OCT, organic cation/anion transporter; OAT, organic anion transporter; SLC, solute carrier.
- Received February 19, 2008.
- Accepted April 7, 2008.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}