


Abstract
Contrast-enhancing magnetic resonance imaging with the liver-specific agent gadolinium-ethoxybenzyl-diethylenetriamine pentaacetic acid (Gd-EOB-DTPA) has been shown to improve the detection rate of focal lesions. There is evidence from preclinical studies that multidrug organic anion transporters are involved in hepatic uptake of Gd-EOB-DTPA. Therefore, we evaluated affinity of the contrast agent to human organic anion-transporting polypeptides (OATP1B1, OATP1B3, OATP2B1) and to the Na+/taurocholate cotransporting polypeptide (NTCP) using stable transfected human embryonic kidney (HEK) 293 cells. In competition assays, Gd-EOB-DTPA inhibited the uptake of bromosulfophthalein (BSP) by OATP1B1 (IC50 = 0.6 mM) and OATP1B3 (IC50 = 0.4 mM). In comparison, the IC50 values for rifampicin were 11.9 (OATP1B1), 1.4 (OATP1B3), and 80.5 μM (OATP2B1), respectively. Uptake of BSP by OATP2B1, uptake of taurocholic acid by NTCP, and viability of all HEK cells were not influenced by Gd-EOB-DTPA in concentrations up to 10 mM. In uptake assays using a new liquid chromatography-tandem mass spectrometry method for quantification, Gd-EOB-DTPA was a substrate for OATP1B1 (Km = 0.7 mM, Vmax = 10.5 pmol/mg × min), OATP1B3 (Km = 4.1 mM, Vmax = 22.7 pmol/mg × min), and NTCP (Km = 0.04 mM, Vmax = 1.4 pmol/mg × min). The uptake by OATP2B1 was not different from the vector control. In conclusion, Gd-EOB-DTPA is a substrate of the liver-specific OATP1B1, OATP1B3, and NTCP.
Magnetic resonance imaging (MRI) enables a contrast resolution in clinical diagnostics that facilitates the distinction between normal and pathological tissues. In addition, contrast-enhancing imaging has increased the accuracy for identification and characterization of pathological findings. For instance, gadolinium-ethoxybenzyl-diethylenetriamine pentaacetic acid (Gd-EOB-DTPA)-enhanced liver imaging has been shown to significantly improve the detection rate of focal liver lesions (Mühler et al., 1992; Schuhmann-Giampieri et al., 1992). The liver-specific contrast agent Gd-EOB-DTPA is rapidly extracted after intravenous administration from portal blood into hepatocytes by unknown carrier(s) and excreted into bile canaliculi via the multidrug resistance-associated protein ABCC2 (formerly named MRP2) (Muhler et al., 1993). There is ample evidence for the hepatic uptake to be mediated by liver specific-uptake transport proteins such as the Na+/taurocholate cotransporting polypeptide (NTCP) and/or members of the organic anion-transporting polypeptides (OATP) family (Meier et al., 1997; Kim, 2003). In rats, it was shown that Gd-EOB-DTPA exploits the transport system used by hepatocytes for uptake of bilirubin (Schuhmann-Giampieri et al., 1993). Gd-EOB-DTPA is a substrate of rat Oatp1a1 but not of human OATP1A2, as confirmed by a transport study using Xenopus laevis oocytes (van Montfoort et al., 1999). Moreover, known substrates of human and rodent OATP/Oatp compete with hepatic enhancement of Gd-EOB-DTPA, such as bromosulfophthalein (BSP) and rifampicin (Clément et al., 1992; van Montfoort et al., 1999; Kato et al., 2002). Finally, enhancement of Gd-EOB-DTPA is highly correlated to the expression level of human OATP1B3 in hepatocellular carcinoma tissue (Narita et al., 2009). Thus far, however, cellular uptake of Gd-EOB-DTPA in cells expressing human uptake carriers for anionic amphiphilic compounds has not been described. Therefore, we investigated affinity of Gd-EOB-DTPA to the most promising candidates (OATP1B1, OATP1B3, OATP2B1, and NTCP) using stable transfected human embryonic kidney (HEK) 293 cells.
Materials and Methods
Cell Culture.
HEK293 cells were purchased from the European Collection of Cell Cultures (Salisbury, United Kingdom) and were grown in minimal essential medium supplemented with 10% fetal bovine serum, 2 mM l-glutamine, 2 mM nonessential amino acids, 100 units/ml penicillin, and 100 μg/ml streptomycin (PAA, Coelbe, Germany) at 37°C, 95% humidity, and 5% CO2.
Cloning and Transfection.
OATP2B1 (GenBank accession no. NM_007256.3) was amplified as described previously (Grube et al., 2006). NTCP (GenBank accession no. NM_003049.2) was cloned as specified before (Hagenbuch and Meier, 1994). The full-length OATP1B1 (GenBank accession no. NM_006446.4) and OATP1B3 (GenBank accession no. NM019844) were amplified using reverse-transcribed RNA from human liver (primer OATP1B1: forward primer, 5′-GCCATGGACCAAAATCAACATTTG-3′ and reverse primer, 5′-TTAACAATGTGTTTCACTATCTGCC-3′; primer OATP1B3: forward primer, 5′-TAGTTTAATAATGGACCAACATCAACA-3′ and reverse primer, 5′-TGTTAGTTGGCAGCAGCATT-3′). Coding sequences of the transporters were cloned into the retroviral expression vector pQCXIN (Clontech, Saint-Germain-en-Laye, France). The HEK293 cells were infected according to the manufacturer's instructions and were selected by 500 μg/ml neomycin. Resistant cell clones were characterized by Western blot analysis and immunofluorescence staining.
Immunoblot Analysis and Immunofluorescence Microscopy.
For Western blot analysis, 50 μg of cell lysate were loaded onto a 10% sodium dodecyl sulfate polyacrylamide gel, and apparent molecular masses were assessed by comparison with the PageRulerPrestained Protein Ladder (Fermentas, St. Leon-Rot, Germany). Immunoblotting was performed using a tank blotting system (Bio-Rad, Munich, Germany). Antibodies against OATP1B1, OATP1B3, OATP2B1, and NTCP as well as the secondary horseradish peroxidase-conjugated goat anti-rabbit antibody (Acris, Herford, Germany) were used as described previously (Kullak-Ublick et al., 1997; König et al., 2000a, 2006; Grube et al., 2006). Immunofluorescence microscopy was performed with the Leica TCS SP5 microscope (Leica Microsystems, Bensheim, Germany). Expression of transporter protein was assessed using the antibodies mentioned previously and with a chicken anti-rabbit antibody conjugated to Alexa Fluor 488 (Invitrogen, Karlsruhe, Germany).
Transport Studies.
All transport studies with HEK293 cells were performed in 24-well plates (Greiner Bio-One, Frickenhausen, Germany) after cultivation in full growth medium for 2 days (initial density of 300,000 cells/well) at 37°C using a standard incubation buffer (Seithel et al., 2007). To measure activities of OATPs and NTCP, [3H]bromosulfophthalein (14 Ci/mmol; Hartmann Analytic, Braunschweig, Germany) and [3H]taurocholic acid (TA) (4.6 Ci/mmol; PerkinElmer Life and Analytical Sciences, Waltham, MA), respectively, were dissolved with unlabeled BSP or TA (Sigma-Aldrich, Taufkirchen, Germany) to reach final concentrations of 0.05 μM for OATP1B1, 1 μM for OATP1B3 and OATP2B1, and 10 μM for NTCP. After incubation for 5 min, the cells were washed with ice-cold phosphate-buffered saline and disintegrated using 0.5% Triton X-100 and 0.5% sodium deoxycholate. Aliquots were mixed with 2 ml of scintillation cocktail (Rotiszint eco plus; Roth, Karlsruhe, Germany) and measured using a scintillation beta counter (type 1409; LKB-Wallac, Turku, Finland).
In competition assays using BSP or TA as substrates, the respective cells were incubated in the presence of rifampicin (Riedel-DeHaën, Seelze, Germany), BSP (Sigma-Aldrich), and Gd-EOB-DTPA (Primovist; Bayer-Schering, Berlin, Germany), respectively. The cellular uptake of Gd-EOB-DTPA (0.125, 0.25, 0.5, 0.75, and 1.0 mM) was measured after incubation for 10 min. After adding 0.2% sodium dodecyl sulfate, Gd-EOB-DTPA was quantified in cell lysates using liquid chromatography-tandem mass spectrometry. Protein concentration was determined using the Bradford (1976) method according to the manufacturer's instructions (Bio-Rad, Munich, Germany). Cell vitality in presence of Gd-EOB-DTPA was confirmed using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay (Mosmann, 1983). All experiments were performed in triplicates. In all uptake experiments, we ensured that the cellular influx of the substrates was still in the linear range during the incubation time of 5 to 10 min.
Quantitative Assay for Gd-EOB-DTPA.
The analyte was quantified in cell homogenates after protein precipitation with acetonitrile and enrichment with ethyl acetate using liquid chromatography-tandem mass spectrometry. The analytical system was equipped with the Agilent 1100 series HPLC system (Agilent Technologies, Waldbronn, Germany) and the MDS Sciex API 4000 mass spectrometer (Applied Biosystems, Darmstadt, Germany). In brief, 200 μl of homogenate was mixed with 25 μl of internal standard (1 μg/ml amoxicillin) and 25 μl of sodium azide (0.2%). Protein was precipitated with 400 μl of ice-cold acetonitrile. The supernatant was added to 1.0 ml of ethyl acetate, which was intensively vortexed for 30 s and centrifuged at 11.000 rpm for 3 min. Five microliters of the watery residue was injected into the chromatographic system. Chromatography (column, Atlantis HILIC Silica, 2.1 × 100 mm; Waters, Milford, MA) was performed using a steep-elution gradient with acetonitrile and ammonium acetate (5 mM, pH 3.8) as mobile phases at a flow rate of 200 μl/min. The tandem mass spectrometry analysis was done in the negative multiple reaction monitoring mode by monitoring the following m/z transitions: Gd-EOB-DTPA 681.3/635.2 and amoxicillin 363.8/222.7. The method was validated between 1.25 and 500 ng/ml, and it was shown to be of adequate specificity, precision (2.6–11% of mean values), and accuracy (−3.8–9.6% of the nominal values). The concentrations of the quality control samples were 2.5, 25, and 500 ng/ml.
Biometry.
All data are presented as mean ± S.D. Transporter kinetics was evaluated using Origin 5.0 (OriginLab Corp., Northampton, MA). The IC50 values were calculated by fitting the data to a sigmoid dose-response regression curve. Michaelis-Menten constant (Km) and maximum transport velocity (Vmax) were calculated using the following equation: y = Vmax × x/(Km + x). The uptake characteristics were corrected for nonspecific uptake into control cells.
Results and Discussion
Our in vitro experiments to evaluate affinity of Gd-EOB-DTPA to hepatic OATP transporters were performed with adequately characterized cell models. Western blot analysis revealed strong bands with the expected molecular masses of 84 kDa for OATP1B1, 120 kDa for OATP1B3, 85 kDa for OATP2B1, and 45 kDa for NTCP. Immunofluorescence analysis of HEK-OATPs and HEK-NTCP cells localized all transporters to the cell surface (Fig. 1). In uptake assays, HEK-OATP and HEK-NTCP showed a time-dependent and saturable accumulation of BSP and TA, respectively, which was in the linear range after incubation for 5 min. The Km values of 0.69 ± 0.16 μM for OATP1B1, 2.86 ± 1.56 μM for OATP1B3, 5.41 ± 3.76 μM for OATP2B1, and 22.11 ± 1.95 μM for NTCP were in line with comparable data published previously. Cui et al. (2001) measured a Km value of 0.14 μM for the OATP1B1-mediated uptake of BSP in stable transfected HEK293 cells, which were stimulated with 10 mM sodium butyrate. For the OATP1B3-mediated uptake of BSP, König et al. (2000b) obtained a Km value of 3.3 μM. For OATP2B1, Kopplow et al. (2005) observed a Km value of 3.4 μM in double-transfected MDCK-OATP2B1/ABCC2 and Kullak-Ublick et al. (2001) obtained 0.7 μM in complementary RNA (cRNA)-injected X. laevis oocytes. To our knowledge, there are no data on BSP uptake in OATP2B1-transfected HEK293 cells. For the NTCP-mediated uptake of TA, the respective Km values were 6.2 μM in cRNA-injected X. laevis oocytes (Hagenbuch and Meier, 1994), 2.1 μM in transiently transfected HEK293 cells (Leslie et al., 2007), and 7.9 μM in stable transfected HeLa cells (Kim et al., 1999). So far, there are no data on the TA uptake in stable transfected HEK293 cells.
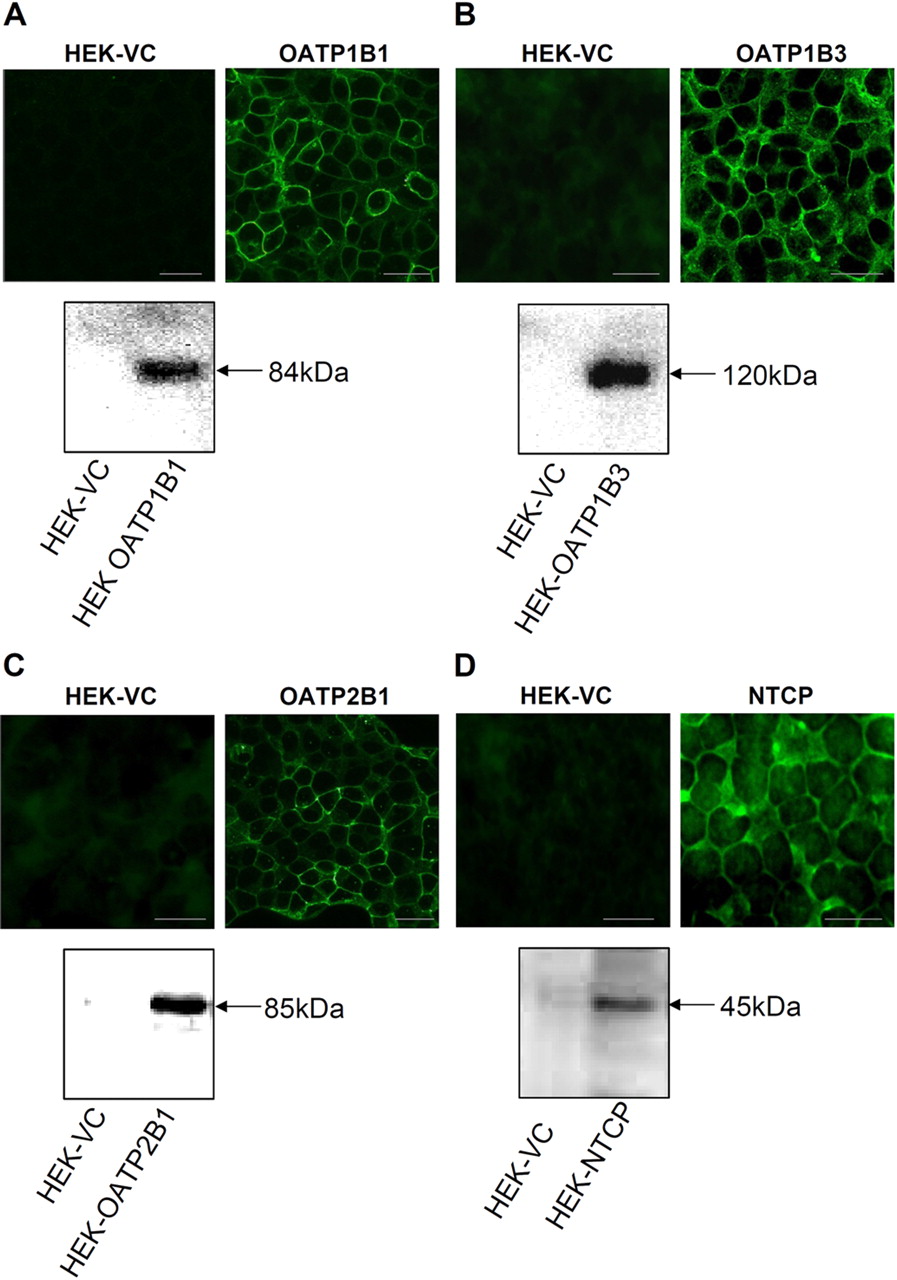
Immunofluorescence microscopy and Western blot analyses of stable transfected cell lines HEK-OATP1B1 (A), HEK-OATP1B3 (B), HEK-OATP2B1 (C), and HEK-NTCP (D). VC, vector-transfected cells. Scale bars, 20 μm.
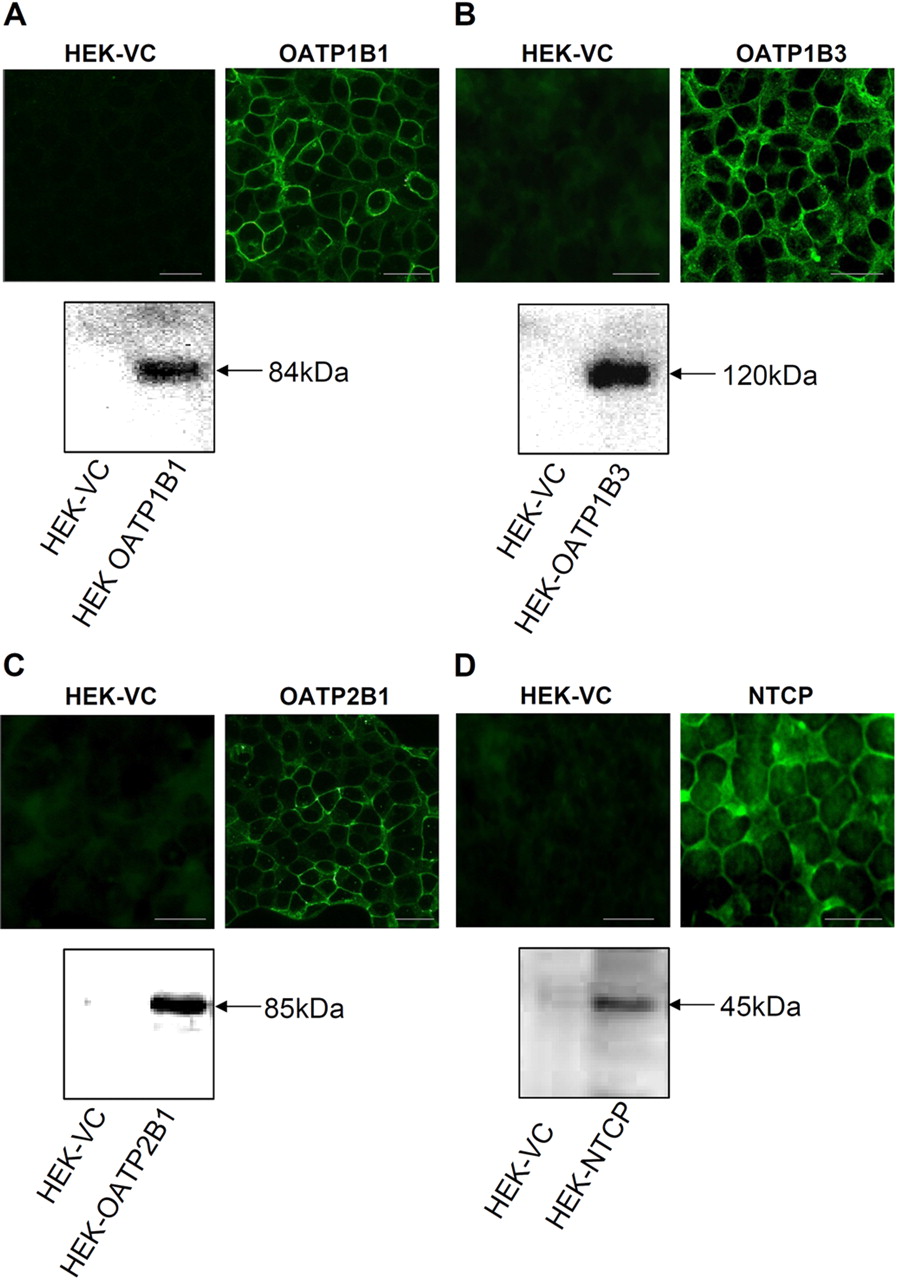
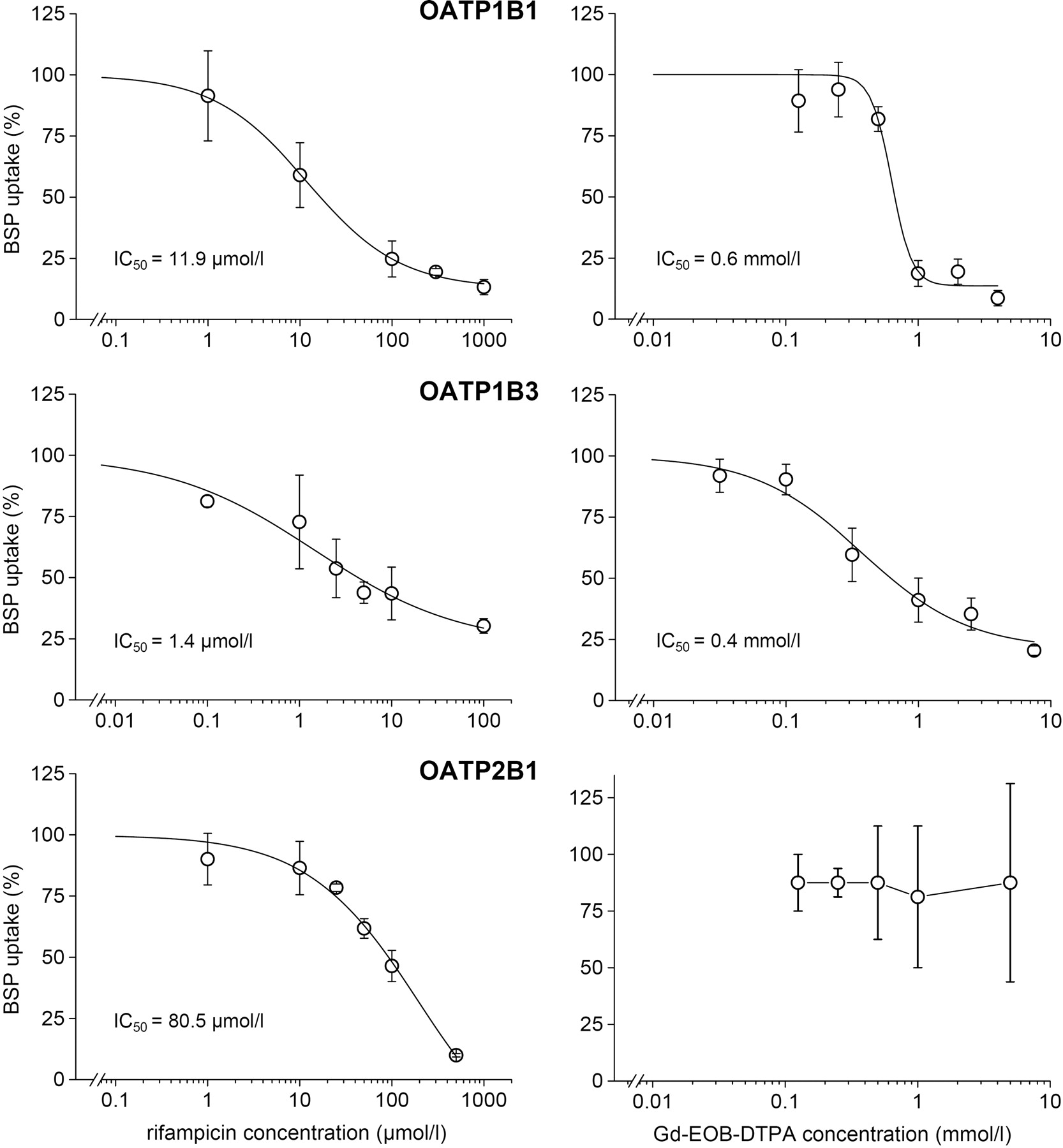
The BSP uptake by OATP1B1, OATP1B3, and OATP2B1 was efficiently inhibited by rifampicin with highest affinity to OATP1B3 (IC50 = 1.4 μM), medium affinity to OATP1B1 (IC50 = 11.9 μM), and lowest affinity to OATP2B1 (IC50 = 80.5 μM) (Fig. 2). In comparison, other authors measured IC50 values of 5 μM (OATP1B1), 17 μM (OATP1B3), and >100 μmol (OATP2B1) using cRNA-injected X. laevis oocytes (Vavricka et al., 2002). As expected, BSP competed with the transport of TA by NTCP (IC50 = 8.8 μM) (Fig. 3). This value was close to the IC50 of 7.3 μmol observed in stable transfected HeLa cells (Kim et al., 1999).

Competition of rifampicin (left) and Gd-EOB-DTPA (right) with the uptake of BSP into HEK cells transfected with OATP1B1, OATP1B3, and OATP2B1. BSP concentrations were 0.05 μM for HEK-OATP1B1 and 1 μM for HEK-OATP1B3 and HEK-OATP2B1. Means ± S.D. are given for n = 3 experiments.

Competition of BSP (left) and Gd-EOB-DTPA (right) with the uptake of 10 μM TA into HEK cells transfected with human NTCP. Means ± S.D. are given for n = 3 experiments.
Gd-EOB-DTPA was found to be an inhibitor of the BSP uptake by OATP1B1 and OATP1B3 with IC50 values of 0.6 and 0.4 mM, respectively. The contrast agent lacked cytotoxic effects up to concentrations of 10 mM, which was confirmed using the MTT assay. Gd-EOB-DTPA did not compete with the BSP uptake by the ubiquitous OATP2B1 (Fig. 2). TA uptake by NTCP cells was also not influenced by Gd-EOB-DTPA (Fig. 3).
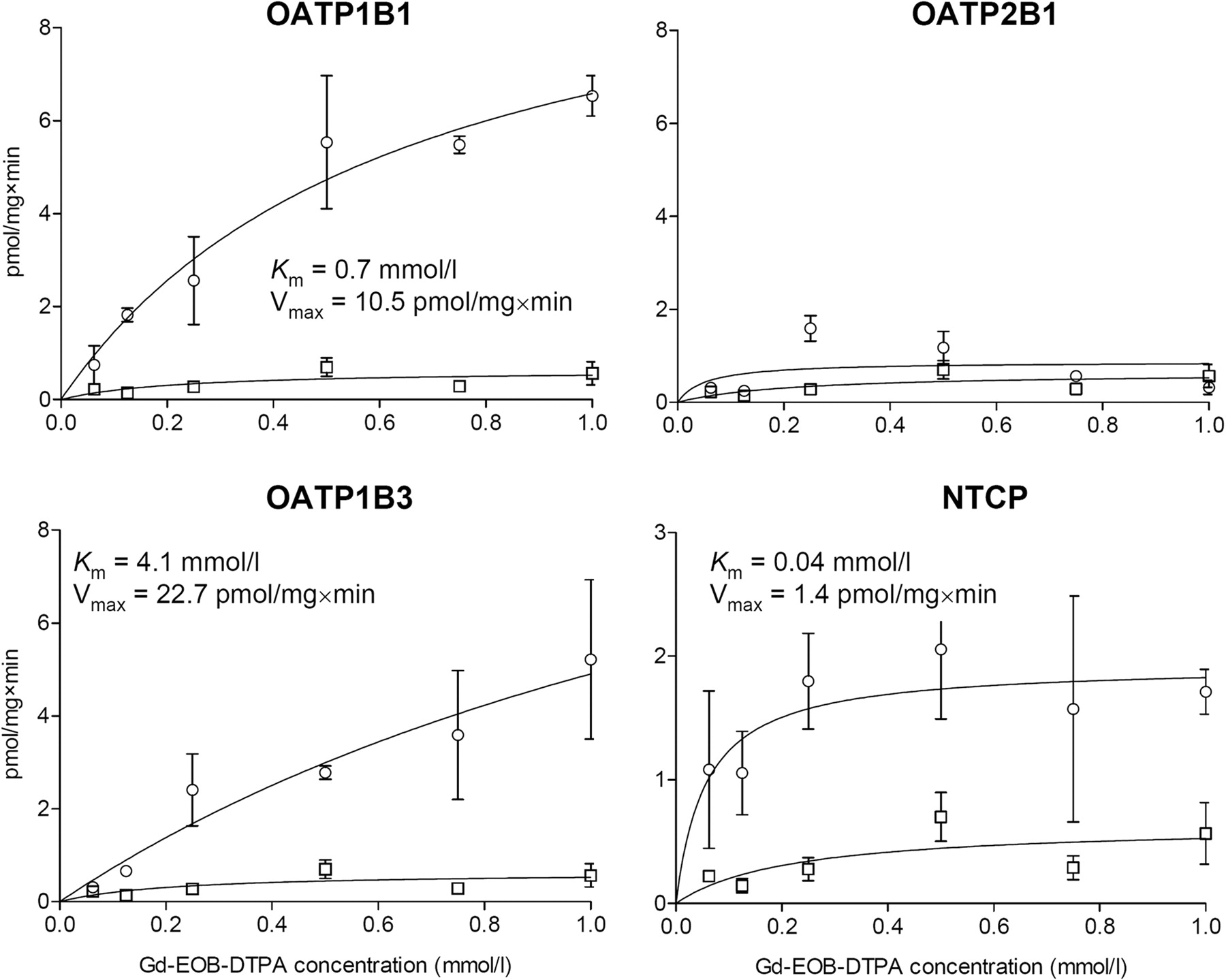
Gd-EOB-DTPA was extracted by the liver-specific OATP1B1 (Km = 0.7 mM, Vmax = 10.5 pmol/mg × min), OATP1B3 (Km = 4.1 mM, Vmax = 22.7 pmol/mg × min), and NTCP (Km = 0.04 mM, Vmax = 1.4 pmol/mg × min) (Fig. 4). NTCP was a carrier with higher affinity but lower capacity to Gd-EOB-DTPA than OATP1B1 and OATP1B3. The ubiquitous OATP2B1 was not involved in the cellular uptake of Gd-EOB-DTPA. Gd-EOB-DTPA-mediated uptake by HEK-OATP1B1-, HEK-OATP1B3-, and HEK-NTCP-transfected cells showed a time-dependent increase that was in the linear range after incubation for 10 min.

Uptake of Gd-EOB-DTPA in HEK cells transfected with human OATP1B1, OATP1B3, OATP2B1, and NTCP (open circles), and vector-transfected control cells (open squares) after incubation for 10 min. Means ± S.D. are given for n = 3 experiments.
We can conclude from the in vitro data that Gd-EOB-DTPA is substrate and inhibitor of the human liver-specific uptake transporters OATP1B1 and OATP1B3 and substrate of the human NTCP. The intrinsic uptake clearance of these transporters was in the same order of magnitude (OATP1B1, 0.015 ml/min × mg; OATP1B3, 0.006 ml/min × mg; NTCP, 0.035 ml/min × mg). The Km values are in the order of plasma concentrations that were measured in peripheral venous blood of healthy subjects after intravenous bolus injection or short-time infusion of 0.2 to 0.35 mmol/kg (0.3–0.6 mM) (our unpublished data; Schuhmann-Giampieri et al., 1997). Therefore, we hypothesize that OATP1B1, OATP1B3, and NTCP may contribute to overall hepatic enhancement of Gd-EOB-DTPA in humans after administration of standard doses for MRI. This conclusion is in line with the already mentioned preclinical data on uptake of Gd-EOB-DTPA in X. laevis oocytes injected with rat Oatp1a1 cRNA and the uptake characteristics in rat liver (van Montfoort et al., 1999; Kato et al., 2002). Furthermore, it was recently shown that enhancement of Gd-EOB-DTPA in hepatocellular carcinoma cells is correlated to expression of OATP1B3 (Narita et al., 2009). Finally, liver enhancement with Gd-EOB-DTPA is closely correlated to the disappearance rate of indocyanine green (Motosugi et al., 2009), which is in turn predicted by the expression levels of OATP1B1 and OATP1B3 but not OATP2B1 (Aoki et al., 2009). In patients with chronic liver disease, the signal intensity in Gd-EOB-DTPA-enhanced MRI, for which indocyanine green retention rate is a significant predictor (Takao et al., 2009), may therefore result from the net uptake rate via OATPs and NTCP. Therefore, the rate of Gd-EOB-DTPA enhancement in the liver may be a surrogate for the function of hepatic anion uptake transporters to be quantified by MRI. In addition, liver enhancement may be controlled by changes in hepatic secretion via ABCB4 and ABCC2, which are efflux carriers for indocyanine green and Gd-EOB-DTPA, respectively (Muhler et al., 1993; Huang and Vore, 2001).
In conclusion, Gd-EOB-DTPA is a substrate of the liver-specific uptake transporters OATP1B1, OATP1B3, and NTCP. Further evaluation is needed to confirm their meaning in overall enhancement in liver magnetic resonance imaging with Gd-EOB-DTPA.
Acknowledgments.
We are grateful to Dr. Jörg König (Department of Experimental and Clinical Pharmacology and Toxicology, Erlangen, Germany) for providing antibodies against OATP1B1 and OATP1B3 and Professor Bruno Stieger (Department of Clinical Pharmacology and Toxicology, Zürich, Switzerland) for providing the coding sequence of NTCP and anti-NTCP antibodies. We also express gratitude to Gitta Schumacher, Sabine Bade, Edita Kaliwe, and Danilo Wegner for excellent technical assistance.
Footnotes
This work was supported by the German Federal Ministry of Education and Research [Grants BMBF InnoProfile 03IP612, BMBF 0314107].
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
doi:10.1124/dmd.110.032862.
-
ABBREVIATIONS:
- MRI
- magnetic resonance imaging
- Gd-EOB-DTPA
- gadolinium-ethoxybenzyl-diethylenetriamine pentaacetic acid
- NTCP
- Na+/taurocholate cotransporting polypeptide
- OATP
- organic anion-transporting polypeptide
- BSP
- bromosulfophthalein
- HEK
- human embryonic kidney
- TA
- taurocholic acid
- MTT
- 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
- MDCK
- Madin-Darby canine kidney
- cRNA
- complementary RNA.
- Received February 16, 2010.
- Accepted April 16, 2010.
- Copyright © 2010 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}