


Abstract
Drug-induced hepatotoxicity, which is a rare but serious adverse reaction to a large number of pharmaceutical drugs, is sometimes associated with reactive metabolites produced by drug-metabolizing enzymes. In the present study, we constructed a cell-based system to evaluate the cytotoxicity of reactive metabolites produced by CYP3A4 using human hepatoma cells infected with an adenovirus vector expressing human CYP3A4 (AdCYP3A4). When seven hepatoma cell lines (HepG2, Hep3B, HLE, HLF, Huh6, Huh7, and Fa2N4 cells) were infected with AdCYP3A4, HepG2 cells showed the highest CYP3A4 protein expression and testosterone 6β-hydroxylase activity (670 pmol · min−1 · mg−1). With the use of AdCYP3A4-infected HepG2 cells, the cytotoxicities of 23 drugs were evaluated by the 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium monosodium salt assay, and the cell viability when treated with 11 drugs (amiodarone, desipramine, felbamate, isoniazid, labetalol, leflunomide, nefazodone, nitrofurantoin, tacrine, terbinafine, and tolcapone) was significantly decreased. Moreover, the transfection of siRNA for nuclear factor erythroid 2-related factor 2 (Nrf2) to decrease the cellular expression level of Nrf2 exacerbated the cytotoxicity of some drugs (troglitazone, flutamide, acetaminophen, clozapine, terbinafine, and desipramine), suggesting that the genes regulated by Nrf2 are associated with the detoxification of the cytotoxicities mediated by CYP3A4. We constructed a highly sensitive cell-based system to detect the drug-induced cytotoxicity mediated by CYP3A4. This system would be beneficial in preclinical screening in drug development and increase our understanding of the drug-induced cytotoxicity associated with CYP3A4.
Introduction
Drug-induced hepatotoxicity is a rare but serious adverse reaction to a large number of pharmaceutical drugs (Boelsterli and Lim, 2007). One of the mechanisms suggested for the drug-induced hepatotoxicity is associated with reactive metabolites produced by drug-metabolizing enzymes (Guengerich and MacDonald, 2007). For example, if the reactive metabolites covalently bind to intracellular proteins, cellular dysfunctions are apparently produced, resulting in the loss of ionic gradients, a decline in ATP levels, actin disruption, cell swelling, and cell rupture (Beaune et al., 1987; Yun et al., 1993). Although drug-induced hepatotoxicity can be evaluated using laboratory animal models, species differences in drug-metabolizing enzymes or other factors between humans and laboratory animals have made the prediction of drug-induced cytotoxicity difficult. Several studies used human hepatocytes to evaluate the drug-induced cytotoxicity mediated by drug-metabolizing enzymes (Li et al., 1999; Gómez-Lechón et al., 2003), but this approach is often not applicable because of the poor availability of human liver, the high cost, the significant variability among human hepatocyte preparations, and, in particular, the unstable enzyme activities of human hepatocytes.
To date, several systems for evaluating drug-induced cytotoxicity mediated by drug-metabolizing enzymes have been developed. Yoshitomi et al. (2001) used HepG2 cells stably expressing human cytochrome P450 (P450) enzymes, and the viabilities of the cells expressing CYP1A2, CYP2E1, and CYP3A4 were decreased in an acetaminophen concentration-dependent manner. Vignati et al. (2005) demonstrated, using HepG2 cells transiently transfected with CYP3A4, that reactive metabolites of various hepatotoxic drugs such as albendazole, flutamide, and troglitazone were produced by CYP3A4. Our previous study showed that benzodiazepines such as flunitrazepam and nimetazepam were metabolically activated by CYP3A4 by coincubation with HepG2 cells and CYP3A4 Supersomes (Mizuno et al., 2009). Thus, several in vitro studies demonstrated that P450 enzymes, especially CYP3A4, are involved in the cytotoxicities of various kinds of drugs. However, transfection of P450 expression plasmids into cells generally could not achieve high expression of P450 enzymes. In addition, many studies used cells with GSH levels decreased by treatment with l-buthionine sulfoximine to easily detect the cytotoxicity. Indeed, the GSH-conjugating process is responsible for the detoxification of reactive metabolites, but we considered that cells with decreased nuclear factor erythroid 2-related factor 2 (Nrf2) levels would also be useful to sensitively detect the cytotoxicity, because Nrf2 is a transcription factor that acts as a main regulator for the up-regulation of a group of genes coding antioxidant proteins and phase II drug-metabolizing enzymes, such as NADPH-quinone oxidoreductase 1, heme oxygenase-1, and glutamate-cysteine ligase, catalytic subunit (GCLC) and modifier subunit (GCLM) (Thimmulappa et al., 2002; Balogun et al., 2003; Nioi et al., 2003). In fact, it has been reported that Nrf2 knockout mice had increased susceptibility to acetaminophen (Chan et al., 2001).
In a previous study, we established an adenovirus that could overexpress human CYP3A4 (AdCYP3A4) (Hosomi et al., 2010). In this study, we developed a highly sensitive in vitro cell-based assay system using AdCYP3A4 and investigated whether CYP3A4 is involved in the cytotoxicity of 23 drugs that are known to cause hepatotoxicity in humans. In addition, we investigated whether treatment with siNrf2 exacerbates the drug-induced cytotoxicity mediated by CYP3A4.
Materials and Methods
Chemicals and Reagents.
Acetaminophen, allopurinol, clozapine, corticosterone, cyclizine, dantrolene sodium, desipramine, disulfiram, erythromycin, felbamate, 3-hydroxyacetanilide (3-HAA), maprotiline, nefazodone, nilutamide, rosiglitazone, sulindac, tacrine, terbinafine, testosterone, tolcapone, troglitazone, and zafirlukast were obtained from Wako Pure Chemicals (Osaka, Japan). Flutamide, 6β-hydroxytestosterone, isoniazid, labetalol, and nitrofurantoin were obtained from Sigma-Aldrich (St. Louis, MO). Amiodarone and leflunomide were obtained from LKT Labs (St. Paul, MN) and Enzo Life Sciences, Inc. (Farmingdale, NY), respectively. ReverTra Ace (Moloney murine leukemia virus reverse transcriptase RNaseH Minus) was from Toyobo (Tokyo, Japan). The Adenovirus Expression Vector Kit (Dual Version), RNAiso, random hexamer, and SYBR Premix Ex Taq were obtained from Takara (Shiga, Japan). The QuickTiter Adenovirus Titer Immunoassay Kit was from Cell Biolabs (Tokyo, Japan). Stealth Select RNAi for Nrf2 (siNrf2) (accession number NM_006164) and Stealth RNAi Negative Control Medium GC Duplex #2 (siScramble), Lipofectamine RNAiMAX Reagent, and Lipofectamine 2000 were obtained from Invitrogen (Carlsbad, CA). Dulbecco's modified Eagle's medium was from Nissui Pharmaceutical (Tokyo, Japan). MFE support media (serum-free) was from MultiCell Technologies (Lincoln, RI). All primers were commercially synthesized at Hokkaido System Sciences (Sapporo, Japan). Other chemicals were of analytical or the highest grade commercially available.
Cell Culture.
The 293 and HepG2 cells were obtained from American Type Culture Collection (Manassas, VA). The HLE and Huh7 cells were obtained from Japan Collection of Research Biosources (Tokyo, Japan) and RIKEN BioResource Center (Ibaraki, Japan), respectively. The Fa2N4 cells were obtained from MultiCell Technologies. The Hep3B, Huh6, and HLF cells were kindly provided by Dr. Shuichi Kaneko (Kanazawa University, Kanazawa, Japan). The 293, HepG2, Hep3B, HLE, HLF, Huh6, and Huh7 cells were maintained in Dulbecco's modified Eagle's medium containing 10% fetal bovine serum (Invitrogen), 3% glutamine, 16% sodium bicarbonate, and 0.1 mM nonessential amino acids (Invitrogen) in a 5% CO2 atmosphere at 37°C. Fa2N4 cells were grown on collagen-coated flasks and maintained in MFE support media containing 10% fetal bovine serum. When adenovirus was used to infect the cell lines, a medium containing 5% fetal bovine serum was used.
siRNA Treatment.
HepG2 cells were transfected with siNrf2 or siScramble by Lipofectamine RNAiMAX Reagent. According to the manufacturer's protocol, RNAi duplex-Lipofectamine RNAiMAX complexes were prepared and added to each well before the HepG2 cells were seeded (2.0 × 104 cells/well). The concentrations of siNrf2 and siScramble were 10 nM.
Total RNA Preparation from Hepatoma Cells and Real-Time RT-PCR.
Total RNA from hepatoma cells was isolated using RNAiso. The RT procedure was described previously (Nakajima et al., 2006). Human Nrf2 was quantified by real-time RT-PCR using the Smart Cycler (Cepheid, Sunnyvale, CA). The sequences of sense and antisense primers were 5′-CAACACACGGTCCACAGC-3′ and 5′-CAATATTAAGACACTGTAACTC-3′, respectively. A 1-μl portion of the reverse-transcribed mixture was added to a PCR mixture containing 10 pmol of sense and antisense primers and 12.5 μl of SYBR Premix Ex Taq solution in a final volume of 25 μl. The PCR conditions were as follows: after an initial denaturation at 95°C for 30 s, the amplification was performed by denaturation at 94°C for 6 s and annealing and extension at 64°C for 20 s for 45 cycles. Human glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA was also quantified as described previously (Tsuchiya et al., 2004). The expression level of Nrf2 mRNA was normalized with the GAPDH mRNA level.
Nrf2 Protein Level.
The Nrf2 protein level was measured as described previously (Nakamura et al., 2008) with slight modifications. Cell lysates (30 μg) were separated on 7.5% polyacrylamide gel electrophoresis and electrotransferred onto Immobilon-P polyvinylidene difluoride membrane (Millipore Corporation, Billerica, MA). The membranes were probed with polyclonal rabbit anti-Nrf2 antibody (H-300) (Santa Cruz Biotechnology, Inc., Santa Cruz, CA), polyclonal rabbit anti-human GAPDH polyclonal antibodies (Sekisui Medical, Tokyo, Japan), and anti-rabbit IgG-conjugated IRDye680, and an Odyssey infrared imaging system (LI-COR Biosciences, Lincoln, NE) were used for the detection. The relative expression level was quantified using ImageQuant TL Image Analysis software (GE Healthcare, Little Chalfont, Buckinghamshire, UK).
CYP3A4 Enzyme Activity.
AdCYP3A4 was constructed in our previous study (Hosomi et al., 2010). HepG2 cells (3 × 105 cells/well) were seeded in 12-well plates. After a 24-h incubation, the cells were infected with AdCYP3A4 for 48 h at a multiplicity of infection (MOI) of 10 (MOI 10). Then, after a 1-h incubation with 100 μM testosterone, the concentration of 6β-hydroxytestosterone, a metabolite of testosterone by CYP3A4, in the medium was measured as described previously (Hosomi et al., 2010).
Cytotoxicity Assay.
HepG2 cells (2.0 × 104 cells/well) were seeded in 96-well plates and, if necessary, were treated with siNrf2 or siScramble as described above. After a 24-h incubation, the cells were infected with AdCYP3A4 or the recombinant adenovirus vector expressing green fluorescence protein (AdGFP) constructed in our previous study (Hosomi et al., 2010). The titers of AdCYP3A4 and AdGFP were 6.4 × 108 and 2.1 × 108 plaque forming units/ml, respectively. Forty-eight hours after infection at MOI 10, the cells were treated with various drugs for 24 h. After incubation with the drugs, cell viability was quantified by 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium monosodium salt (WST-8) and ATP assays according to the manufacturer's protocol. The WST-8 assay, which is a modified 3-(4,5-dimethylthiazol-2-yl)2,5-diphenyl tetrazolium bromide assay, was performed using Cell Counting Kit-8 (Wako Pure Chemicals). After incubation with the drugs, Cell Counting Kit-8 reagent was added, and absorbance of WST-8 formazan was measured at 405 nm. The ATP assay was performed using a CellTiter-Glo Luminescent Cell Viability Assay (Promega, Madison, WI). After incubation with the drugs, CellTiter-Glo Reagent was added, and the generation of luminescent signals was recorded by using a 1420 ARVO MX luminometer (PerkinElmer Life and Analytical Sciences-Wallac Oy, Turku, Finland).
Statistical Analyses.
Comparisons of two and several groups were made with a unpaired, two-tailed Student's t test and Dunnett's test, respectively. P < 0.05 was considered statistically significant.
Results
CYP3A4 Enzyme Activity in Various Hepatoma Cell Lines Infected with AdCYP3A4.
To determine which hepatoma cell line is suitable for the evaluation of drug-induced cytotoxicity mediated by CYP3A4, seven hepatoma cell lines (HepG2, Hep3B, HLE, HLF, Huh6, Huh7, and Fa2N4 cells) were infected with AdCYP3A4 at MOI 10, and the testosterone 6β-hydroxylase activity was measured at a concentration of 100 μM (Fig. 1). Among these cell lines, HepG2 cells showed the highest enzyme activity (670 pmol · min−1 · mg protein−1). Huh7 cells also showed moderate enzyme activity (233 pmol · min−1 · mg protein−1), but the other cells showed activity at less than 100 pmol · min−1 · mg protein−1. From this result, HepG2 cells were used in the subsequent studies.

CYP3A4 enzyme activity in various cell lines infected with AdCYP3A4. Cells were infected with AdCYP3A4 at MOI 10 for 48 h, and the testosterone 6β-hydroxylase activity was measured as described under Materials and Methods. Data are the means of two independent experiments.
Cytotoxic Effects of CYP3A4 on Cell Viability of HepG2 Cells.
To investigate whether CYP3A4 is associated with the cytotoxicity of 23 drugs (acetaminophen, allopurinol, amiodarone, clozapine, cyclizine, dantrolene, desipramine, disulfiram, erythromycin, felbamate, flutamide, isoniazid, labetalol, leflunomide, maprotiline, nefazodone, nitrofurantoin, sulindac, tacrine, terbinafine, tolcapone, troglitazone, and zafirlukast), HepG2 cells infected with AdCYP3A4 at MOI 10 for 48 h were treated with the drugs for 24 h (Fig. 2). As a negative control, AdGFP was infected at MOI 10. The cytotoxicity assay was performed at the concentrations less than the concentration that showed the typical decreased cell viability by the parent drugs. However, the range of concentrations was at least 30 times the clinically efficacious concentration or 100 μM according to a previous report (O'Brien et al., 2006). Cytotoxicity was evaluated by the WST-8 assay (Fig. 2), which showed that the viabilities of the cells infected with AdCYP3A4 were significantly decreased compared with that of those infected with AdGFP when treated with amiodarone (10 and 25 μM), desipramine (10 and 25 μM), disulfiram (25–100 μM), isoniazid (50 and 100 μM), leflunomide (100 μM), nefazodone (25 μM), tacrine (100 μM), terbinafine (10–50 μM), or tolcapone (20 and 50 μM). However, the other drugs did not affect the cell viability by the overexpression of CYP3A4 in the HepG2 cells.

Cytotoxic effects of CYP3A4 expression on cell viability of HepG2 cells treated with various drugs. HepG2 cells were infected with AdCYP3A4 (●) or AdGFP (○) at MOI 10 for 48 h and treated with 23 drugs for 24 h. Cell viability was measured by the WST-8 assay. Each point represents the mean ± S.D. (n = 3). *, P < 0.05; **, P < 0.01; ***, P < 0.001, compared with the AdGFP group at each concentration of the drug.
Cytotoxic Effects of Knockdown of Nrf2 Expression on Cell Viability of HepG2 Cells.
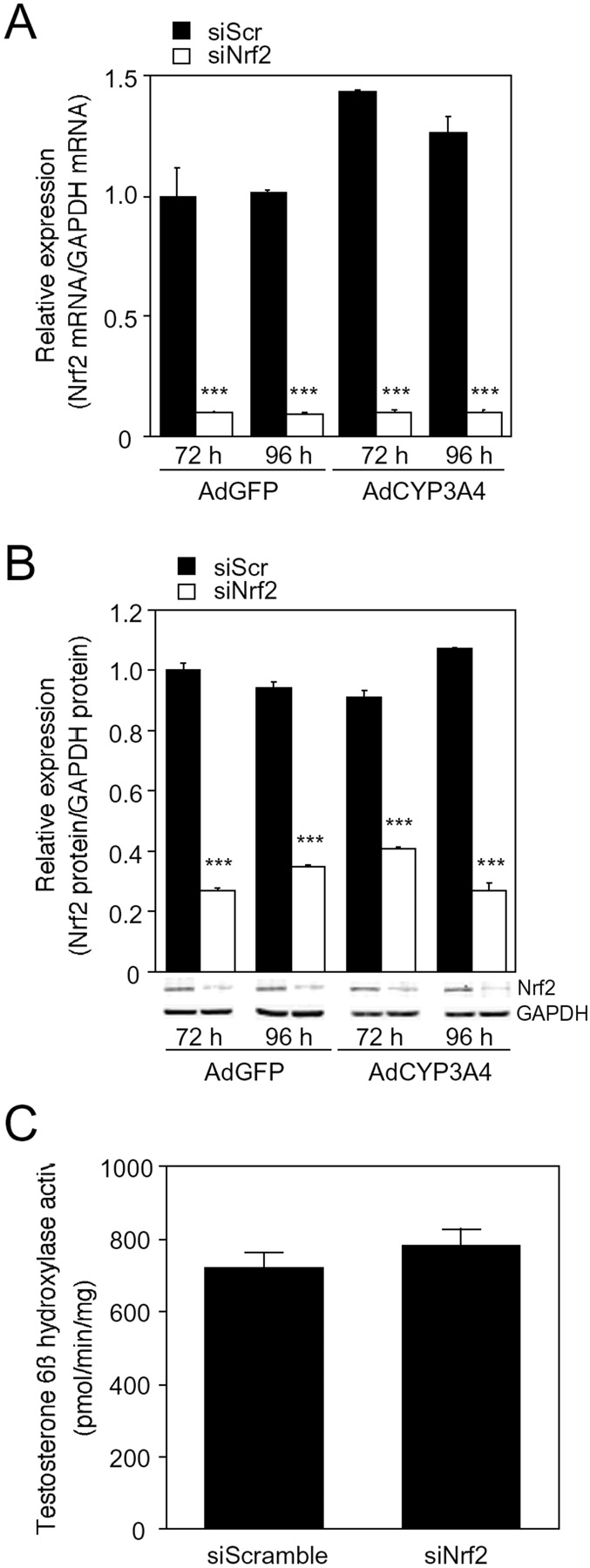
Nrf2 is a transcription factor that acts as an important regulator of the up-regulation of a group of genes coding antioxidant proteins and phase II drug-metabolizing enzymes, such as NADPH-quinone oxidoreductase, heme oxidase-1, and GCLC. Therefore, decreased Nrf2 levels in the cells may enhance the drug-induced cytotoxicity caused by CYP3A4. In this study, the drug-induced cytotoxicity mediated by CYP3A4 was investigated using AdCYP3A4-infected HepG2 cells transfected with siNrf2. The expression level of Nrf2 mRNA in HepG2 cells treated with siNrf2 for 72 and 96 h was lower than 90% of that of cells treated with siScramble (Fig. 3A). The expression level of Nrf2 protein in HepG2 cells treated with siNrf2 was also 55 to 75% lower than that of cells treated with siScramble (Fig. 3B). The effect of siNrf2 on the expression levels of Nrf2 mRNA and protein was not different between the treatments with AdGFP and AdCYP3A4. In addition, the HepG2 cell lysate treated with siNrf2 showed testosterone 6β-hydroxylase activity (781 ± 47 pmol · min−1 · mg protein−1) similar to that of cell lysate treated with siScramble (720 ± 40 pmol · min−1 · mg protein−1) (Fig. 3C). Therefore, it was confirmed that the decreased level of Nrf2 expression did not affect the testosterone 6β-hydroxylase activity.

Expression level of Nrf2 mRNA (A) and protein (B) and testosterone 6β-hydroxylase activity (C) in AdCYP3A4-infected HepG2 cells with siNrf2. After HepG2 cells were transfected with siNrf2 or siScramble at 10 nM followed by the infection with AdCYP3A4 or AdGFP at MOI 10 for 48 h, the expression level of Nrf2 mRNA and protein and testosterone 6β-hydroxylase activity were measured as described under Materials and Methods. Each column represents the means ± S.D. (n = 3). ***, P < 0.001 compared with AdCYP3A4-infected cells with siScramble.
With the use of HepG2 cells, the drug-induced cytotoxicity was investigated (Fig. 4). As a negative control against siNrf2, siScramble was transfected to HepG2 cells. Four drugs (troglitazone, flutamide, acetaminophen, and clozapine) that did not show cytotoxicity in the presence of CYP3A4 in the previous analysis (Fig. 2) and two drugs (desipramine and terbinafine) that showed cytotoxicity (Fig. 2) were investigated. In the WST-8 assay, the viabilities of AdCYP3A4-infected cells with siNrf2 were significantly decreased compared with those with siScramble for treatment with troglitazone (25 and 50 μM), flutamide (10–50 μM), acetaminophen (2.5–20 mM), clozapine (25–50 μM), desipramine (10 and 25 μM), or terbinafine (10 and 25 μM) (Fig. 4A). The ATP assay showed results similar to those of the WST-8 assay, although the magnitude of the difference in cell viability between cells with siNrf2 and those with siScramble was smaller (Fig. 4B). Thus, these results suggested that AdCYP3A4-infected HepG2 cells with decreased levels of Nrf2 were useful to evaluate the drug-induced cytotoxicity mediated by CYP3A4.

Effects of the decreased level of Nrf2 on cell viability of CYP3A4-expressing HepG2 cells treated with the drugs. HepG2 cells were transfected with siNrf2 (●) or siScramble (○) at 10 nM for 24 h followed by infection with AdCYP3A4 at MOI 10 for 48 h. After cells were treated with six drugs for 24 h, WST-8 (A) and ATP (B) assays were performed for evaluation of the cell viability. Each point represents the mean ± S.D. (n = 3). *, P < 0.05; **, P < 0.01; ***, P < 0.001, compared with AdCYP3A4-infected cells with siScramble in each concentration of drug.
To investigate whether any drugs may affect the viability of cells with decreased levels of Nrf2, we examined the changes in cell viability by treatment with rosiglitazone, nilutamide, and 3-HAA, which have been reported to show a lower risk of hepatotoxicity but have structures similar to troglitazone, flutamide, and acetaminophen (Supplemental Fig. 1). However, rosiglitazone, nilutamide, and 3-HAA did not show decreased cell viability even when the Nrf2 level in cells was decreased. This result was consistent with the fact that these drugs are not likely to be associated with CYP3A4-mediated hepatotoxicity.
Discussion
Drug-induced hepatotoxicity is a rare but serious adverse reaction to a large number of pharmaceutical drugs (Boelsterli and Lim, 2007). Because drug-induced hepatotoxicity is sometimes associated with reactive metabolites produced by drug-metabolizing enzymes (Guengerich and MacDonald, 2007), it would be ideal to be able to predict the hepatotoxicity mediated by these enzymes using an in vitro system. In the present study, we constructed a highly sensitive cell-based system to detect the cytotoxicity of drugs using HepG2 cells expressing CYP3A4, which is a predominant P450 isoform in human liver that is responsible for more than 50% of drug metabolism (Guengerich, 2008).
In general, infection of cells with a recombinant adenovirus can lead to high expression of specific genes of interest. In our previous study, we introduced the overexpression of CYP3A4 protein in rat hepatoma cells, H4IIE cells, by the infection of AdCYP3A4 (Hosomi et al., 2010). Likewise, in this study, seven human hepatoma cell lines were infected with AdCYP3A4, and the testosterone 6β-hydroxylase activity was measured. Among them, HepG2 cells showed the highest enzyme activity (Fig. 1). This result might reflect the fact that HepG2 cells express high endogenous levels of NADPH-P450 reductase and cytochrome b5 (Aoyama et al., 1990) in addition to the high efficiency of infection. The testosterone 6β-hydroxylase activity of AdCYP3A4-infected HepG2 cells (670 pmol · min−1 · mg protein−1 at 100 μM testosterone) was more than 2.5-fold higher than the Vmax values of the activity in human hepatocytes reported by Donato et al. (1995) and Gómez-Lechón et al. (2001) (177 ± 98 and 253 ± 110 pmol · min−1 · mg protein−1, respectively). Given the large interindividual variability in CYP3A4 protein expression (a 32-fold variability) (Westlind-Johnsson et al., 2003), the cells used in this study would mimic human hepatocytes that exhibit the highest CYP3A4 enzyme activity.
In this study, we investigated the CYP3A4-mediated cytotoxicity of 23 drugs that are known to cause hepatotoxicity in humans (Xu et al., 2008). The viability of cells infected with AdCYP3A4 was significantly decreased by treatment with amiodarone, desipramine, disulfiram, isoniazid, leflunomide, nefazodone, nitrofurantoin, tacrine, terbinafine, and tolcapone compared with those infected with AdGFP. It has already been reported that the hepatotoxicity of some drugs is associated with the metabolic activation mediated by CYP3A4. The major metabolite of amiodarone, desethylamiodarone, was reported to cause cytotoxicity in HepG2 cells and rat hepatocytes at lower concentrations than the parent drug amiodarone (McCarthy et al., 2004). Desethylamiodarone is produced mainly by CYP3A4 and by CYP2C8 in humans (Ohyama et al., 2000). Erythromycin-associated hepatic injury is usually cholestatic and may mimic obstructive jaundice (Klatskin, 1975). Although CYP3A4 is involved in erythromycin metabolism and the substrate to which erythromycin binds has been identified (Watkins, 1992), it has no demonstrated relevance to erythromycin metabolism-associated hepatic injury. Nefazodone is metabolized to hydroxynefazodone, followed by the formation of quinone-imine and 2-chloro-1,4-benzoquinone, which would be associated with the hepatotoxicity, in human liver microsomes. These reactions are mediated by CYP3A4 (Kalgutkar et al., 2005). Terbinafine is mainly metabolized to N-demethylterbinafine, 7,7-dimethylhept-2-ene-4-ynal (TBF-A), and several hydroxy forms in human liver. Among them, TBF-A, which is produced by CYP3A4, was reported to cause terbinafine-induced hepatotoxicity (Iverson and Uetrecht, 2001). However, no association of CYP3A4 with desipramine-, disulfiram-, isoniazid-, leflunomide-, nitrofurantoin-, and tolcapone-induced hepatotoxicity has been reported, although most of these drugs appeared to be metabolically activated by P450 enzymes. For example, CYP1A2 is reported to be associated with the formation of iminoquinone from tacrine and tolcapone (Balson et al., 1995; Smith et al., 2003). Furthermore, CYP2E1 is considered to be associated with isoniazid-induced hepatotoxicity by metabolizing acetylhydrazine, one of the metabolites of isoniazid, to N-acetylhydrazine (Huang et al., 2003). Further studies on the involvement of CYP3A4 in the metabolic activation of these drugs are needed. Although pharmacological drug interactions were reported in cyclizine, a metabolic activation pathway would not be involved.
Nitroaromatic drugs such as tacrine and nitrofurantoin have been associated with liver injury (Boelsterli et al., 2006). In the reductive pathways from nitro to the fully reduced amine catalyzed mainly by cytochrome P450 reductase, several reactive metabolites including nitroso and N-hydroxylamine derivatives could be produced. The reductive metabolite of tacrine is produced by cytochrome P450 reductase in HepG2 cells, and the enhanced reactive oxygen species production results in cytotoxic effects (Osseni et al., 1999). Cytochrome P450 reductase is also involved in nitrofurantoin-induced redox cycling and cytotoxicity (Wang et al., 2008). There is no report suggesting the involvement of CYP3A4 in the metabolic activation of tacrine and nitrofurantoin. From these lines of evidence, the positive cytotoxic effects of tacrine and nitrofurantoin in Fig. 2 would be due to the cytochrome P450 reductase activity, which is highly expressed in tumor cells compared with hepatocytes. Flutamide is known to induce severe hepatic dysfunction. Ohbuchi et al. (2009) suggested that CYP3A4 catalyzed the N-oxidation of the amino metabolite of flutamide, which had hepatotoxic effects. Dantrolene was also implicated in the generation of reactive oxygen species via cytochrome P450 reductase. However, we previously reported that CYP3A4-mediated cytotoxicity was not involved in the effects of the nitroaromatic drugs, flutamide and dantrolene, which we investigated using CYP3A4-expressing rat BRL3A cells (Yoshikawa et al., 2009), supporting our present data in Fig. 2. Although the cytochrome P450 reductase-mediated redox reactions were suggested in flutamide- and dantrolene-induced hepatotoxicities, the underlying mechanism for metabolic activation is likely to be different for different drugs, producing different cytotoxic outcomes, an issue that needs to be clarified.
Felbamate induces the activity of CYP3A4, which accounts for the in vivo drug interactions in human (Glue et al., 1997). It was reported that felbamate heteroactivates the CYP3A4-mediated pathway (Egnell et al., 2003). However, these kinds of adverse effects were not detected in the present assay system, because only the cytotoxic effect by the CYP3A4-mediated metabolic activation of drug could be assessed. In addition, hepatotoxicity and hypersensitivity syndromes are associated with allopurinol (Al-Kawas et al., 1981; Arellano and Sacristán, 1993). Although the involvement of the CYP3A4-mediated reaction was still not clear, this kind of hepatotoxicity was also not detected in the present system.
A cytotoxic effect of metabolic activation of drug by CYP3A4 (Fig. 2, ●) was clearly demonstrated in a dose-dependent manner at 10 and 25 μM desipramine and nitrofurantoin. The cytotoxicity of parent drugs was demonstrated to be potent at 50 μM desipramine and nitrofurantoin and amiodarone (Fig. 2, ●). Thus, the effect of metabolic activation by CYP3A4 could not be detected at 50 μM concentrations of the drugs (Fig. 2), indicating that it should be considered the cytotoxicity of active metabolite(s) as well as that of the parent drugs at the various concentrations of drugs used in this in vitro assay system.
Nrf2 is the main regulator of the up-regulation of genes associated with detoxification reactions and is known to be constitutively and ubiquitously expressed in several tissues and cell lines. In the absence of cellular stress, Nrf2 is localized in the cytosol by binding with Kelch-like erythroid cell-derived protein with CNC homology-associated protein 1. When cells are exposed to oxidative stress, the newly synthesized Nrf2 accumulates in the nucleus (Kobayashi et al., 2006). However, it has been reported that Nrf2 is localized in the nucleus in HepG2 cells under nonstressed conditions (Nguyen et al., 2005). In the present study, HepG2 cells with decreased Nrf2 expression were used to detect the drug-induced cytotoxicity more sensitively. The Nrf2 mRNA level in HepG2 cells could be decreased to less than 10% by siNrf2 (Fig. 3A). Given that the Nrf2 mRNA level in HepG2 was 1.6-fold higher than that in human hepatocytes (data not shown), the HepG2 cells used in this study had approximately 20% of the Nrf2 mRNA expression as that in human hepatocytes.
The present study was conducted as shown in Fig. 4 with four drugs (troglitazone, flutamide, acetaminophen, and clozapine) that did not show cytotoxicity in AdCYP3A4-infected HepG2 cells without siNrf2 and two drugs that did show cytotoxicity (desipramine and terbinafine). The viabilities of cells transfected with siNrf2 were significantly decreased by treatment with the drugs compared with those transfected with siScramble (Fig. 4). These results suggest that the genes regulated by Nrf2 play important roles in the detoxification of the drug-induced cytotoxicities activated by CYP3A4. It has been reported that troglitazone is metabolized to quinone-type forms, which may be the reactive metabolites produced by CYP3A4 and CYP2C8 (Yamazaki et al., 1999). Although it has been reported that a postulated reactive metabolite of clozapine was produced by myeloperoxidase, leading to depletion of the cellular GSH level (Williams et al., 1997), there is no information about the association of CYP3A4 with the cytotoxicity of clozapine. Flutamide is reported be oxidized into reactive metabolites that covalently bind to microsomal proteins metabolized by CYP3A4 and CYP1A2 (Berson et al., 1993; Fau et al., 1994). In addition, acetaminophen is metabolized to N-acetyl-p-benzoquinone imine, a postulated ultimate reactive metabolite, by CYP2E1 and CYP3A4 in humans. These reactive metabolites could be trapped with GSH in a process that is the major detoxification pathway. Because GCLC, GCLM, and glutathione peroxidase, which are important factors for maintaining the cellular GSH level, are regulated by Nrf2 (Thimmulappa et al., 2002), the decreased viability of AdCYP3A4-infected cells with siNrf2 by these drugs may be due to the decreased level of cellular GSH. The reactive metabolite of terbinafine, TBF-A, is known be trapped with GSH (Iverson and Uetrecht, 2001). The viability of AdCYP3A4-infected HepG2 cells was decreased by treatment with terbinafine even without siNrf2 transfection. This result was different from those by the treatment with troglitazone, flutamide, and acetaminophen. In addition, information about the association of CYP3A4/Nrf2 with desipramine-induced hepatotoxicity is limited, although epoxide formation and nitrosoalkane species were suggested to be related to the toxicity.
The WST-8 assay is a modified 3-(4,5-dimethylthiazol-2-yl)2,5-diphenyl tetrazolium bromide assay, a conventional cytotoxicity assay, that uses a highly water-soluble disulfonated tetrazolium salt as a chromogenic indicator. The WST-8 assay was used to measure the mitochondrial NADH enzymes as an indicator of cell viability. The ATP assay was used to measure the cellular concentration of ATP, which is produced in mitochondria. Thus, in both assays, the cell viability was evaluated by measuring the mitochondrial activity, but the sensitivity of the ATP assay is lower than that of the WST-8 assay (Fig. 4), and the result should therefore be carefully evaluated.
Many researchers have used cells transfected with P450 expression plasmids to investigate the drug-induced cytotoxicity mediated by P450s, but this system generally could not induce high expression of P450 enzymes. However, our established adenovirus-using system induced the overexpression of CYP3A4 in HepG2 cells, resulting in more than 2.5-fold higher enzyme activity than that of human hepatocytes. In addition, the transfection of siNrf2 to HepG2 cells could decrease to approximately 20% the Nrf2 expression level in human hepatocytes. Thus, the combination of AdCYP3A4 infection with the transfection of siNrf2 would enable sensitive detection of the drug-induced cytotoxicity mediated by CYP3A4.
In conclusion, we constructed a highly sensitive cell-based system to detect the drug-induced cytotoxicity mediated by CYP3A4. Furthermore, we found that a decrease in the level of Nrf2 could increase the sensitivity of detection for drug-induced cytotoxicity. It is generally known that very complicated mechanisms are involved in drug-induced liver injury and the basic mechanism is likely to be different for different drugs. Therefore, this in vitro assay system is not likely to predict in vivo drug-induced liver injury. However, this system would be beneficial in the preclinical screening in drug development and increase our understanding of the drug-induced cytotoxicity associated with CYP3A4.
Authorship Contributions
Participated in research design: Hosomi, Fukami, Nakajima, and Yokoi.
Conducted experiments: Hosomi and Iwamura.
Performed data analysis: Hosomi, Fukami, and Iwamura.
Wrote or contributed to the writing of the manuscript: Hosomi, Fukami, and Yokoi.
Acknowledgments
We thank Brent Bell for reviewing the manuscript.
Footnotes
This work was supported by Health and Labor Science Research Grants from the Ministry of Health, Labor, and Welfare of Japan [Grant H20-BIO-G001].
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
doi:10.1124/dmd.110.037077.
↵
 The online version of this article (available at http://dmd.aspetjournals.org) contains supplemental material.
The online version of this article (available at http://dmd.aspetjournals.org) contains supplemental material.-
ABBREVIATIONS:
- P450
- cytochrome P450
- Nrf2
- nuclear factor erythroid 2-related factor 2
- GCLC
- glutamate-cysteine ligase, catalytic subunit
- GCLM
- glutamate-cysteine ligase, modifier subunit
- 3-HAA
- 3-hydroxyacetanilide
- RT
- reverse transcription
- PCR
- polymerase chain reaction
- GAPDH
- glyceraldehyde-3-phosphate dehydrogenase
- MOI
- multiplicity of infection
- WST-8
- 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium monosodium salt
- TBF-A
- 7,7-dimethylhept-2-ene-4-ynal.

- Received November 4, 2010.
- Accepted May 3, 2011.
- Copyright © 2011 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}