


Abstract
Ionic liquids (ILs) are a class of salts that are expected to be used as a new source of solvents and for many other applications. Our previous studies revealed that selected ILs, structurally related organic cations, are eliminated exclusively in urine as the parent compound, partially mediated by renal transporters. This study investigated the inhibitory effects of N-butylpyridinium chloride (NBuPy-Cl) and structurally related ILs on organic cation transporters (OCTs) and multidrug and toxic extrusion transporters (MATEs) in vitro and in vivo. After Chinese hamster ovary cells expressing rat (r) OCT1, rOCT2, human (h) OCT2, hMATE1, or hMATE2-K were constructed, the ability of NBuPy-Cl, 1-methyl-3-butylimidazolium chloride (Bmim-Cl), N-butyl-N-methylpyrrolidinium chloride (BmPy-Cl), and alkyl substituted pyridinium ILs to inhibit these transporters was determined in vitro. NBuPy-Cl (0, 0.5, or 2 mg/kg per hour) was also infused into rats to assess its effect on the pharmacokinetics of metformin, a substrate of OCTs and MATEs. NBuPy-Cl, Bmim-Cl, and BmPy-Cl displayed strong inhibitory effects on these transporters (IC50 = 0.2–8.5 μM). In addition, the inhibitory effects of alkyl-substituted pyridinium ILs on OCTs increased dramatically as the length of the alkyl chain increased. The IC50 values were 0.1, 3.8, 14, and 671 μM (hexyl-, butyl-, and ethyl-pyridinium and pyridinium chloride) for rOCT2-mediated metformin transport. Similar structurally related inhibitory kinetics were also observed for rOCT1 and hOCT2. The in vivo coadministration study revealed that NBuPy-Cl reduced the renal clearance of metformin in rats. These results demonstrate that ILs compete with other substrates of OCTs and MATEs and could alter the in vivo pharmacokinetics of such substrates.
Introduction
Ionic liquids (ILs) are a growing class of industrial chemicals that are being increasingly investigated for a variety of applications (Plechkova and Seddon, 2008). They are usually composed of organic cations and various inorganic or organic anions. They have melting points at or less than 100°C. Because of the chemical diversity of the cations/anions, the number of available ILs is almost unlimited (Baker et al., 2005). ILs with specific melting points, viscosities, densities, or ionic conductivities can be formulated for special applications (Welton, 1999). Therefore, ILs are expected to be used widely in analytical methods, engineering processes, consumer products, and biomedical applications (Plechkova and Seddon, 2008). Because of their extremely low vapor pressure, their capacity to pollute the air is minimal. Thus, a primary application of these compounds is to replace the classic volatile organic solvents (Rogers and Seddon, 2003).
Although the physical/chemical characteristics of ILs may minimize the risk of atmospheric contamination, other sources of environmental exposure are of concern. For example, Couling et al. (2005) suggested that the increased use of ILs on a large scale in industry could result in water pollution. Exposures from occupational and consumer products are also likely to occur. Some studies have already investigated the toxic effects caused by pyridinium- and imidazolium-based ILs. On a molecular level, such ILs altered the function of some biological enzymes, such as oxidoreductase and mushroom tyrosinase (Pinto et al., 2008; Yang et al., 2009). Zhao et al. (2007) have summarized the toxicities of these two groups of ILs to various organisms, such as water algae, bacteria, fungi, and mammalian cell lines. To some organisms, ILs are much more toxic than the conventional organic solvents (Ranke et al., 2004).
ILs based on imidazolium, pyridinium, phosphonium, and ammonium have been tested on Cyclotella meneghiniana, Selenastrum capricornutum, Daphnia magna, Pseudokirchneriella subcapitata, and zebrafish (Latala et al., 2005; Cho et al., 2007; Pretti et al., 2009; Yu et al., 2009). It was found that shorter alkyl-substituted chains demonstrated lower toxicity compared with the cations with a longer alkyl substituent. This structure-activity relationship (SAR) was also observed for biological enzymes, such as acetylcholine esterase (Stock et al., 2004; Stasiewicz et al., 2008). Lactic acid production by Lactobacillus, an acid-producing bacterium, was decreased as the alkyl chain length on the imidazolium cations increased (Matsumoto et al., 2004). Ranke et al. (2007) compared the cation lipophilicity and the cytotoxicity of 74 ILs on IPC-81 leukemia cells and suggested that the length of the alkyl chain played a dominant role in their cytotoxicity.
Few studies have investigated the disposition and toxicity of ILs in mammals. Because N-butylpyridinium chloride (NBuPy-Cl), 1-methyl-3-butylimidazolium chloride (Bmim-Cl), and N-butyl-N-methylpyrrolidinium (BmPy-Cl) are starting materials for many other ILs, they were selected by the National Toxicology Program for investigation of their disposition, metabolism, and toxicity. In previous studies, we reported that these three compounds are absorbed (40–70%) from the rodent gastrointestinal tract and then are widely distributed in rats (Sipes et al., 2008; Cheng et al., 2009; Knudsen et al., 2009). For the three ILs, the portion of the dose that became systemically available was eliminated exclusively in the urine as the parent compound. Renal elimination was facilitated, in part, by tubular secretion, presumably mediated by organic cation transporter 2 (OCT2).
In this study, we examined the in vitro inhibitory effects of NBuPy-Cl, Bmim-Cl, and BmPy-Cl on rat OCT1/2 and human OCT2 and also determined whether these ILs inhibited human MATE1/2-K. How the length of the alkyl chain affected the degree of inhibition was further investigated on OCTs. The ILs used were pyridine hydrochloride (Py-Cl), 1-ethylpyridinium chloride (EtPy-Cl), and 1-hexylpyridinium chloride (HePy-Cl), derivatives of NBuPy-Cl (Fig. 1). In addition, in vivo studies were conducted to determine whether NBuPy-Cl can influence the pharmacokinetics of metformin, a known substrate of OCTs and MATEs.

Chemical structures of NBuPy-Cl and other structurally related ionic liquids.
Materials and Methods
Chemicals and Materials.
[3H]Tetraethylammonium trifluoroacetate (TEA) (54 Ci/mmol) was synthesized by GE Healthcare (Chalfont St. Giles, Buckinghamshire, UK). [14C]Metformin (112 mCi/mmol) was received from Moravek Biochemicals (Brea, CA). [3H]1-Methyl-4-phenylpyridinium (MPP) (80 Ci/mmol) was synthesized by the Department of Chemistry and Biochemistry, University of Arizona (Tucson, AZ). Metformin (98% purity) was purchased from Sigma-Aldrich (St. Louis, MO). NBuPy-Cl, Bmim-Cl, and BmPy-Cl (all ≥98% purity) (Fig. 1) were obtained from Merck KGaA (Darmstadt, Germany). Py-Cl, EtPy-Cl, and HePy-Cl (all ≥98% purity) were purchased from Acros Organics (Geel, Belgium). Ketamine/xylazine and pentobarbital sodium salt were purchased from Sigma-Aldrich. Solvable and Pico-Flour scintillation cocktail solution were obtained from PerkinElmer Life and Analytical Sciences (Waltham, MA). Fetal bovine serum, penicillin/streptomycin, phleomycin (Zeocin), and hygromycin B were obtained from Invitrogen (Carlsbad, CA). Kaighn's modification (F12K) medium and other chemicals (the highest quality available) were purchased from Sigma-Aldrich.
In Vitro Studies.
Cell culture and transfection.
Chinese hamster ovary cells containing a single integrated Flp recombination target site (CHO Flp-In) were acquired from Invitrogen and were used for stable expression of the rat ortholog of OCT1 (rOCT1) and OCT2 (rOCT2) and the human ortholog of OCT2 (hOCT2), MATE1 (hMATE1), and MATE2-K (hMATE2-K). Transporter cDNAs were inserted into the pcDNA5/FRT/V5-His-TOPO plasmid vector (Invitrogen) following the manufacturer's instructions. The sequences were confirmed by the DNA Sequencing Facility at the University of Arizona (Tucson, AZ). CHO cells were maintained in F12K medium with 10% fetal bovine serum, penicillin (100 units/ml), streptomycin (100 μg/ml), and phleomycin (100 μg/ml) at 37°C in a humidified atmosphere with 5% CO2, and 5 × 106 cells in 400 μl of media were electroporated (BTX ECM 630) at 260 V (time constant of ∼25 ms) with 10 μg of salmon sperm DNA (Invitrogen), 18 μg of pOG44, and 2 μg of the plasmid vector containing transporter cDNA. The cells were then transferred to T75 cell culture flasks. After a 24-h incubation, medium containing hygromycin (300 μg/ml), instead of phleomycin, was applied as selection pressure for 2 weeks.
Transport studies.
The transport of [3H]TEA (12 nM) and [14C]metformin (9 μM) was first characterized over time in CHO_hOCT2, CHO_rOCT1, and CHO_rOCT2 cells along with [3H]MPP (15 nM) in CHO_hMATE1 and CHO_hMATE2-K cells. On the basis of the time-dependent transport curve, the inhibitory effects of ILs toward OCTs were determined during the linear phase of uptake (30 s for [3H]TEA uptake and 60 s for [14C]metformin uptake by hOCT2; 120 s for [3H]TEA uptake by rOCT1, [14C]metformin uptake by rOCT2, and [3H]MPP uptake by hMATE1 and hMATE2-K).
To determine the inhibitory effects of ILs, the transport of [3H]TEA (12 nM) by rOCT1 and hOCT2 and of [14C]metformin (9 μM) by rOCT2 and hOCT2 was measured in the presence of increasing concentrations of NBuPy-Cl, Bmim-Cl, BmPy-Cl, Py-Cl, EtPy-Cl, or HePy-Cl. Various concentrations of NBuPy-Cl were used to investigate its inhibitory effects on hMATE1- or hMATE2-K-mediated [3H]MPP transport. The transport assay was performed as described previously (Cheng et al., 2009). In general, cells were cultured in 12-well plates. Once reaching confluence, they were incubated in 0.4 ml of Waymouth's buffer containing [14C]metformin, [3H]TEA, or [3H]MPP and one of the ILs for a predetermined time as described above. After solubilization of the cells with 0.4 ml of NaOH (0.5 M) with 1% SDS and neutralization with 0.2 ml of HCl (1 M), the cell lysate solution (0.5 ml) was counted using a liquid scintillation counter (LSC).
Animal Studies.
Animal surgeries.
Male Fischer-344 rats (300–350 g) were purchased from Harlan (Indianapolis, IN). The rats were housed in the University of Arizona Animal Care Facility (accredited by the Association for Assessment and Accreditation for Laboratory Animal Care) with controlled temperature (25°C), humidity (40–60%), and light/dark cycle (12 h). After 7 to 10 days of acclimation, 1 ml/kg ketamine/xylazine solution was administered (intraperitoneally) to induce anesthesia. Once the animals were totally anesthetized, the jugular vein and carotid artery were cannulated with PE-50 tubing (i.d. 0.58 mm and o.d. 0.965 mm; BD, Franklin Lakes, NJ). The surgical openings were covered with water-saturated gauze sponges. After surgery, pentobarbital (65 μg/kg) was administered subcutaneously to maintain anesthesia. The rats were then placed in a ventral position on a heating pad maintained at 37°C. A syringe infusion pump (Harvard Apparatus, Holliston, MA) was used for the infusion of NBuPy-Cl. All the protocols were approved by the Institutional Animal Care and Use Committee at the University of Arizona.
Dosing selection and sample collection.
It was observed previously that NBuPy-Cl (5 mg/kg) did not affect the GFR in rats after a single intravenous dose (Cheng et al., 2009). However, when this dose (5 mg/kg per hour, 2 ml/kg per hour) was infused over 4 h (total dose 20 mg/kg), the clearance of the metformin (assessed over 3 h) was reduced to <1% of that observed in the control (saline group). Therefore, two lower doses (2 mg/kg per hour, 0.5 mg/kg per hour) of NBuPy-Cl were used in the studies described here. They provided total doses of 8 and 2 mg/kg, respectively.
After surgery, the jugular vein was infused with saline (2 ml/kg per hour) or NBuPy-Cl (0.5 or 2 mg/kg per hour, 2 ml/kg per hour) over 4 h. A bolus dose of [14C]metformin (5 mg/kg, 50 μCi/kg) was administered through the jugular vein after 1 h of perfusion. Blood samples (300 μl) were then collected at 7.5, 15, 30, 45, 60, 90, 120, 150, and 180 min from the carotid artery. An equal volume of saline was administered to replace the blood that was withdrawn. At the end of the experiment (3 h after metformin dosing), animals were euthanized by CO2 inhalation. Selected tissues (adipose, heart, kidney, liver, lung, muscle, spleen, and testes) were collected.
Sample Analysis.
Blood samples were centrifuged at 750g for 10 min. Aliquots of the plasma samples (two 50-μl samples) were mixed with 15 ml of the Pico-Flour scintillation cocktail solution, and samples were counted using a LSC. Tissues samples were analyzed as described by Sipes et al. (2008). In brief, the samples were solubilized with Solvable, quenched with H2O2 (30%), and counted using a LSC.
Data Analysis.
For the in vitro transport studies, estimates of the IC50 values were determined by the modified Michaelis-Menten equation (Malo and Berteloot, 1991; Groves et al., 1994):
 J is the transport rate of radiolabeled compound at the concentration of [T*], Jmapp is the Jmax for [*T] transport times the ratio of Ki/Kt, IC50 is equal to Ki(1 + [T*/Kt]), [I] is the concentration of unlabeled inhibitor, and C is a constant representing the component of total uptake that is not saturable over the concentration range tested.
J is the transport rate of radiolabeled compound at the concentration of [T*], Jmapp is the Jmax for [*T] transport times the ratio of Ki/Kt, IC50 is equal to Ki(1 + [T*/Kt]), [I] is the concentration of unlabeled inhibitor, and C is a constant representing the component of total uptake that is not saturable over the concentration range tested.
The application of this equation, given its structure, carries the tacit assumption that the inhibitory interactions observed are competitive in nature and reflect binding of substrate and inhibitor at a common binding site. Although the substrates/inhibitors studied here have all been shown to be transported by OCTs, inhibitory interactions between OCT substrates have, in some cases, been shown to reflect a “mixed-type” inhibitory profile, presumably reflecting both competition for a common binding region within the transporter and longer-range allosteric interactions (e.g., Koepsell et al., 2007). Thus, we refer to the kinetic constants calculated through the above equation as “IC50” values and make no claim as to their precise mechanistic basis.
The pharmacokinetics of metformin after intravenous administration in rats was defined by WinNonlin software (version 6.1; Pharsight, Mountain View, CA) using a noncompartment model. All individual data points are presented as means ± S.E.M. The data were analyzed statistically by Student's t test or one-way analysis of variance with the Newman-Keuls post-test using GraphPad Prism 4 (GraphPad Software Inc., San Diego, CA). A level of p < 0.05 was considered significant.
Results
Functional Expression of OCTs and MATEs in CHO Cells.
The accumulation of 12 nM [3H]TEA by CHO cells stably expressing OCTs was time-dependent, i.e., nearly linear for at least 60 s and approaching steady state by 10 min (Fig. 2A). In all three cell lines, the presence of 5 mM unlabeled TEA reduced the 10-min accumulation of [3H]TEA by 90% or more. The uptake of [3H]TEA displayed kinetic characteristics of carrier-mediated transport minimally influenced by surface binding (Suhre et al., 2005). The profiles of hOCT2- and rOCT2-mediated [14C]metformin transport were similar to that of [3H]TEA, but the transport rate of [14C]metformin by rOCT1 was too low to conduct an inhibition study (data not shown). Therefore, uptake of [3H]TEA was used to determine the inhibitory effects of ILs on rOCT1.

Characterization of the function of OCTs and MATEs in transfected CHO cells. A, [3H]TEA uptake by rOCT1 (■), rOCT2 (▴), or hOCT2 (●) over time in the absence (solid symbols) or presence (open symbols) of 5 mM TEA. B, [3H]MPP uptake by hMATE1 (■) or hMATE2-K (●) over time in the absence (solid symbols) or presence (open symbols) of 1 mM MPP. n = 3; mean ± S.E.M.
The function of CHO cells expressing hMATE1 or hMATE2-K was also determined, and the results are shown in Fig. 2B. As with the OCTs, MATE1- or MATE2-K-mediated transport of radiolabeled substrate, in this case, [3H]MPP, was blocked more than 90% by unradiolabeled 1 mM MPP. These results demonstrate that OCTs and MATEs were expressed in the CHO cells and were functionally active.
Inhibitory Effect of NBuPy-Cl, Bmim-Cl, and BmPy-Cl on OCTs.
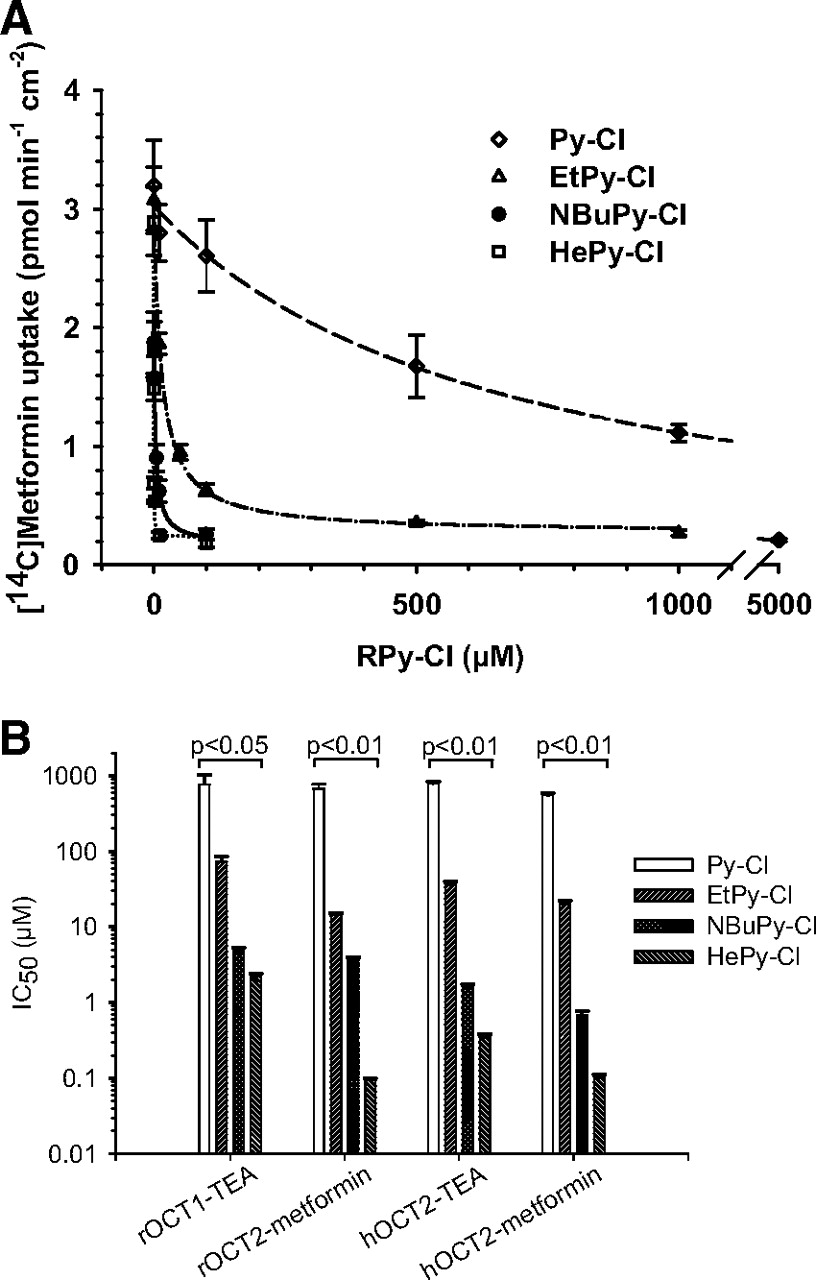
In the presence of NBuPy-Cl, the intracellular uptake of [3H]TEA by rOCT1 was inhibited in a concentration-dependent manner (Fig. 3A). Similar inhibitory kinetics of NBuPy-Cl on [14C]metformin transport were observed for rOCT2 and hOCT2 (Fig. 3, B and C). The IC50 values, which are the concentrations of inhibitors (ILs) that result in half-maximum transport of TEA or metformin by OCTs, are presented in Table 1. The three ILs were potent inhibitors of rOCT1/2 and hOCT2, with IC50 values ranging from 0.15 to 7.53 μM. As shown in Table 1, IC50 values were significantly lower for hOCT2 when metformin was used as the probe substrate.

Inhibitory effects of NBuPy-Cl on OCTs. Intracellular uptake of [3H]TEA by CHO_rOCT1 (A, 120 s) and [14C]metformin by CHO_rOCT2 (B, 120 s) and CHO_hOCT2 (C, 60 s) in the presence of increasing concentrations of NBuPy-Cl (n = 3–5; mean ± S.E.M.).
Inhibitory effects (IC50) of NBuPy-Cl and structurally related ILs on OCT-mediated TEA or metformin transport
Data are means ± S.E.M., n = 3 to 5.
Inhibition of OCTs by Alkyl-Substituted Pyridiniums.
To determine the relationship between the structure of ILs and their inhibitory effects (structure-activity relationship) on OCTs, pyridinium-based ILs with different alkyl chain lengths were investigated. Uptake of TEA or metformin by OCTs was measured in the presence of increasing concentrations of Py-Cl, EtPy-Cl, NBuPy-Cl, and HePy-Cl. As shown in Fig. 4A, all the pyridinium compounds tested inhibited rOCT2-mediated [14C]metformin transport. Py-Cl was the weakest inhibitor with an IC50 of 671 μM (Table 1). The inhibition curve shifted left gradually, and the IC50 values decreased approximately 10-fold for every 2 carbons added to the alkyl chain. HePy-Cl showed the strongest inhibitory activity (IC50 = 0.1 μM). The structurally related inhibitory effects of these pyridinium ILs on rOCT1 and hOCT2 were similar to those of rOCT2 (Fig. 4B). The IC50 values are presented in Table 1 for both probe substrates, TEA and metformin.

Influence of alkyl chain length on the inhibitory effects of pyridinium-based ILs on OCTs. A, intracellular uptake of [14C]metformin (120 s) by CHO_rOCT2 in the presence of increasing concentrations of RPy-Cl (RPy-Cl: Py-Cl, EtPy-Cl, NBuPy-Cl, and HePy-Cl; n = 3–5, mean ± S.E.M.). B, relationship between inhibitory effects (IC50) and alkyl chain length (number of carbons) of RPy-Cl (n = 3–5; mean ± S.E.M.). The statistical analysis compared the log(IC50) using one-way analysis of variance with the Newman-Keuls post-test.
Inhibitory Effects of NBuPy-Cl on hMATE1 and hMATE2-K.
The inhibitory effects of NBuPy-Cl on the apical membrane transporters, hMATE1 and hMATE2-K, were also determined. Figure 5 shows the inhibitory kinetic curves of NBuPy-Cl on intracellular uptake of [3H]MPP by CHO cells expressing hMATE1 or hMATE2-K. The IC50 values were 8.5 ± 2.6 and 1.6 ± 0.2 μM, respectively.
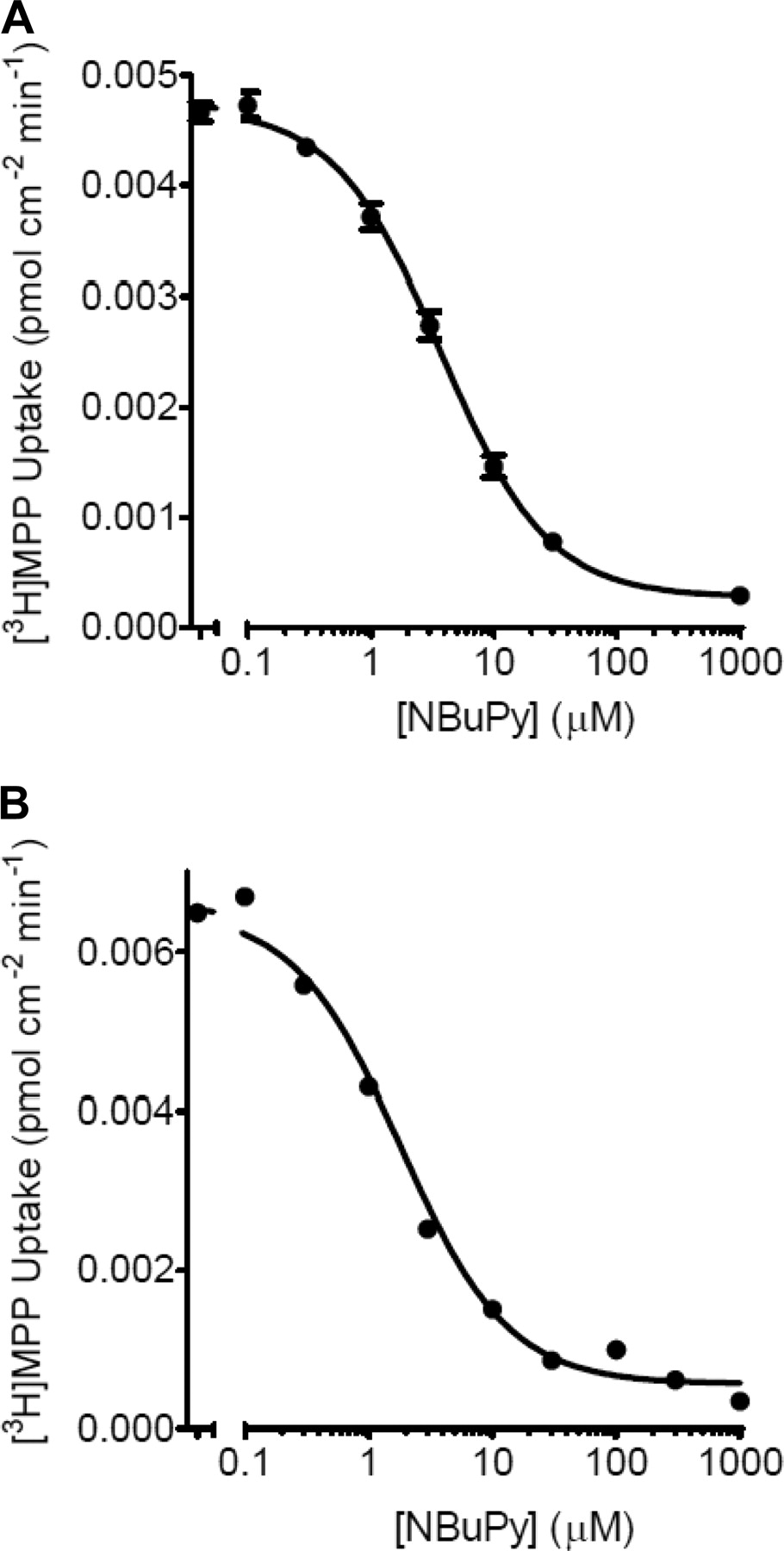
Inhibitory effects of NBuPy-Cl on hMATE1 and hMATE2-K. Intracellular uptake of [3H]MPP by CHO_hMATE1 (A) or CHO_hMATE2-K (B) in 2 min with the presence of increasing concentrations of NBuPy-Cl (n = 3–6; mean ± S.E.M.).
Intravenous Coadministration of NBuPy-Cl and Metformin.
Pharmacokinetic analysis using a noncompartment model revealed that the elimination half-life of [14C]metformin (5 mg/kg i.v.) from plasma of male F-344 rats was 2.3 h (Fig. 6; Table 2). The volume of distribution was 484 ml, and the systemic clearance was 3.4 ml/min. Because the liver metabolism/excretion of metformin is negligible, its systemic clearance reflects its renal clearance (Scheen, 1996). Infusion of NBuPy-Cl increased the plasma level of metformin in a dose-dependent manner (Fig. 6). Thus, the plasma area under the curve from 0 to 180 min for metformin was significantly increased because of the reduced renal clearance (Table 3). NBuPy-Cl had a minimal effect on the volume of distribution of metformin. This observation was further substantiated by the findings that tissue levels of metformin were not significantly elevated when they were harvested at the 3-h time point (Table 3). The small increases that were observed may reflect the presence of blood, which, in the NBuPy-Cl-infused rats, had elevated concentrations of metformin. The only tissue that showed a small increase in the tissue/plasma ratio, indicative of actual increases in tissue levels, was the kidney, but this value was not significant because of the large interanimal variations.

Effects of NBuPy-Cl on the pharmacokinetics of metformin. Plasma concentrations of [14C]metformin (n = 3–4; mean ± S.E.M.) after a single intravenous administration (5 mg/kg) to male F344 rats with saline (▴, 2 ml/h) or NBuPy-Cl (■, 0.5 mg/kg per hour; ●, 2 mg/kg per hour) infusion through the jugular vein.
Kinetic parameters of metformin in male F-344 rats after intravenous administration (5 mg/kg) with NBuPy-Cl or saline infusion
Data are means ± S.E.M.; n = 3 to 4. Predicted parameters were calculated using noncompartment model analysis.
Metformin concentration in selected tissues of F-344 rats after intravenous administration of metformin (5 mg/kg) to male F-344 rats infused with saline or NBuPy-Cl
Data are presented as means ± S.E.M. [tissue (g)/plasma (ml) metformin concentration ratio].
Discussion
OCTs are widely expressed on the basolateral membrane of proximal tubule cells, where they function to transport a wide array of organic cations from plasma to the intracellular space of these cells (Choi and Song, 2008). OCT2 is richly expressed in human and rat kidneys, but OCT1 is weakly detected in human kidneys (Okuda et al., 1996; Motohashi et al., 2002). On the apical membrane of these tubular cells, organic cations are most likely exported to the urinary lumen by MATEs (Nies et al., 2011). There are two mammalian MATEs, MATE1 and MATE2-K. Human kidneys express both homologs, whereas rodent kidneys express only MATE1 (Terada et al., 2006). In several studies, we observed that three ILs, NBuPy-Cl, Bmim-Cl, and BmPy-Cl, were excreted rapidly in the urine of rats as the parent compounds (Sipes et al., 2008; Cheng et al., 2009; Knudsen et al., 2009). Because the rate of clearance from plasma exceeded the glomerular filtration rate, it was proposed that these three ILs, which are organic cations, undergo transport-mediated secretion.
NBuPy-Cl and BmPy-Cl were significantly accumulated in CHO cells expressing hOCT2 after a 10-min incubation, compared with naive CHO cells (Cheng et al., 2009; Knudsen et al., 2009). The transport kinetics of these two ILs, as assessed by Kt values (Kt = 18 μM for NBuPy-Cl and 37 μM for BmPy-Cl), were similar to that of the model substrate of hOCT2, TEA (Kt = 40 μM). NBuPy-Cl and BmPy-Cl were also well transported in CHO cells expressing rOCT1 or rOCT2 (unpublished data). Results of subsequent experiments performed after the disposition studies on Bmim-Cl were published revealed that Bmim-Cl was also transported by hOCT2, rOCT1, and rOCT2 (data not presented). These in vitro results suggest that OCTs could contribute to the rapid renal elimination of these ILs.
The results presented in this article demonstrate that, in addition to serving as substrates, NBuPy-Cl, BmPy-Cl, and Bmim-Cl are also potent inhibitors of rOCT1/2 and hOCT2. The IC50 values for inhibition of hOCT2-mediated TEA transport by these three ILs ranged from 0.5 to 2.3 μM. These inhibitory values are similar to those observed for MPP (2 μM) and tetrapentylammonium (10 μM), compounds considered to be potent inhibitors of hOCT2 (Suhre et al., 2005). When metformin, the widely prescribed type 2 antidiabetic drug, was used as the probe substrate for hOCT2, these ILs demonstrated even stronger inhibitory effects. This observation was also noted when tetraalkylammonium compounds were used to inhibit hOCT2-mediated MPP and metformin transport; i.e., transport of metformin was inhibited to a greater degree (Dresser et al., 2002; Choi et al., 2007). The reason for the different inhibitory effects observed for the two probe substrates is still not clear. The transport characteristic of NBuPy-Cl by hOCT2 (Kt = 18 μM) is also significantly different from its inhibitory potency on hOCT2-mediated metformin uptake (IC50 = 0.7 μM). These results might be related to different binding sites of two chemicals within the large binding surface of OCTs and/or the influence of noncompetitive or other allosteric interactions (Koepsell et al., 2007).
ILs are called “designable chemicals” because they can be customized structurally to fit specific applications. These alterations in structure not only will change the chemical and physical properties of ILs but also may alter their disposition, transport, and toxicity. As demonstrated by the SAR results presented here, the inhibitory effects of the pyridinium-based ILs changed dramatically as the length of the alkyl side chain was altered. The IC50 values decreased significantly with an increase in the number of carbons on the alkyl chain. This SAR has also been reported for other chemical classes (Ullrich, 1997; Bednarczyk et al., 2003; Suhre et al., 2005; Choi et al., 2007). It was suggested that the hydrophobicity of the ILs increased with increasing alkyl chain length (Ranke et al., 2007). This increased hydrophobicity could facilitate their interaction on the pharmacophore of OCTs and thus contribute to the enhanced inhibitory activity (Bednarczyk et al., 2003).
Results presented here show that NBuPy-Cl is also a potent inhibitor of hMATE1 and hMATE2-K. Because both OCTs and MATEs play critical roles in the renal secretion of organic cations, disruption of any of these transporters may alter the renal elimination profile of certain chemicals. For example, renal secretion of TEA was reduced in both rats and mice with impeded OCT1/2 function (Jonker et al., 2003; Matsuzaki et al., 2008). Likewise, plasma concentrations of metformin were increased in MATE1 knockout mice, and its urinary excretion was significantly reduced (Tsuda et al., 2009). Thus, it is not surprising that infusion of NBuPy-Cl markedly altered the plasma pharmacokinetics of metformin. It has the capacity to inhibit the transport processes related to both the uptake of metformin into proximal tubules and its extrusion from these cells into the tubular lumen.
Metformin is a comparatively low-affinity substrate for hOCT1, hOCT2, hMATE1, and hMATE2-K, with Kt values of 1.47, 0.99, 0.78, and 1.98 mM, respectively (Koepsell et al., 2007; Tanihara et al., 2007). As determined here, NBuPy-Cl blocks both OCTs and MATEs with IC50 values in the low micromolar range. Based on the equation for calculating the steady-state plasma concentration after intravenous infusion (Rowland and Tozer, 1995), the estimated steady-state concentration of NBuPy-Cl in plasma is 0.8 μg/ml (4.7 μM), with an infusion dose of 2 mg/kg per hour. Therefore, it was not surprising that renal clearance of metformin was reduced in the presence of this compound. Indeed, the NBuPy-Cl inhibitory profile suggests that a plasma concentration of ∼5 μM would block approximately half of the rOCT1 (IC50 = 4.7 μM) and rOCT2 (IC50 = 3.8 μM) activity. Because of the negative intracellular potential (∼70 mV) of renal proximal tubule cells, NBuPy-Cl, the positively charged moiety, could reach substantially higher levels intracellularly and inhibit MATEs more extensively. Thus, exit of metformin from renal proximal tubule cells may have been rate-limiting under these conditions. In fact, the exit step was shown to be rate-limiting in renal secretion of organic cations (Schäli et al., 1983), which is consistent with the accumulation of metformin in kidney tissue associated with the highest dose of NBuPy-Cl.
Because metformin is primarily eliminated in the urine as the parent compound and at a clearance rate that exceeds the GFR (Scheen, 1996), the increased plasma area under the curve of metformin in NBuPy-Cl-infused animals relates, at least in part, to the inhibitory effects of NBuPy-Cl on OCTs and/or MATEs. In our previous publication, we reported that NBuPy-Cl, at a single intravenous dose of 5 mg/kg, did not alter the plasma clearance of inulin in unanesthetized animals. This result indicated that NBuPy-Cl at this dose did not affect rat GFR. However, it should be cautioned that the GFR could have been decreased in animals coadministered NBuPy-Cl and the anesthetics that were used. For example, pentobarbital has been reported to decrease GFR (Walker et al., 1986). On the basis of the clearance of metformin in unanesthetized rats (1.4 ml/min per 100 g) reported by Choi et al. (2010) and that of the control rats (1.0 ml/min per 100 g) reported here, anesthesia could account for up to 30% of the reduced metformin clearance. Another factor that could reduce GFR is impairment of renal function because of renal toxicity. However, in studies with Bmim-Cl, a closely related ionic liquid, no evidence of renal toxicity was observed (as assessed by both serum chemistry and histopathology) at oral doses up to 50 mg/kg. This dose, which was >50% bioavailable, maintained blood levels of Bmim-Cl at ∼2 μg/ml for several hours, close to the estimated steady-state blood levels of NBuPy-Cl reported here. Thus, the in vivo data presented here support the hypothesis that ionic liquids have the potential to inhibit renal transporters and alter the pharmacokinetics of substrates of these transporters.
It should be noted that the doses of NBuPy-Cl infused into rats were high. It is unlikely that such blood levels would be achieved and maintained in humans exposed orally and/or dermally to environmental/occupational levels of NBuPy-Cl or other ILs. In rats, dermal absorption of these three ILs was less than 35% of the applied dose (5 mg/kg, 125 μg/cm2), and the absorbed dose was readily eliminated. Dermal absorption in humans is not expected to exceed that observed for rats. It is clear that additional studies are needed to focus on how alterations in structure affect absorption of ILs after oral dosing or dermal application. More lipophilic ILs may achieve higher internal concentrations. This action, coupled with the greater inhibitory effects on OCTs, could influence their pharmacokinetic parameters as well as drug-chemical interactions.
In summary, in in vitro studies, NBuPy-Cl strongly inhibited OCTs and MATEs. In in vivo studies with rats, this inhibition resulted in a reduction in the plasma clearance of metformin. The structurally related ILs, Bmim-Cl and BmPy-Cl, and pyridinium-based ILs with increasing alkyl chain length, were also inhibitors of OCTs. The inhibitory effect increased as the length to the alkyl chain increased.
Authorship Contributions
Participated in research design: Cheng, Wright, Kuester, and Sipes.
Conducted experiments: Cheng and Martinez-Guerrero.
Contributed new reagents or analytic tools: Hooth.
Performed data analysis: Cheng, Martinez-Guerrero, and Sipes.
Wrote or contributed to the writing of the manuscript: Cheng, Wright, Hooth, and Sipes.
Acknowledgments
We thank Dr. Michael Cunningham (National Toxicology Program and National Center for Toxicogenomics, National Institute of Environmental Health Sciences) for his advice and support. We also thank Xiaohong Zhang (Department of Physiology, University of Arizona) for her assistance in cell transfection work, as well as Dr. Helen Cunny and Dr. Matt Stout (National Toxicology Program, National Institute of Environmental Health Sciences) for their critical review of this manuscript and Dr. Binfeng Xia (University of South Carolina) for his support in the pharmacokinetic modeling.
Footnotes
This work was supported by the National Institutes of Health National Institute of Environmental Health Sciences [Contract N01-ES45529] (National Toxicology Program); the National Institutes of Health National Institute of Diabetes and Digestive and Kidney Diseases [Grant DK58251]; and the National Institutes of Health National Institute of Environmental Health Sciences [Grant ES06694].
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
doi:10.1124/dmd.110.035865.
-
ABBREVIATIONS:
- IL
- ionic liquid
- SAR
- structure-activity relationship
- NBuPy-Cl
- N-butylpyridinium chloride
- Bmim-Cl
- 1-butyl-3-methylimidazolium chloride
- BmPy-Cl
- N-butyl-N-methylpyrrolidinium chloride
- OCT
- organic cation transporter
- MATE
- multidrug and toxic extrusion transporter
- Py-Cl
- pyridine hydrochloride
- EtPy-Cl
- 1-ethylpyridinium chloride
- HePy-Cl
- 1-hexylpyridinium chloride
- TEA
- tetraethylammonium trifluoroacetate
- MPP
- 1-methyl-4-phenylpyridinium
- CHO
- Chinese hamster ovary
- r
- rat
- h
- human
- LSC
- liquid scintillation counter
- GFR
- glomerular filtration rate
- Kt
- concentration of substrate that results in half-maximal transport.
- Received August 13, 2010.
- Accepted June 1, 2011.
- U.S. Government work not protected by U.S. copyright





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}