


Abstract
HMR1098, a novel KATP-blocking agent, is metabolized to form an S-glucuronide in rat and dog bile. Synthesis of the S-glucuronide metabolite was studied in human liver and kidney microsomes. Recombinant UPD-glucuronosyltransferases (UGTs) were screened for activity, and kinetic analysis was performed to identify the isoform or isoforms responsible for the formation of this novel S-glucuronide in humans. S-Glucuronidation is relatively rare, but from this study it appears that S-glucuronides are not generated exclusively by a single UGT isoform. From the panel of recombinant isoforms used, both UGT1A1 and UGT1A9 catalyzed the glucuronidation of HMR1098. The Vmax values in both instances were similar, but the Km for UGT1A1 was substantially lower than that measured for UGT1A9, 82 μM compared with 233 μM, respectively. Liver and kidney microsomes displayed similar Km values, but the Vmax in kidney was more than 20-fold less than in liver microsomes, which is suggestive of a significant role for the bilirubin UGT in catalysis of HMR1098, although other UGTs may play a secondary role.
HMR1098 is a new KATP-blocking agent being developed as a drug for prevention of sudden cardiac death by Aventis Pharma Deutschland GmbH. Metabolic studies in rats, dogs, and humans have shown that the main metabolite, M1, was characterized by 2D1 NMR as HMR1098-S-β-d-glucuronide (see this paper), and the glucuronide was linked via the thiourea group (Fig. 1). S-Glucuronides are rare metabolites, and the individual UDP-glucuronosyltransferases (UGTs) catalyzing their formation have never been identified.

The structure of HMR1098 and the formation of HMR1098-S-glucuronide.
UGTs are a family of microsomal enzymes in humans catalyzing the glucuronidation of thousands of compounds by transferring glucuronic acid from UDP-glucuronic acid to the aglycone substrate (Burchell et al., 1997). The substrate specificity of individual UGTs has been partially characterized by study of cloned/expressed transferases. This work has determined that UGT subfamily 1 is responsible for glucuronidation of bilirubin, phenols, and drugs, whereas subfamily 2 enzymes catalyze glucuronidation of steroids, bile acids, and drugs (Burchell et al., 1997). Key probe substrates such as bilirubin for UGT1A1 are used to indicate the specific activity of an individual isoform (Burchell et al., 1995).
The formation of thiol drug glucuronides is a relatively rare event, and few examples have been reported up to 1980 (see Dutton, 1980). The antithyroid drug 6-n-propyl-2-thiouracil forms a confirmed S-glucuronide in rats (Lindsay et al., 1977). A pyrithone (Omedine) metabolite, 2-mercaptopyridine-N-oxide, was detected as an S-glucuronide in rat, rabbit, and rhesus monkey urine (Mitoma et al., 1983). The thiocarbonate herbicide, SUTAN [bis(2-methylpropyl)carbamothioic acid S-ethyl ester], was significantly metabolized to an S-glucuronide found in rat urine (Peffer et al., 1991). A recent search of the literature failed to reveal any reference to formation of S-glucuronides in humans.
In this paper we have used human liver and kidney microsomes and cloned expressed enzymes to identify a key UGT in the synthesis of HMR1098-S-β-d-glucuronide.
Materials and Methods
Chemicals and Reagents. Bilirubin was obtained from Sigma-Aldrich (Gillingham, Dorset, UK). Propofol and 1-naphthol were also purchased from Sigma-Aldrich. Morphine sulfate was obtained from Thorter and Ross Ltd., Linthwaite Laboratories (Huddersfield, UK). HMR1098 (sodium salt of HMR1883) and HMR1098-S-β-d-glucuronide were synthesized by Aventis Pharma Deutschland GmbH (Frankfurt, Germany) and kindly provided by Heinrich Englert. Ammonium acetate and acetonitrile (HPLC grade) were used as mobile phase, and all other chemicals were from BDH (Poole, Dorset, UK).
Chemical Synthesis of HMR1098-S-Glucuronide Methyl Ester.HMR1098 was dissolved in sodium methoxide/methanol and stirred with acetobromo-α-d-glucuronic acid methyl ester for 12 h at 25°C. The corresponding acetoester (S 96 3535) was isolated by silica gel chromatography. Subsequently, the glucuronide was deacetylated by hydrolysis in 2 M HCl in methanol for 40 h at 25°C. The HMR1098-S-glucuronide methyl ester was purified by silica gel chromatography. Total yield of the final product was 15% (S 96 3536).
Isolation and Identification of Major HMR1098 Metabolite, Ml. The major metabolite of HMR1098 in rat, dog, and human bile was M1. M1 was isolated by HPLC from frozen bile using a Nucleosil C18 column (Macherey-Hagel, Duren, Germany) and a mobile phase of 0.01 M ammonium acetate buffer, pH 6.5, and a methanol gradient of 10 to 70% (v/v).
NMR spectroscopy was done with an AC 200 with 4.7 T magnet and DPX400/500 with 9.4/11.7 T magnet (Bruker, Rheinstetten, Germany) for one-dimensional 1H NMR spectroscopy and two-dimensional heteronuclear shift correlation NMR spectroscopy. Samples were dissolved in dimethyl sulfoxide (DMSO)-D6 (Dentero GmbH, Heresback, Germany).
Human liver microsomal preparations were obtained from In Vitro Technologies Inc. (Baltimore, MD). Human kidney microsomes were obtained as previously reported (Soars et al., 2001).
Tissue Culture. V79 and recombinant cell lines were grown in Dulbecco's modified Eagle's medium containing 10% fetal bovine serum, 100 units/ml penicillin, and 0.1 mg/ml streptomycin. Cell cultures were grown in 75-cm2 flasks (Costar, Cambridge, MA) fitted with vented caps in humidified incubators at 37°C with the atmosphere maintained at 5% CO2. V79 cells heterologously expressing human UGTs were maintained under constant selection concentrations of geneticin (G418; Invitrogen, Paisley, Scotland).
Control UGT Assay Conditions. The typing of human liver and kidney microsomes and the assays used as controls for the LC/MS-MS assays were performed by the method of Ethell et al. (1998). Briefly, substrates were incubated with microsomes or recombinant UGTs in 100 mM Tris-maleate buffer containing 5 mM MgCl2, 2 mM UDPGA (0.1 μCi of [14C]UDPGA) for 40 min in a volume of 100 μl. After this time, reactions were terminated by the addition of 100 μl of prechilled methanol. Precipitated proteins were removed by centrifugation at 1000g for 10 min, and the 150 μl of supernant were injected onto a gradient HPLC system fitted with an online radioactivity flow monitor (Canberra Packard Ltd., Pangbourne, Berkshire, UK) fitted with a heterogeneous scintillant flow cell. Substrate concentrations varied according to the substrate used: 122 μM bilirubin, 1 mM morphine, 250 μM 5α-androsterone 3α,17 β-diol, hyodeoxycholic acid, propofol, and 1-naphthol at 500 μM. Microsomes and cell lines expressing human UGT isoforms were activated by sonication as previously described (Ethell et al., 1998).
LC/MS-MS Assays of HMR1098-Glucuronide Formation in Vitro. A 1 mg/ml solution of the synthesized HMR1098-glucuronide was prepared in 50:50 acetonitrile/water and diluted 1:1000. This solution was infused (Pump 11; Harvard Instruments, Edenbridge, Kent, UK) into the electrospray interface (Quattro LC; Micromass, Manchester, UK) in negative ion mode at a rate of 10 μl/min to generate the mass spectrum of the parent glucuronide. The [M - H]- ion was recorded (646.2 atomic mass units; Fig. 2A) and optimized for probe position as well as cone and capillary voltages. The spectrometer was then switched into MS-MS mode, the daughter ion spectrum was recorded using argon as the collision gas, and the collision energy was optimized. Daughter ion spectra at 15- and 10-eV collision energies are shown in Fig. 2, B and C. At 10 eV, some parent ion remained but had been completely fragmented by 15 eV. The most abundant ion in negative ion mode was the fragment at 209 (Fig. 2, B and C), and this was selected for the multiple reaction monitoring transition for this method.

Parent ion and daughter ion spectra of HMR1098.
A, the parent ion spectrum of HMR1098; B and C, the daughter ion spectra of HMR1098 S-β-d-glucuronide at 15 eV and 10 eV, respectively. The fragmentation pattern of the glucuronide is illustrated on the chemical structure.
The LC conditions (HP1100; Agilent Technologies, Stockport, UK) for the analysis of glucuronide conjugates comprised a binary gradient of 10 mM ammonium acetate in acetonitrile from 0 to 100% developed over 7 min on a 2.1 mm × 15 cm Spherisorb ODS2 column at a flow rate of 0.3 ml/min. This flow rate did not necessitate a flow split, and the column was coupled directly to the electrospray source. Calibration was performed by diluting the glucuronide standard to give quantities of 20, 10, 5, 2, and 1 ng in 10-μl injections. The resulting calibration curve was linear with a correlation coefficient of >0.99.
Incubation of HMR1098 with microsomes and recombinant UGTs was performed under the same conditions as the control assays. Owing to the limited quantity of HMR1098 and microsomes, the incubations were scaled down to 40 μl and were stopped with an equivalent of prechilled methanol.
Detection was easily achieved by injection of 10 μl of supernatant onto the LC/MS-MS system.
Protein Determination. Protein determinations on the microsomal preparations and for the sonicated cells were performed by the method of Lowry et al. (1951).
Results
Isolation and Identification of the Major Metabolite (M1) of HMR1098. M1 was isolated from bile by HPLC after administration of HMR1098. LC-MS analysis had indicated M1 was a conjugate with β-d-glucuronic acid (see Fig. 1 and 2), but there was no evidence to indicate whether the glucuronide was connected via the nitrogen or sulfur sites. One- and two-dimensional NMR experiments were performed with M1, and a synthesized methylated HMR1098-S-glucuronide derivative was used for comparison.
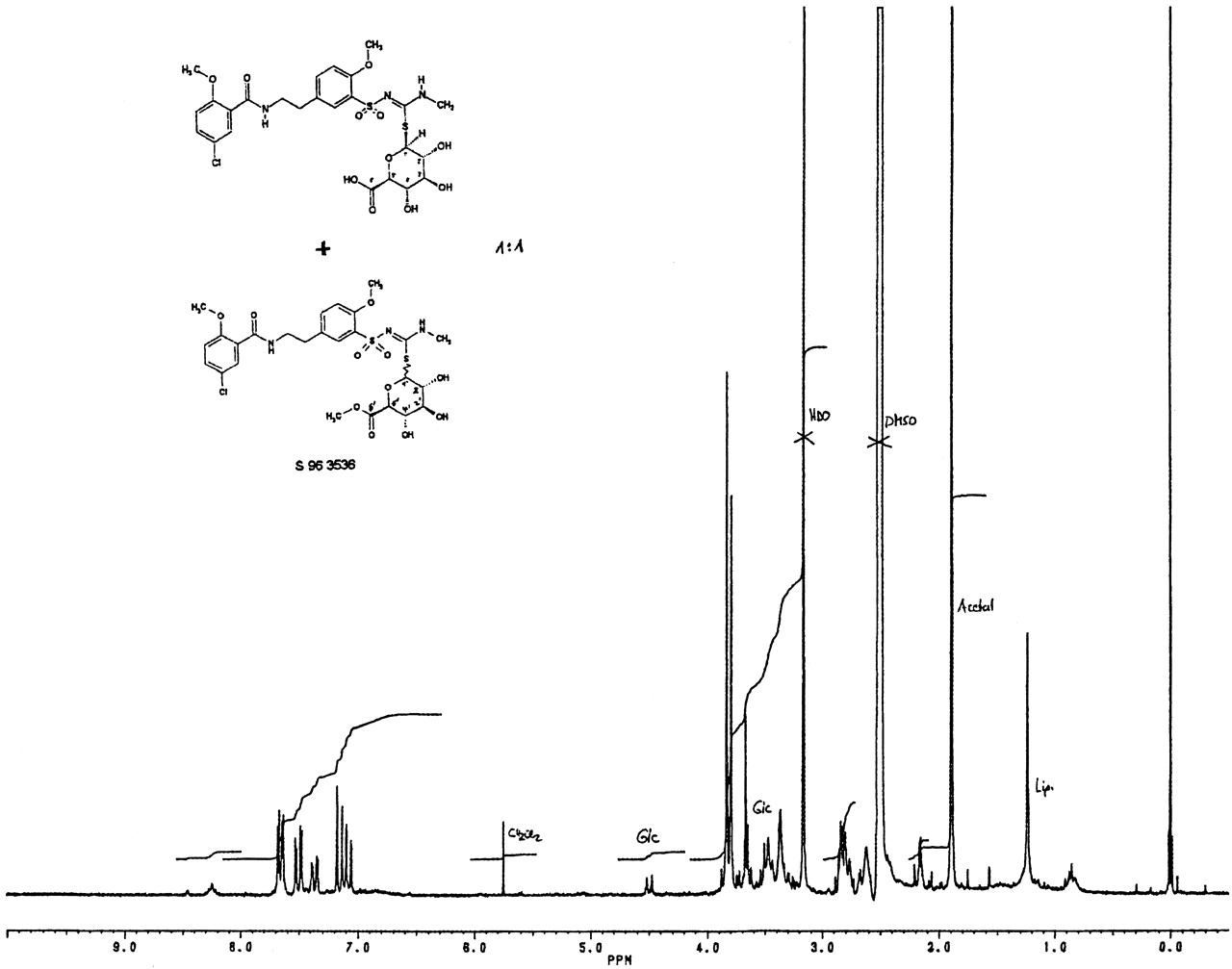
One-dimensional 1H NMR spectroscopy of M1 was performed to identify the 1′-proton of the β-d-glucuronic acid (Fig. 3). The assignments were also based on the shifts for HMR1098. This mixed sample indicates the presence of the chemical S-conjugated β-d-glucuronic acid in M1. No further signal indicating an N-conjugated β-d-glucuronic acid could be observed.

HMR1098-S-glucuronide.
One-dimensional 1H 200 MHz spectrum of a 1:1 mixture of isolated metabolite M1 and the synthetic S-glucuronic acid derivative S 96 3536 in DMSO-D6.
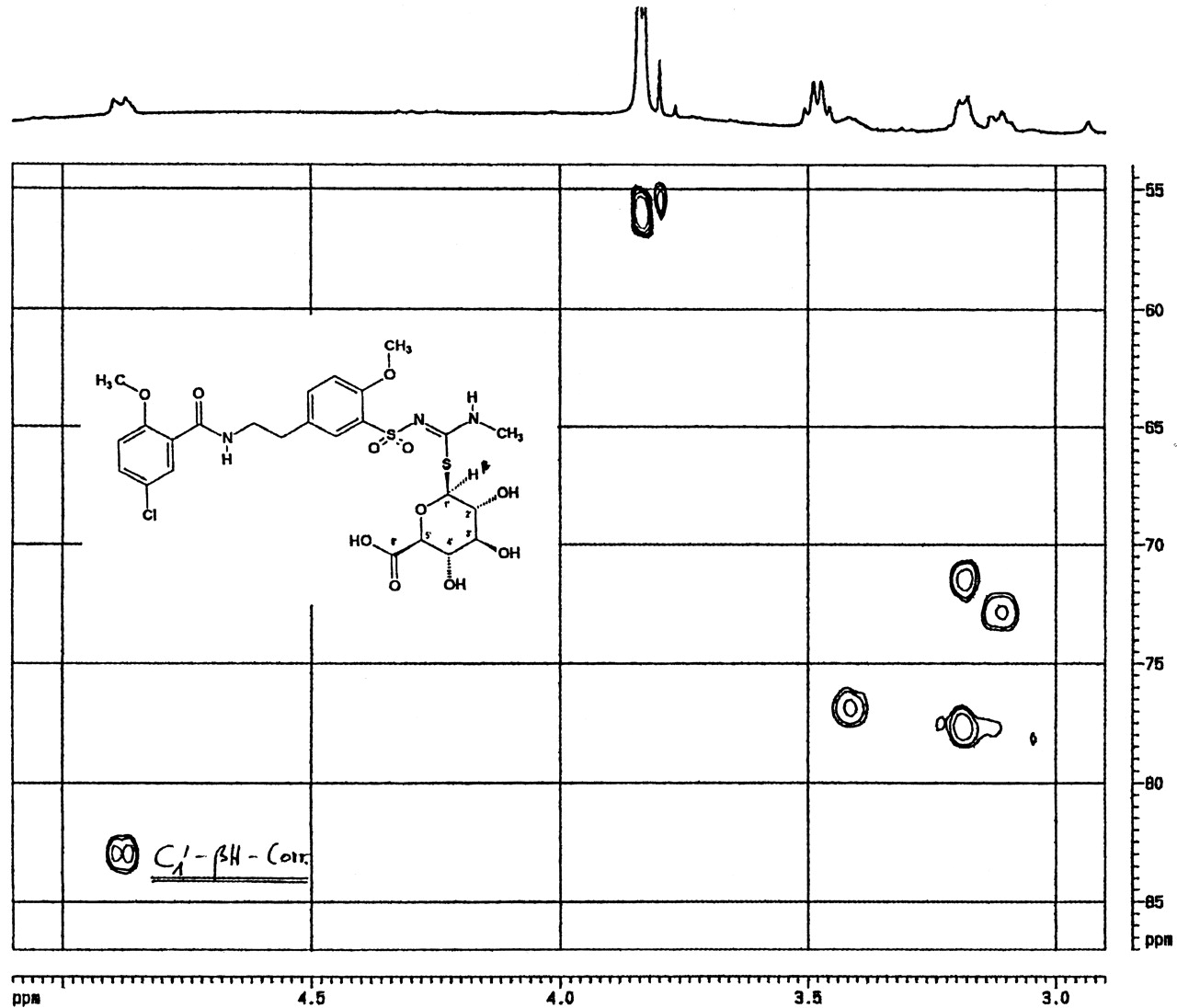
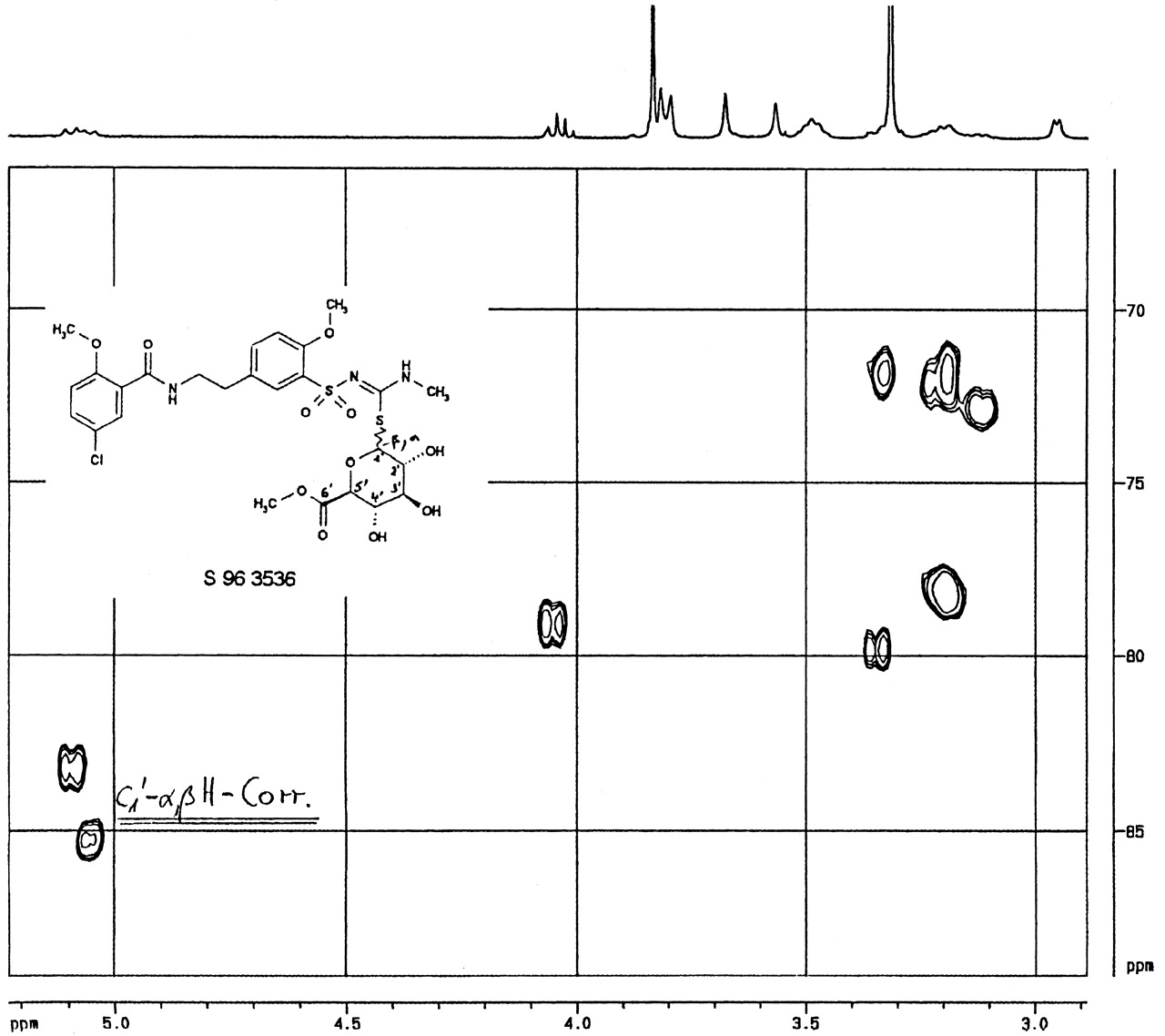
Additionally, the metabolite M1 was investigated by two-dimensional C1H-correlated NMR spectroscopy (Fig. 4) in comparison with the synthetic S-glucuronide derivative S 96 3536 (Fig. 5). The spectral 13C-region (δ≈80) in which the 1′H-proton correlates with the C1-carbon was found to be at 83 ppm for M1 (Fig. 4) and at 83.5/84.5 ppm for the S-glucuronide derivative (Fig. 5), indicating that both compounds were connected via the sulfur-site. The doublet signal at 84 ppm for the synthetic S-glucuronide derivative in Fig. 5 indicated that both anomeric forms of the conjugated glucuronic acid were made during the chemical synthesis. Using NMR spectroscopy as described, it was possible to identify the HMR1098 metabolite M1 as S-β-d-glucuronic acid conjugate of the parent compound.

HMR1098-S-glucuronide.
2D C1H correlated 400-MHz NMR spectrum of isolated metabolite M1 in DMSO-D6.

HMR1098-S-glucuronide methyl ester.
2D C1H correlated 400 MHz NMR spectrum of the synthetic S-glucuronic acid derivative S 96 3536 in DMSO-D6.
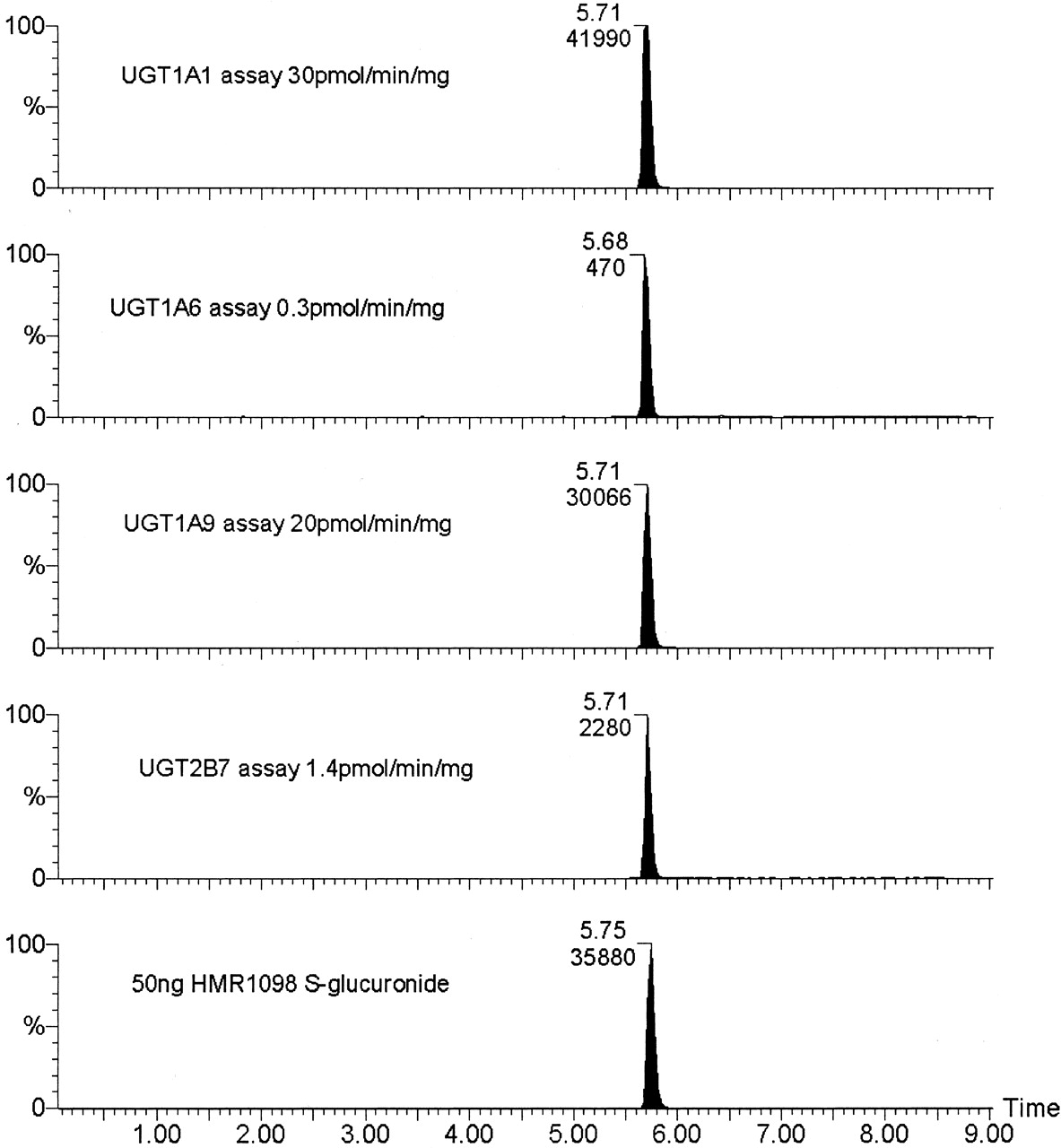
In Vitro Synthesis and Detection of HMR1098-S-Glucuronide by Mass Spectroscopy. The elution of the glucuronides, which were formed by in vitro incubation of HMR1098, was chromatographically identical to the synthesized reference material provided by Aventis. The fragmentation of the parent ion to the daughter ion described under Materials and Methods (Fig. 2) was highly selective for this particular compound, leaving no doubt that what is in fact formed in these reactions is a genuine S-glucuronide. Figure 2 shows the molecular fragmentation releasing three characteristic daughter ions, S-glucuronic acid m- = 209 and glucuronic acid m- = 175, and how the ion m- = 131 was generated. The S-glucuronic acid is a characteristic ion used in the mass spectrophotometric assay (see Materials and Methods). Comparison of the standard synthesized reference glucuronide with the product from the UGT assays is shown in Fig. 6. Assays incubated in the absence of UDPGA had no peak present at 5.7 min, which corresponded to the presence of HMR1098-S-glucuronide.

LC/MS-MS chromatograms of HMR1098 glucuronidation assays of human UGT1A1, UGT1A6, UGT1A9, and UGT2B7.
The enzyme activities for the recombinant enzymes with control substrates were UGT1A1, octylgallate, 1.3 nmol/min/mg of protein; UGT1A6, 1-naphthol, 1.8 nmol/min/mg of protein; UGT1A9, propofol, 0.85 nmol/min/mg of protein; UGT2B7, hydeoxycholic acid, 0.38 nmol/min/mg of protein.
Synthesis of HMR1098-S-Glucuronide by Human Tissue Microsomes. The human microsomes used were characterized by assaying with standard marker substrates of known specificity to demonstrate that they were competent for glucuronidation activity (see legend to Fig. 7). Propofol glucuronidation has been used as a marker for the presence of UGT1A9 in liver and kidney microsomes, and here the levels of UGT activity toward propofol are similar in microsomes from both hepatic and renal tissue. However, kidney microsomes do not catalyze bilirubin glucuronidation, which signifies the absence of human UGT1A1 protein from kidney (McGurk et al., 1998; Soars et al., 2001).

The kinetic determinations for the glucuronidation of HMR1098 in human liver microsomes (A) and human kidney microsomes (B).
Activities for human tissue microsomes using control substrates and 2 mM UDP-glucuronic acid were with bilirubin (122 μM), 0.59 nmol/min/mg of protein in liver, zero activity in kidney; propofol (500 μM), 0.19 nmol/min/mg of protein in liver, 0.3 nmol/min/mg of protein in kidney; morphine (1000 μM), 1.7 nmol/min/mg of protein in liver, 0.05 nmol/min/mg of protein in kidney; and 1-naphthol (500 μM), 2.4 nmol/min/mg of protein in liver, 0.7 nmol/min/mg of protein in kidney.
Screening of human liver microsomes showed that both human liver and kidney were capable of catalyzing the glucuronidation of HMR1098. Kinetic analyses revealed that both liver and kidney microsomes catalyzed glucuronidation of HMR1098 with similar Km values: liver microsomes, -295 μM; kidney microsomes, -256 μM (Fig. 7). However, the maximal rate of glucuronidation was substantially lower in kidney, only 7.3 pmol/min/mg, compared with liver, 161 pmol/min/mg (Fig. 7).
Formation of HMR1098-S-Glucuronide Catalyzed by Expressed UGT Isoforms. The Vmax for glucuronidation of HMR1098 by UGT1A1 and UGT1A9 was 70 pmol/min/mg and 44 pmol/min/mg, respectively, and the Km for UGT1A1 was 82 μM compared with a Km of 233 μM for UGT1A9 (Fig. 8).

The kinetic determinations for the glucuronidation of HMR1098 by recombinant human UGT1A1 (A) and UGT1A9 (B).
Km = 82 μM and Vmax = 70 pmol/min/mg of protein for UGT1A1 and Km = 233 μM and Vmax = 44 pmol/min/mg of protein for UGT1A9.
Of the four isoforms screened, UGT1A6 and UGT2B7 did not glucuronidate HMR1098 significantly, although a low rate of synthesis was detected by this sensitive method (Fig. 6). The two other UGT1A1 isoforms, UGT1A1 and UGT1A9, both showed high activity toward this substrate. The activities for the control substrates are given in the legend to Fig. 6.
Discussion
The glucuronide of parent HMR1098 metabolite M1 was isolated from dog bile. The exact structure of M1 could be identified as HMR1883-S-β-d-glucuronide by NMR (abbreviated HMR1883-S-glucuronide). The presence of an N-glucuronide in isolated M1 fraction was excluded.
M1 is the predominant metabolite of the drug in dog bile. Based on preliminary evaluations in the actual ongoing kinetic and metabolism study, the parent compound accounted for about 9% of the given dose until 8 h after administration. Assuming the same UV absorption for M1 as for parent HMR1098, at least 40% of the dose was estimated for metabolite M1. Additional metabolites in small amounts were also present. The 0.5- to 1-h bile sample contained the highest concentration on M1.
HMR1098-glucuronide is also the predominant metabolite in human and rat bile in vivo and in rat hepatocytes in vitro. This glucuronide and the present identified M1 have an identical behavior in the HPLC system used. In contrast to bile, M1 is only a minor metabolite in plasma and urine of dog and rat. This confirms that, after its synthesis, metabolite M1 is rapidly excreted via bile. In conclusion, the S-glucuronidation is important in the metabolism of HMR1098 in the dog and humans. The glucuronide (M1) is the predominant metabolite in bile and a minor compound in plasma and urine.
The use of human recombinant isoforms can greatly help in the identification of which UGT is involved in the glucuronidation of a new substrate. In the situation in which more than one isoform is involved, it is difficult to make an assessment of the relative importance of the two (or more). Using probe substrates in microsomes from different tissues alongside the assays with human recombinant enzymes can help to estimate the relative contribution of each isoform.
From the assays with human recombinant UGTs, it can clearly be seen that UGT1A1 and UGT1A9 are capable of glucuronidating HMR1098 at much higher rates than UGT1A6 and UGT2B7. The kinetic parameters measured for these two isoforms show a 2.8-fold lower Km and a 1.6-fold higher Vmax with UGT1A1 than with UGT1A9, resulting in 4.5-fold higher intrinsic clearance by the bilirubin UGT (0.85 μl/min/mg) compared with UGT1A9 (0.19 μl/min/mg).
It is also known from previously published work that UGT1A1 is present at high levels in human liver but not in human kidney (Findlay et al., 2000) (as demonstrated again in this report by the absence of bilirubin activity). It has also been reported that UGT1A9 is expressed highly in human kidney and liver. Propofol was used as a probe substrate for UGT1A9 in these tissues, and the similar levels of activity toward this substrate suggest similar levels of expression.
These data suggest that UGT1A9 may be the only significant isoform in kidney that catalyzed glucuronidation of HMR1098, because of the similar Km values of UGT1A9 and kidney microsomes. The 20-fold higher Vmax measured in human liver microsomes might be attributable to the presence of another UGT isoform. The human recombinant data suggest that UGT1A1 is a significant contributor to the greater HMR1098 activity in liver microsomes, but there is also the possibility that other UGTs such as UGT1A9 and UGT2B7 may be contributing to S-glucuronidation of HMR1098 in liver microsomes, and their contribution to the synthesis of HMR1098-glucuronide may cause the high Km observed in comparison with that of expressed UGT1A1.
The formation of S-glucuronides is rare, and of the examples that have been reported, most involve the glucuronidation of aromatic thiols like 4-nitrothiophenol, 2-aminothiophenol, and 2-mercaptopyridine-N-oxide by non-human species (Illing and Dutton, 1973; Mitoma et al., 1983; Smith et al., 1992). The thiourea functionality in HMR1098, which acts as the nucleophile, is also present in another compound, 6-n-propyl-2-thiouracil, which has been reported to form S-glucuronides in rats, although in this case the thiourea functionality is present in a cyclic system.
S-glucuronide formation has been demonstrated for the first time using human microsomes and human recombinant UGT enzymes. The significance of this finding is increased when considering that this compound is not merely a simple thiophenol derivative but a more complex molecule developed as a potential drug. This report indicates that compounds that contain this functionality could be subject to glucuronidation as a major aspect of their metabolism, as has been demonstrated for HMR1098.
Acknowledgments
We thank the Wellcome Trust and Aventis for support of this work.
Footnotes
-
↵1 Abbreviations used are: 2D, two-dimensional; UGT, UDP-glucuronosyltransferase; HPLC, high-performance liquid chromatography; DMSO, dimethyl sulfoxide; LC, liquid chromatography; MS-MS, tandem mass spectrometry; UDPGA, UDP-glucuronic acid; eV, electron volt.
- Received October 28, 2002.
- Accepted April 16, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}