


Abstract
In a caffeine test previously performed with healthy Japanese volunteers, we found that the CYP1A2 index defined as urinary {5-acetylamino-6-amine-3-methyluracil (AAMU) + 1-methylxanthine (1X) + 1-methyluric acid (1U)}/1,7-dimethyluric acid (17U) was affected by the whole deleted allele of CYP2A6 (CYP2A6*4). Since the high value of the CYP1A2 index could be caused by a low urinary concentration of 17U, we postulated that CYP2A6 was responsible for the 1,7-dimethylxanthine (17X) metabolism to generate 17U (17X 8-hydroxylation). Thus, the role of CYP2A6 in the 17X 8-hydroxylation was fully examined in the present study. Among 10 isoforms of human cytochrome P450 (CYP1A2, CYP2A6, CYP2B6, CYP2C8, CYP2C9, CYP2C19, CYP2D6, CYP2E1, CYP3A4, or CYP3A5) expressed in Escherichia coli cells, CYP2A6 and CYP1A2 showed high catalytic activities for the 17X 8-hydroxylation. The 17X 8-hydroxylase activities significantly associated with coumarin 7-hydroxylase activities (r = 0.67, p < 0.01) in liver microsomes from 17 individuals, but not with ethoxyresorufin O-deethylase activities. Tranylcypromine, an inhibitor of CYP2A6, reduced the 17X 8-hydroxylase activities of human liver microsomes. The 17X 8-hydroxylase activities of CYP2A6.7, CYP2A6.10, and CYP2A6.11 expressed in E. coli cells were 12, 13, and 22% of that of CYP2A6.1, respectively. The 17X 8-hydroxylase activities were found to be low in liver microsomes from individuals possessing the deletion or mutations in the CYP2A6 gene. Based on these data, we conclude that CYP2A6 is a main 17X 8-hydroxylase and that the catalytic activities for the 17X 8-hydroxylation are reduced by the genetic polymorphisms of the CYP2A6 gene.
Caffeine (1,3,7-trimethylxanthine, 137X) is extensively transformed in vivo into a wide variety of metabolites in humans. Since caffeine is a commonly ingested and relatively innocuous compound, the amounts of caffeine metabolites in human urine have been determined as a useful marker to estimate the activities of drug-metabolizing enzymes involved in the caffeine metabolism, including cytochrome P450 1A2 (CYP1A2), N-acetyltransferase 2, and xanthine oxidase, simultaneously (Kalow and Tang, 1993).
In our in vivo caffeine test previously performed with healthy Japanese volunteers (Saruwatari et al., 2002), we found that the distribution of the CYP1A2 index defined as urinary {5-acetylamino-6-amine-3-methyluracil (AAMU) + 1-methylxanthine (1X) + 1-methyluric acid (1U)}/1,7-dimethyluric acid (17U) showed a curvilinearity. No relationship could be found between the distribution of the CYP1A2 index and the genetic polymorphisms of the CYP1A2 gene which altered the inducibility of CYP1A2. However, we found that the whole deleted allele of the CYP2A6 gene (CYP2A6*4) existed in the subjects who showed a high value of the CYP1A2 index (Saruwatari et al., 2002). Since the high value of the apparent CYP1A2 index was caused by a low urinary concentration of 17U, we assumed that CYP2A6 would be responsible for 1,7-dimethylxanthine (17X) 8-hydroxylation to yield 17U as shown in Fig. 1. Thus, we considered that the determination of urinary caffeine metabolites could be of use for the estimation of the in vivo CYP2A6 activity.
CYP2A6 is the principal enzyme involved in the metabolism of nicotine (Nakajima et al., 1996), coumarin (Miles et al., 1990), tegafur (Komatsu et al., 2000), and fadrozole (Pelkonen et al., 2000). Large interindividual variations in the activities of CYP2A6 have been noted in humans (Rautio et al., 1992). This interindividual variation can be explained, at least, in part, by the CYP2A6 genetic polymorphisms (Inoue et al., 2000). Many variant alleles of the CYP2A6 gene have been reported to date (http://www.imm.ki.se/CYPalleles/cyp2a6.htm). Our laboratory has found the whole gene deletion-type mutants that cause the lack of enzymatic activity (Nunoya et al., 1999a,1999a,b), as well as the single nucleotide polymorphisms (CYP2A6*7 and CYP2A6*11) that reduce the in vivo and in vitro metabolic capacity (Ariyoshi et al., 2001b; Daigo et al., 2002). In addition, CYP2A6*9, which contains a –48T to G nucleotide substitution in the TATA box of the 5′-flanking region of the CYP2A6 gene (Pitarque et al., 2001), was reported to reduce the expression levels and the catalytic activities of the CYP2A6 (Kiyotani et al., 2003). Recently, the CYP2A6*10 allele containing the amino acid substitutions of both Ile471Thr and Arg485Leu was also found (Xu et al., 2002; Yoshida et al., 2002). The subjects possessing the CYP2A6*10 allele showed a lowered capacity to metabolize nicotine (Xu et al., 2002; Yoshida et al., 2002). Based on these genetic data, individuals can be classified into three groups, i.e., poor metabolizers (PMs), intermediate metabolizers, or extensive metabolizers, according to the genotypes. PMs for CYP2A6 have a reduced metabolic capacity, which can result in higher plasma concentration and the increased risk of adverse effects at ordinary drug dosages, whereas extensive metabolizers do not achieve therapeutic drug levels. In addition, prodrugs such as tegafur need to be metabolically activated by CYP2A6 and are therefore inactive in PMs. Actually, previous reports have shown that the in vivo metabolism of nicotine, coumarin, and tegafur was affected by the genetic polymorphisms of the CYP2A6 gene (Daigo et al., 2002; Xu et al., 2002). Therefore, if a caffeine test was available for the phenotype analysis of CYP2A6, it would provide clinically important information for individuals. However, it has not been completely clarified as to whether CYP2A6 would be involved in the 17X 8-hydroxylation.

Metabolic pathways of caffeine and its primary metabolite, 17X, in humans.
In this study, we investigated the role of CYP2A6 involved in the 17X 8-hydroxylation by using genetically engineered Escherichia coli cells expressing P450, together with NADPH-P450 reductase (CPR) and human liver microsomes. We document, herein, that CYP2A6 is a principal 17X 8-hydroxylase as well as that the genetic polymorphism of the CYP2A6 gene affects the 17X 8-hydroxylation.
Materials and Methods
Chemicals. 4-Acetoamidophenol, 17X, 17U, and 1X (Fig. 1) were obtained from Sigma (St. Louis, MO). NADP+, glucose 6-phosphate, and glucose-6-phosphate dehydrogenase were obtained from Oriental Yeast Co., Ltd, (Tokyo, Japan). All other chemicals were of the highest quality commercially available.
Enzyme Preparation. Human livers were obtained from patients after pathological examination of specimens isolated after death or during surgery (Nakamura et al., 2002). The use of the human livers for this study was approved by the ethics committee of Hokkaido University. Human liver microsomes were prepared in 10 mM Tris-HCl buffer (pH 7.4) containing 1.0 mM EDTA and 20% (v/v) glycerol as described previously (Yamazaki et al., 1999). Commercial human liver microsomes were obtained from XenoTech (Reaction Phenotyping Kit Ver. 5; Xenotech, Lenexa, KS).
Expression plasmids carrying each P450 (CYP1A2, CYP2A6, CYP2B6, CYP2C8, CYP2C9, CYP2C19, CYP2D6, CYP2E1, CYP3A4, or CYP3A5) or a mutant CYP2A6 (CYP2A6.7 or CYP2A6.11) cDNA together with CPR cDNA were constructed as described previously (Iwata et al., 1998; Ariyoshi et al., 2001a,b; Daigo et al., 2002; Yamaori et al., 2003). To introduce an amino acid substitution of CYP2A6.10, G-base was substituted by a T-base at the position of 1454 base pairs in the cDNA encoding CYP2A6.7 by the primer-directed enzymatic amplification method reported previously (Saiki et al., 1988; Ariyoshi et al., 2001b). The primers, 5′-CACtAAACTACACCATGAGCT-3′ and 5′-TAGTTTaGTGGGATCGTGGC-3′, were used to introduce the single nucleotide substitution, which codes for Arg485Leu in exon 9. The entire coding region, including the mutated site, was verified by sequencing with an ABI PRISM 3100 Genetic Analyzer (Applied Biosystems, Foster City, CA). Each isoform of P450 and CPR in the genetically engineered E. coli cells was expressed as described previously (Iwata et al., 1998; Ariyoshi et al., 2001a,b; Daigo et al., 2002; Yamaori et al., 2003). Membrane fraction was prepared from the E. coli cells according to the method reported by Sandhu et al. (1994). The membrane fraction was suspended in 100 mM Tris-HCl buffer (pH 7.5) containing 20% (v/v) glycerol and kept at –80°C until use. Each isoform of P450 in membranes had enough catalytic function toward typical substrates.
The contents of P450 in human liver microsomes and the membrane fraction of E. coli were determined spectrally by the method of Omura and Sato (1964). The protein contents were determined by using the Pierce BCA Protein Assay Kit (Pierce Chemical, Rockford, IL) with bovine serum albumin as a standard.
Enzyme Assays. The 17X 8-hydroxylase activity as shown in Fig. 1 was determined according to the method described below. The principal 17X concentration of 100 μM was chosen because blood concentrations of the parent compound, caffeine, were approximately 100 μM after intake of a cup of coffee (Campbell et al., 1987). A typical incubation mixture consisted of 100 mM potassium phosphate buffer (pH 7.4), 100 μM 17X, human liver microsomes (0.25 mg protein/ml), or the membrane fraction of E. coli expressing each P450 (80 pmol/ml) with cytochrome b5 (80 pmol/ml), and an NADPH-generating system (0.5 mM NADP+, 5 mM glucose 6-phosphate, and 1 unit/ml glucose phosphate dehydrogenase). Inhibitory effect of tranylcypromine on 17X 8-hydroxylation catalyzed by human liver microsomes was examined at a tranylcypromine concentration of 2 μM. Incubations were carried out at 37°C for 1 h and terminated by dichloromethane/2-propanol (4:1, v/v), and then 4-acetoamidophenol was added as an internal standard. The linearity of product formation for the incubation time was confirmed with recombinant CYP2A6 (for 90 min) and human liver microsomes (for 60 min). The solvent was evaporated after removal of protein by centrifugation. The residue was dissolved in 200 μl of solvent that was used as a mobile phase, 4% (v/v) CH3CN containing 10 mM CH3COONa (pH 4.0), and 100 μl of the sample was injected to a high-performance liquid chromatograph (L-7100 pump, L-7200 autosampler, and L-7400 UV detector; Hitachi, Tokyo, Japan) equipped with a Mightysil RP-18 GP Aqua column (150 × 4.6 mm, 5 μm; Kanto Chemical, Tokyo, Japan). Elution was performed at a flow rate of 1.0 ml/min. The formation of 17U was monitored at a wavelength of 280 nm. The detection limit of 17X was <0.5 pmol/ml or <1 pmol of product formation/min/nmol recombinant P450 under the present conditions.
The 17X 8-hydroxylase activity of 9000g supernatant (S9) fraction from E. coli cells expressing mutant CYP2A6 was also measured by the method described above. Kinetic parameters for the 17X 8-hydroxylation were estimated with a computer program (Microcal Origin; Microcal Software, Northampton, MA), designed for a nonlinear regression analysis.
Genotyping of theCYP2A6. Genomic DNA was isolated from the peripheral lymphocytes obtained from 111 healthy Japanese subjects and 42 human livers according to the method of phenol-chloroform extraction followed by ethanol precipitation (Sambrook et al., 1989). Genotyping of the CYP2A6 gene was carried out by the methods previously developed (Ariyoshi et al., 2000, 2001b; Kiyotani et al., 2003; Fujieda et al., 2004).
Caffeine Test. Apparent CYP1A2 index, defined as (AAMU + 1X + 1U)/17U, was calculated from data obtained from our in vivo caffeine test previously performed (Saruwatari et al., 2002). The sample population in this study comprised 111 CYP2A6-genotyped subjects extracted from 182 unrelated healthy Japanese in the previous caffeine test (Saruwatari et al., 2002). They were 68 males (including 16 smokers) and 43 females (including 2 smokers). The age of the subjects ranged from 20 to 40 years (mean age ± S.D., 22 ± 2.3 years). The body weight of the subjects ranged from 36 to 108 kg (mean weight ± S.D., 58.2 ± 10.6 kg). The number of cigarettes smoked ranged from 3 to 20 per day per smoker (mean ± S.D., 14 ± 5.6 per day). All of the volunteers provided written informed consent. The associations between the CYP1A2 index and the CYP2A6 genotypes were assessed by Dunnett's test. A p value <0.05 was considered to be statistically significant.
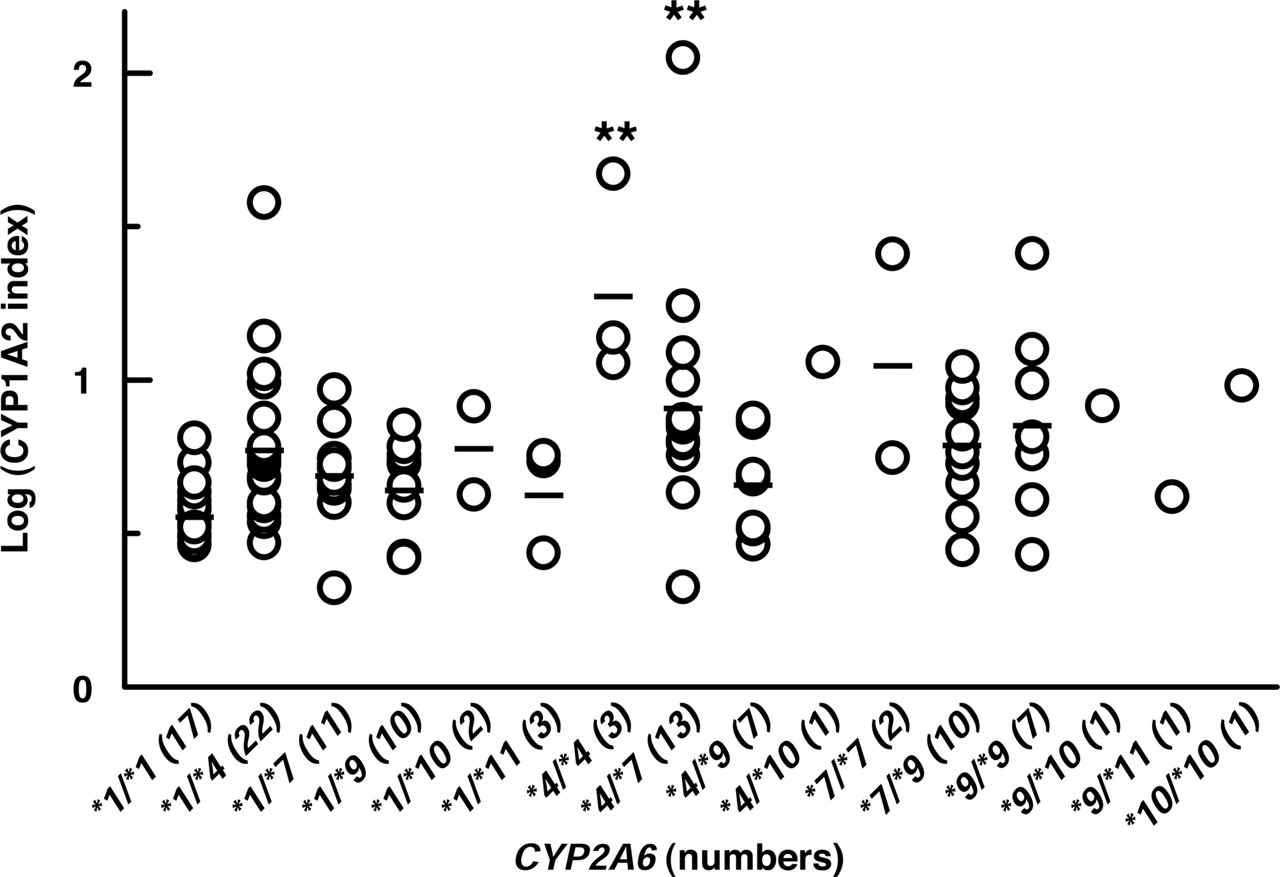
Relationship between CYP2A6 genotypes and log-transformed CYP1A2 index (AAMU + 1X + 1U)/17U obtained from 111 healthy Japanese volunteers. CYP2A6 genotypes were determined with genomic DNA isolated from peripheral blood samples. CYP1A2 index was determined by analyzing the caffeine metabolites in urine collected 8 h after oral administration of 150 mg caffeine. The bar represents the mean value of the group. The significance of the difference for the CYP1A2 index between each genotype carrying variant alleles and CYP2A6*1/*1 was evaluated by Dunnett's test (**, p < 0.01).
Results
Relationship between CYP2A6 Genotype and the CYP1A2 Index Obtained from Caffeine Test. Genomic DNA samples from 111 healthy Japanese subjects were analyzed for each CYP2A6 genotype to investigate whether or not the variant alleles of the CYP2A6 (CYP2A6*4, CYP2A6*7, CYP2A6*9, CYP2A6*10 and CYP2A6*11) influenced the CYP1A2 index. The association between the CYP2A6 genotype and the log-transformed CYP1A2 index is shown in Fig. 2. Analyzing the data by Dunnett's test, the apparent CYP1A2 indexes of subjects possessing CYP2A6*4/*4 and CYP2A6*4/*7 genotypes were significantly higher than that of subjects carrying CYP2A6*1/*1 genotype (** p < 0.01). Based upon this finding, we assumed that CYP2A6 would play an important role in the 17X 8-hydroxylation.
17X 8-Hydroxylase Activities of P450s Expressed inE. coliMembranes. The role of 10 forms of P450 in the 17X 8-hydroxylation was examined by using E. coli membranes, each expressing a form of human P450 and CPR (Fig. 3). The 17X 8-hydroxylase activities of CYP1A2 and CYP2A6 were 0.24 and 0.62 nmol/min/nmol P450, respectively, at a 17X concentration of 100 μM. Thus, the 17X 8-hydroxylase activity of CYP2A6 was 2.6 times higher than that of CYP1A2, CYP2C9, CYP2C19, and CYP3A4 were also involved in the 17X 8-hydroxylation. However, these activities were extremely low (<0.012 nmol/min/nmol P450) compared with CYP1A2 or CYP2A6. Representative HPLC chromatograms for the 17X metabolism catalyzed by the CYP2A6 or CYP1A2 expressed in E. coli membranes are shown in Fig. 4. Although 1X, a known metabolite of 17X (Kalow and Tang, 1993; Saruwatari et al., 2002), was also seen after the incubation of 17X, 17U was the main metabolite under the present assay condition.
17X 8-Hydroxylase in Human Liver Microsomes. To further clarify the contribution of CYP2A6 and CYP1A2 to the 17X 8-hydroxylation, correlation between 17X 8-hydroxylase activities and enzyme activities for typical substrates of CYP2A6 or CYP1A2 was examined with commercial human liver microsomes. Data of activities of coumarin 7-hydroxylase (CYP2A6), ethoxyresorufin O-deethylase (CYP1A2), diclofenac 4′-hydroxylase (CYP2C9), S-mephenytoin 4′-hydroxylase (CYP2C19), and testosterone 6β-hydroxylase (CYP3A) of the human liver microsomes were obtained from manufacturer's instructions. The 17X 8-hydroxylase activities significantly correlated with coumarin 7-hydroxylase activities (r = 0.67, ** p < 0.01; n = 17) (Fig. 5B). In contrast, no significant correlation was observed between the 17X 8-hydroxylase activities and ethoxyresorufin O-deethylase activities (Fig. 5A). No significant correlation between the 17X 8-hydroxylase activities and coumarin 7-hydroxylase activities was found in the presence of antibodies to CYP2A6 (data not shown). None of the other correlations were seen among 17X 8-hydroxylase and marker activities of CYP2C9, CYP2C19, or CYP3A4 (data not shown).

Activities of 10 isoforms of P450 to metabolize 17X to yield 17U. 17X (100 μM) was incubated at 37°C for 1 h with each P450 (80 pmol/ml) expressed in E. coli membranes.
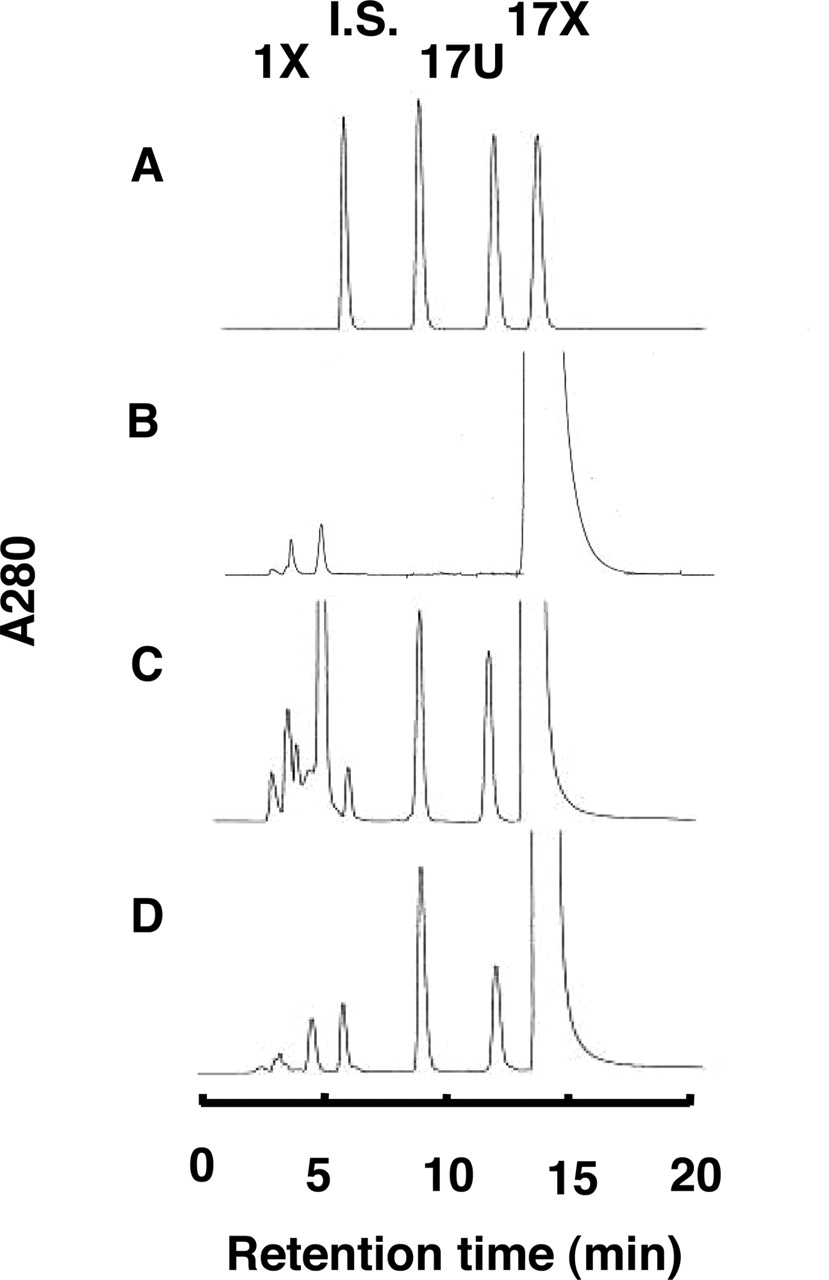
Representative HPLC chromatograms of 17X metabolites catalyzed by recombinant CYP2A6. A, authentic sample peaks formed from 17X as shown in Fig. 1. B, C, and D, 17X (100 μM) was incubated at 37°C for 1 h with control membranes and CYP2A6 and CYP1A2 (80 pmol/ml), respectively.
We examined the inhibitory effects of tranylcypromine, an inhibitor of CYP2A6 (Draper et al., 1997), on the 17X 8-hydroxylation using 16 preparations of human liver microsomes (Fig. 6). Tranylcypromine inhibited more than 80% of the 17X 8-hydroxylase activities of human liver microsomes except for one sample (sample 16).

Correlation between 17X 8-hydroxylase activities and enzyme activities for typical substrates of CYP1A2 or CYP2A6 in human liver microsomes. The 17X 8-hydroxylase activities of 17 human liver microsomes were plotted for ethoxyresorufin O-deethylase activities (CYP1A2) (A) or coumarin 7-hydroxylase activities (CYP2A6) (B).

Inhibitory effects of tranylcypromine on 17X 8-hydroxylation catalyzed by human liver microsomes. The 17X 8-hydroxylase activities of 16 human liver microsomes were determined in the absence (open bars) or presence (closed bars) of tranylcypromine (2 μM).
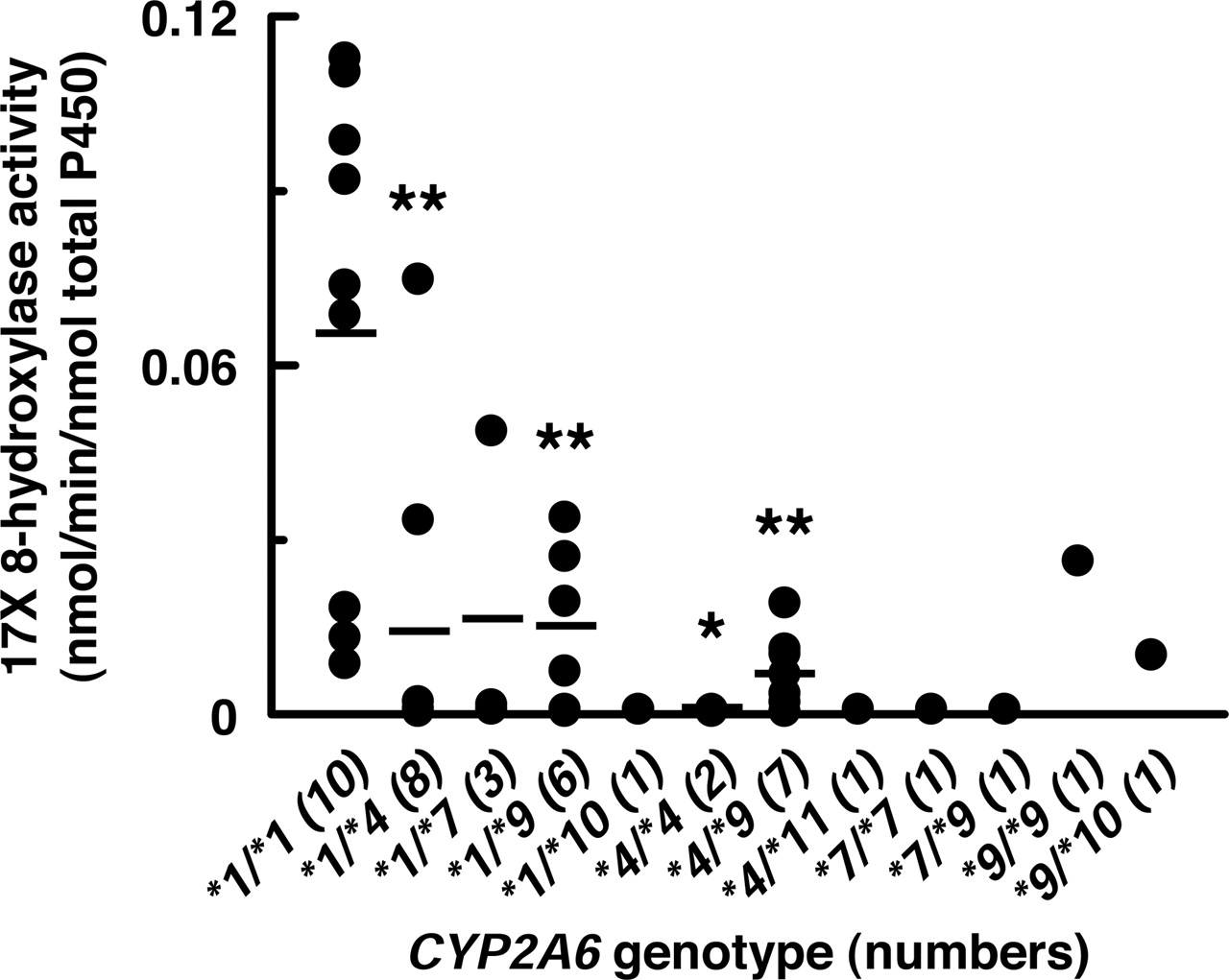
Association between Genetic Polymorphism of CYP2A6 and 17X 8-Hydroxylase Activities of Human Liver Microsomes. To investigate the effects of the genetic polymorphism of the CYP2A6 gene on the 17X 8-hydroxylation, we measured the 17X 8-hydroxylase activities of liver microsomes prepared from 42 Japanese subjects previously genotyped for CYP2A6 (Fig. 7). The 17X 8-hydroxylase activities of liver microsomes from subjects carrying mutant CYP2A6 genotype were remarkably low. The 17X 8-hydroxylase activities of liver microsomes derived from subjects genotyped as CYP2A6*4/*4 were not detectable. Liver microsomes prepared from individuals who harbored CYP2A6*1/*4, CYP2A6*1/*9, and CYP2A6*4/*9 showed significantly lower 17X 8-hydroxylase activities (** p < 0.01). Such lower activities were also seen in liver microsomes from the subjects possessing either the CYP2A6*7, CYP2A6*10, or CYP2A6*11 alleles, with no statistical significance by limited sample numbers.
Kinetic Analysis of Drug Oxidations Catalyzed by Mutant CYP2A6 Expressed in S9 Fractions ofE. coli. Kinetic analysis was performed for the 17X 8-hydroxylation catalyzed by the wild (CYP2A6.1) or mutant CYP2A6s (CYP2A6.7, CYP2A6.10 or CYP2A6.11) (Table 1). The Vmax values of each CYP2A6 protein for coumarin 7-hydroxylation were almost the same in our preliminary study (data not shown), showing that the systems of E. coli cells expressing each CYP2A6 protein were catalytically active. The Vmax and Km values of CYP2A6.1 for the 17X 8-hydroxylation were 0.19 ± 0.02 nmol/min/nmol CYP2A6 and 940 ± 220 μM, respectively. Kinetic parameters of CYP2A6.7, CYP2A6.10 and CYP2A6.11 for the 17X 8-hydroxylation could not be calculated because the activities of these CYP2A6 variants were not detected even at a substrate concentration of 1000 μM. Therefore, the 17X 8-hydroxylase activities at a 17X concentration of 2000 μM were shown in the table. These velocities of CYP2A6.7, CYP2A6.10, and CYP2A6.11 were 12, 13, and 22% of that of CYP2A6.1, suggesting that the 17X 8-hydroxylase activities were reduced by the genetic polymorphism of the CYP2A6 gene.
Kinetic parameters for 17X 8-hydroxylation catalyzed by recombinant CYP2A6 variants and CYP1A2
17X (200-2000 μM) was incubated with E. coli S9 fractions expressing CYP2A6 or CYP1A2. 17X 8-hydroxylase activities were determined as described under Materials and Methods. Kinetic parameters (mean ± S.E.) were calculated by nonlinear regression analysis.

Association between the genetic polymorphism of the CYP2A6 and 17X 8-hydroxylase activities of human liver microsomes. The 17X 8-hydroxylase activities were determined in 42 human liver microsomes at 17X concentrations of 100 μM. The bar represents the mean value of the group. The significance of the difference for 17X 8-hydroxylase activities between each of the genotypes carrying CYP2A6 variant alleles and wild-type CYP2A6*1/*1 was evaluated by Dunnett's test (**, p < 0.01, *, p < 0.05).
Discussion
Caffeine is extensively converted in vivo to its 1-, 3- or 7-demethylated metabolites in humans (Kalow and Tang, 1993). Among them, the 3-demethylation of caffeine to generate 17X, as shown in Fig. 1, catalyzed by CYP1A2 is the main metabolic pathway of caffeine (Gu et al., 1992; Kalow and Tang, 1993). Since it has been reported that CYP1A2 activities account for approximately 95% of the primary systemic caffeine clearance (Kalow and Tang, 1993), several methods for determining an individual's CYP1A2 activity by using caffeine as a probe have been developed with various urinary metabolite ratios such as (17X + 17U)/137X (Butler et al., 1992), (5-acetylamino-6-formylamine-3-methyluracil (AFMU) + 1X + 1U)/17X (Grant et al., 1983), (AFMU + 1X + 1U)/17U (Campbell., 1987), and (AAMU + 1X + 1U)/17U. Previous studies with white subjects have validated the (AAMU + 1X + 1U)/17U ratio more precisely in a variety of different conditions, revealing that this ratio was most closely correlated with the CYP1A2 activity of individuals (Kalow and Tang, 1993; Denaro et al., 1996).
When we recently performed the caffeine test and calculated the (AAMU + 1X + 1U)/17U ratio as the CYP1A2 index (Saruwatari et al., 2002), we found that the distribution of the CYP1A2 index showed a curvilinearity. No relationship could be found between the distribution of the CYP1A2 index and the genetic polymorphisms of the CYP1A2 gene. It has been reported that CYP1A2 was induced by smoking (Pantuck et al., 1974). We also found that Japanese smokers excreted high amounts of 1X in the urine, probably by CYP1A2 induced by smoking (data not shown). The average of the CYP1A2 index of smokers in the caffeine test was significantly higher than that of nonsmokers (* p < 0.05, data not shown), although the smoking could not explain the curvilinearity of distribution of the CYP1A2 index completely.
We investigated the effect of CYP2A6 activity on the CYP1A2 index because urinary concentration of the 17U, which was considered to be generated from 17X by CYP2A6 (Gu et al., 1992; Nowell et al., 2002), seemed to be the most important factor to influence the CYP1A2 index. Then, we could find that the genetic polymorphism of the CYP2A6 gene affected the apparent CYP1A2 index. In contrast to our results, the apparent CYP1A2 index was log-normally distributed in the studies with white subjects and failed to show any evidence of the genetic effects of the CYP2A6 gene (Kalow and Tang, 1991; Carrillo and Benitez, 1994). This interethnic difference may be caused by different frequencies of the CYP2A6 alleles between Japanese and white populations. The CYP2A6*1, CYP2A6*4, CYP2A6*7, CYP2A6*9, CYP2A6*10, and CYP2A6*11 alleles are popular in a Japanese population: the frequencies of these CYP2A6 alleles in Japanese are 42.0, 19.8, 12.6, 20.7, 4.3, and 0.7%, respectively (Fujieda et al., 2004). Based on these frequencies, 82.4% of Japanese are expected to possess any of these mutant alleles. Particularly, the frequency of Japanese subjects carrying the CYP2A6*4/*4 genotype is estimated to be 4.3%. Xu et al. (2002) have compared the frequency of CYP2A6 genotype between Japanese and white populations. They have reported that the frequencies of CYP2A6*1/*1 and CYP2A6*4/*4 were 57.1 and 7.9% in Japanese (n = 63), and 94.4 and 0% in whites (n = 301) (Xu et al., 2002), respectively, suggesting that the potential for detecting the genetic effects of CYP2A6 was minimized when the (AAMU + 1X + 1U)/17U ratio was applied to the white population. These conclusions on the roles of polymorphic CYP2A6 in caffeine metabolism in Japanese were consistent with the previous findings, indicating a significantly decreased 17U excretion in Orientals as compared to whites (Grant et al., 1983). Since we used this ratio for the first time in a caffeine test with Japanese subjects, we were able to find the effects of the genetic polymorphisms of the CYP2A6 gene on the apparent CYP1A2 index.
In general, coumarin and nicotine have been used for the CYP2A6 phenotyping because these substrates, at concentrations used in the phenotyping studies, were metabolized almost exclusively by CYP2A6 (Ujjin et al., 2002; Xu et al., 2002). In the present study, we propose that caffeine is useful as a probe for estimating an individual's CYP2A6 activities because the mutant alleles of CYP2A6, such as CYP2A6*4 and CYP2A6*7, are frequently found among the subjects who showed high values of the apparent CYP1A2 index in our previous caffeine test (Fig. 2) (Saruwatari et al., 2002). In addition, caffeine is generally ingested from coffee or tea and is a relatively innocuous compound, whereas coumarin and nicotine are non-innocuous substrates toward CYP2A6. Therefore, the caffeine test is considered to be a novel and safe method to evaluate CYP2A6 phenotype in humans. In our preliminary studies, the ratio of 17U to 1X in spot urine samples under the dietary caffeine intake could be of use for phenotyping of polymorphic CYP2A6 in Japanese nonsmokers.
The roles of P450s in the 17X 8-hydroxylation (Fig. 1) have not been fully investigated except for two reports with limited information (Gu et al., 1992; Nowell et al., 2002). Gu et al. (1992) examined the isoforms of P450 responsible for the 17X 8-hydroxylation by using lysate fractions prepared from HepG2 cells expressing CYP1A2, CYP2A6, CYP2B6, CYP2E1, CYP3A4, or CYP3A5. They found that CYP1A2 and CYP2A6 catalyzed the 17X 8-hydroxylation at the 17X concentration of 1000 μM and that the activity of CYP2A6 was 1.3 times higher than that of CYP1A2 (Gu et al., 1992). In the present study, we also clarified that CYP1A2 and CYP2A6 expressed in E. coli membranes were involved in the 17X 8-hydroxylation at the 17X concentration of 100 μM (Fig. 4). In contrast to our result, a recent report of Nowell et al. (2002) has shown that CYP2A6 expressed in lymphoblastoid cells catalyzed the 17X 8-hydroxylation at the 17X concentration of 100 μM, whereas CYP1A2 did not. This inconsistency may be attributable to the different expression systems. When we measured the 17X 8-hydroxylase activities using E. coli cells, insect cells or lymphoblastoid cells expressing P450, the 17X 8-hydroxylase activities of CYP1A2 differed from the expression systems, although the activities of CYP2A6 were almost similar to each other (data not shown). CYP2A6 has been considered to be a principal enzyme responsible for the 17X 8-hydroxylation from the study of Gu et al. (1992). However, we cannot totally exclude the possibility that CYP1A2 was also the 17X 8-hydroxylase in humans, because the contents of CYP1A2 protein in human livers were approximately 3-fold higher than those of CYP2A6 (Shimada et al., 1994). Therefore, we used human liver microsomes to clarify the contribution of CYP2A6 to the 17X 8-hydroxylation in the present study.
We revealed that the 17X 8-hydroxylase activities of human liver microsomes significantly correlated with coumarin 7-hydroxylase activities (Fig. 5B), and tranylcypromine inhibited more than 80% of the 17X 8-hydroxylase activities in most of the human liver microsomes. Previous reports have demonstrated that tranylcypromine also inhibits the activity of CYP2C19 other than CYP2A6 (Inaba et al., 1985). However, the 17X 8-hydroxylase activity of CYP2C19 expressed in E. coli membranes was extremely low compared with CYP2A6 (Fig. 3). No significant correlation was found between the 17X 8-hydroxylase activities and S-mephenytoin 4′-hydroxylase (CYP2C19) activities in human liver microsomes (data not shown). Therefore, the inhibitory effects of tranylcypromine on the 17X 8-hydroxylase activities were due to the inhibition of CYP2A6 activities in human liver microsomes. Exceptionally, one liver microsomal sample (sample 16) showed a residual activity of 56% in the presence of tranylcypromine. This result might be attributable to the involvement of CYP1A2 in a part of the 17X 8-hydroxylation, because this sample showed a high activity of CYP1A2 and a low activity of CYP2A6 in the correlation analysis (Fig. 5). The 17X 8-hydroxylase activities of liver microsomes from subjects carrying mutant CYP2A6 genotype were remarkably low (Fig. 7). Particularly, the 17X 8-hydroxylase activities of liver microsomes derived from subjects genotyped as CYP2A6*4/*4 were not detected. Based on these data, we considered that CYP2A6 was the principal enzyme responsible for the 17X 8-hydroxylation in human livers. Nevertheless, we cannot exclude the contribution of CYP1A2 to this reaction, but it was predicted to be small.
In conclusion, we demonstrate that CYP2A6 is the principal 17X 8-hydroxylase in human livers and the genetic polymorphism of the CYP2A6 gene has a substantial influence on the 17X 8-hydroxylation shown in Fig. 1 in vitro and in vivo. It is proposed that caffeine test may be a useful tool for the CYP2A6 phenotyping.
Footnotes
-
This work was supported in part by a Grant-in-Aid (No. 99-2) from the Organization for Pharmaceutical Safety and Research and the Ministry of Education, Science, Sports and Culture of Japan. This work was also supported in part by a Grant-in-Aid for Cancer Research from the Ministry of Health, Labour and Welfare of Japan and a Smoking Research Foundtion Grant for Biomedical Research in Japan.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.105.004796.
-
ABBREVIATIONS: 137X, 1,3,7-trimethylxanthine (caffeine); P450, cytochrome P450; CPR, NADPH-P450 reductase; 17X, 1,7-dimethylxanthine; 1X, 1-methylxanthine; 17U, 1,7-dimethyluric acid; 1U, 1-methyluric acid; AAMU, 5-acetylamino-6-amine-3-methyluracil; AFMU, 5-acetylamino-6-formylamine-3-methyluracil; PM, poor metabolizer.
- Received March 22, 2005.
- Accepted June 22, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}