


Abstract
Because cancer is often treated with combination therapy, unexpected pharmacological effects can occur because of drug–drug interactions. Several drugs are able to cause upregulation or downregulation of drug transporters or cytochrome P450 enzymes, particularly CYP3A4. Induction of CYP3A4 may result in decreased plasma levels and therapeutic efficacy of anticancer drugs. Since the pregnane X receptor (PXR) is one of the major transcriptional regulators of CYP3A4, PXR antagonists can possibly prevent CYP3A4 induction. Currently, a limited number of PXR antagonists are available. Some of these antagonists, such as sulphoraphane and coumestrol, belong to the so-called complementary and alternative medicines (CAM). Therefore, the aim was to determine the potential of selected CAM (β-carotene, Echinacea purpurea, garlic, Ginkgo biloba, ginseng, grape seed, green tea, milk thistle, saw palmetto, valerian, St. John's Wort, and vitamins B6, B12, and C) to inhibit PXR-mediated CYP3A4 induction at the transcriptional level, using a reporter gene assay and a real-time polymerase chain reaction assay in LS180 colon adenocarcinoma cells. Furthermore, computational molecular docking and a LanthaScreen time-resolved fluorescence resonance energy transfer (TR-FRET) PXR competitive binding assay were performed to explore whether the inhibiting CAM components interact with PXR. The results demonstrated that milk thistle is a strong inhibitor of PXR-mediated CYP3A4 induction. The components of milk thistle responsible for this effect were identified as silybin and isosilybin. Furthermore, computational molecular docking revealed a strong interaction between both silybin and isosilybin and PXR, which was confirmed in the TR-FRET PXR assay. In conclusion, silybin and isosilybin might be suitable candidates to design potent PXR antagonists to prevent drug–drug interactions via CYP3A4 in cancer patients.
Introduction
Cancer patients are often treated with multiple anticancer drugs or hormonal agents in combination with supplemental therapies to treat or prevent side effects (Harmsen et al., 2009). However, this treatment is not always successful and often unwanted pharmacological effects occur in cancer patients, such as unresponsiveness or toxicity. A common cause for these unpredictable effects is up and down regulation of drug transporters or drug-metabolizing enzymes such as cytochrome P450 (Harmsen et al., 2009; Chen et al., 2010). Of these enzymes, CYP3A4 is quantitatively the most important because it metabolizes more than 50% of all drugs and the majority of the currently prescribed anticancer drugs (Anzenbacher and Anzenbacherová, 2001).
CYP3A4 is highly inducible by rifampicin and multiple anticancer drugs, such as erlotinib, paclitaxel, docetaxel, ifosfamide, tamoxifen, and flutamide (Harmsen et al., 2009). These drugs elevate the levels of CYP3A4 and promote their own metabolism by CYP3A4, a process called autoinduction (Kerbusch et al., 2000). Furthermore, concomitant use of these drugs can also result in an enhancement of each other’s metabolism. As a result, plasma levels decrease, potentially leading to a reduced therapeutic effect. Consequently, the therapeutic outcome of anticancer therapy can become less predictable (Sparreboom et al., 2004; Tascilar et al., 2006).
One of the major transcriptional regulators of CYP3A4 is the pregnane X receptor (PXR) (Pascussi et al., 2003; Harmsen et al., 2008). After activation of PXR by ligand binding, PXR forms a heterodimer with retinoid X receptor in the nucleus. The heterodimer of PXR with retinoid X receptor binds and activates response elements present in the regulatory region of target genes (Pascussi et al., 2003). When the transcription of target gene CYP3A4 is activated, the level of CYP3A4 expression increases. Because PXR is the major transcriptional regulator of CYP3A4 induction, the hypothesis is that a PXR antagonist could inhibit CYP3A4 induction and increase the exposure of affected anticancer drugs.
The identification of potent PXR antagonists may therefore be important. However, it is difficult to predict interactions between PXR and potential antagonists, because of its flexible ligand binding pocket (Watkins et al., 2003). Consequently, there are a limited number of PXR antagonists available. Nevertheless, the list of PXR antagonists is continuously growing. However, the discovery of more potent and selective PXR antagonists is required (Ekins et al., 2007). Some of the existing PXR antagonists, like sulphoraphane and coumestrol, belong to the so-called complementary and alternative medicines (CAM) (Biswas et al., 2009). Therefore, other CAM may have the potential to inhibit PXR. In addition, it is interesting to screen CAM because these relatively safe supplements are frequently used in combination with chemotherapy by cancer patients (Sparreboom et al., 2004).
The aim of the present study is to identify CAM with inhibiting effects toward PXR-mediated CYP3A4 induction. To address this issue, the following CAM were tested in vitro: β-carotene, Echinacea (Echinacea purpurea), garlic (Allium sativum), green tea (Camellia sinensis), Ginkgo biloba, ginseng (Eleutherococcus senticosus), grape seed (Vitis vinifera), milk thistle (Silybum marianum), saw palmetto (Serenoa serrulata), St. John’s Wort (Hypericum perforatum), valerian (Valeriana officinalis), and vitamins B6, B12, and C. These CAM were selected based on the frequency of use by cancer patients and the potential to interact with anticancer drugs (McCune et al., 2004; Sparreboom et al., 2004; Werneke et al., 2004; Gupta et al., 2005; Tascilar et al., 2006).
Materials and Methods
Reagents and Chemicals
All cell culture media and supplements were purchased from PAA Laboratories (Pasching, Austria). Standardized extracts of milk thistle, Ginkgo biloba, Echinacea, ginseng, St. John’s Wort, saw palmetto, grape seed, green tea, valerian, and vitamin C were obtained from Chromadex (Irvine, CA). Standardized extracts of β-carotene, garlic, and vitamins B6 and B12 were purchased from ABCR GmbH & Co KG (Karlsruhe, Germany), Organic Herb Inc. (Changsha City, China), UPS (Rockville, MD), and Sigma-Aldrich (Zwijndrecht, The Netherlands), respectively. Table 1 provides details of all standardized extracts. Siliphos and Vitotaal (commercial milk thistle products) were purchased from Thorne Research Inc. (Dover, ID) and Santesa B.V. (Weesp, The Netherlands), respectively. Milk thistle’s flavonolignans silybin A+B and flavonoid taxifolin were obtained from Sigma-Aldrich and isosilybin A+B, silydianin, and silychristin from PhytoLab (Vestenbergsgreuth, Germany). Details about these flavonolignans and flavonoid are shown in Table 2. Dimethylsulfoxide (DMSO) was purchased from Acros Organics (Fair Lawn, NJ), rifampicin, hyperforin (dicyclohexylammonium) salt, SR12813 [tetraethyl 2-(3,5-di-tert-butyl-4-hydroxyphenyl)ethenyl-1,1-bisphosphonate], ketoconazole, and paclitaxel from Sigma-Aldrich, and erlotinib from Sequoia Research Products Ltd (Pangbourne, UK). (S)-1-[(1S,3S,4S)-4-[(S)-2-(3-benzyl-2-oxo-imidazolidin-1-yl)-3,3-dimethyl-butyrylamino]-3-hydroxy-5-phenyl-1-(4-pyridin-2-yl-benzyl)-pentylcarbamoyl]-2,2-dimethyl-propyl-carbamic acid methyl ester (A-792611)was a kind gift from Abbott Laboratories (North Chicago, IL) and clotrimazole was purchased from LTK Laboratories Inc. (St. Paul, MN). Corning (Amsterdam, The Netherlands) kindly provided the 384-well Corning plates. The following reagents were purchased from Invitrogen (Blijswijk, The Netherlands) for the LanthaScreen time-resolved fluorescence resonance energy transfer (TR-FRET PXR) competitive binding assay: TR-FRET PXR assay buffer, Fluormone PXR green, dithiothreitol, terbium-labeled anti-glutathione S-transferase antibody, and hPXR-LBD (ligand-binding domain).
Details of the standardized CAM extracts
Details of milk thistle’s flavonolignans and flavonoids
Cell Culture
The human colon adenocarcinoma-derived cell line LS180 was purchased from the American Type Culture Collection (Manassas, VA). This cell line was maintained in Roswell Park Memorial Institute (RPMI) 1640 medium with 25 mM HEPES and ι-glutamine, supplemented with 10% (v/v) heat-inactivated fetal bovine serum, 100 U/ml penicillin, and 100 μg/ml streptomycin (RPMI 1640 complete medium) at 37°C under a humidified atmosphere of 5% CO2.
Plasmids
The pCDG-hPXR expression vector was generously provided by Dr. Ron Evans (The Howard Hughes Medical Institute, La Jolla, CA), the pGL3-CYP3A4-XREM (proximal, −362/+53; distal, −7836/−7208) luciferase reporter construct was provided by Dr. Richard Kim (Vanderbilt University, Nashville, TN), and the pRL-TK control plasmid was obtained from Promega (Madison, WI). Plasmids were checked by enzyme restriction and agarose gel electrophoresis, and purified using Promega’s Pureyield Midi-prep according to the manufacturer’s instructions.
CYP3A4 Reporter Gene Assay
LS180 cells were plated at a density of 5.0 × 104 cells/well in 96-well plates (Greiner Bio-One BV, Alphen aan den Rijn, The Netherlands) in 200 μl RPMI 1640 complete medium and incubated overnight in a 5% CO2-humidified atmosphere at 37°C. After 24 hours of incubation, the cells were transfected with 62.5 ng/well nuclear receptor expression vector (pCDG-hPXR), 175 ng/well CYP3A4 luciferase reporter construct (pGL3-CYP3A4-XREM), and 12.5 ng/well renilla luciferase expression control plasmid (pRL-TK), using 0.8 μl/well nanofectin transfection reagent from PAA Laboratories in 150 mM NaCl. After 24 hours, the medium was removed and the cells were washed with phosphate-buffered saline (37°C). Subsequently, 200 μl/well of fresh medium was added containing 0.3% DMSO or CYP3A4 inducers [10 μM rifampicin, 20 μM erlotinib, or 20 μM paclitaxel (Harmsen et al., 2009)] with or without addition of known PXR antagonists [10 μM ketoconazole and 10 μM A-792611 (Healan-Greenberg et al., 2008)] or 100 μg/ml standardized CAM extracts (β-carotene, Echinacea, garlic, green tea extract, Ginkgo biloba, ginseng, grape seed extract, milk thistle, saw palmetto, St. John’s Wort, valerian, or vitamins B6, B12, and C) for the CAM screen. According to a cell viability assay, these concentrations were not cytotoxic (unpublished data).
For further investigation of milk thistle, standardized milk thistle extract and extracts of the commercial products Siliphos and Vitotaal were tested at concentrations of 50, 75, and 100 μg/ml (see the subsequent section on the preparation of milk thistle extracts). Furthermore, milk thistle’s flavonolignans silybin (mixture of diastereomers A+B 1:1), isosilybin (mixture of diastereomers A+B 1:1), silychristin, silydianin, and flavonoid taxifolin, were evaluated at concentrations of 89, 133, and 200 μM (43, 64, and 96 μg/ml for all components except for taxifolin, which were 27, 41, and 61 μg/ml). To determine the potency of silybin and isosilybin to inhibit CYP3A4 induction, IC50 curves were prepared at a concentration range from 2 to 250 μM and 2 to 200 μM, respectively. The concentrations of the standardized milk thistle extract, both extracts of commercial products and milk thistle’s components, were not cytotoxic according to the performed cell viability assay (unpublished data).
All compounds were dissolved in DMSO and the final solvent concentration did not exceed 0.3%. After 24 hours of exposure, the medium was removed. Subsequently, the cells were washed with phosphate-buffered saline and lysed with 25 μl/well Passive Lysis Buffer (Promega) for 15 minutes on a vortex. The cell lysates (10 μl) were transferred to a white half-area 96-well plate (Corning B.V., Schiphol-Rijk, The Netherlands). The reporter activities of firefly luciferase and renilla luciferase were measured with the Dual-Luciferase Reporter Assay System (Promega) according to the manufacturer’s manual, with reagent volumes adjusted to the cell lysate volume. Luminescence was recorded on a Mithras LB 940 microplate reader (Berthold Technologies, Bad Wildbad, Germany). The firefly luciferase signals were normalized to the renilla luciferase signal. To calculate fold inductions, the ratios of all incubations were compared with normalized signals of the negative control DMSO.
Preparation of Milk Thistle Extracts.
The content of the Siliphos and Vitotaal capsules was suspended in ethanol (10 ml ethanol per gram capsule content) and refluxed for 1 hour. Subsequently, the residues were centrifuged for 10 minutes at 650g. After filtration, the fraction containing milk thistle was evaporated and freeze-dried. The yields are calculated by comparing the mass of the extract with the mass of the capsule content (Table 3). The extracts of Siliphos and Vitotaal were stored at −20°C.
Yield of the commercial milk thistle products
Quantitative Real-Time Polymerase Chain Reaction
LS180 cells were plated at a density of 2.5 × 105 cells/well in 12-well plates (Greiner Bio-One BV) in 1 ml RPMI 1640 complete medium and were incubated for 48 hours in a 5% CO2-humidified atmosphere at 37°C. After 48 hours, this medium was removed and fresh medium was added containing 0.3% DMSO or 10 μM rifampicin (Harmsen et al., 2009) with or without known PXR antagonists (10 μM ketoconazole and 10 μM A-792611) or 50, 75, and 100 μg/ml standardized milk thistle extract. The quantitative real-time polymerase chain reaction (qRT-PCR) experiment was also performed with 100 μM milk thistle’s components silybin A+B, isosilybin A+B, taxifolin, silychristin, and silydianin.
After 24 hours, the cells were washed and total RNA was isolated using the SV Total RNA Isolation System (Promega). The RNA quantity and integrity were determined with a Nanodrop Diode Array Spectrophotometer (Nanodrop 1000; Isogen Life Science, IJsselstein, The Netherlands). To synthesize cDNA, 1 μg RNA was reversed transcribed by using the RevertAid First Strand cDNA Synthesis Kit (Fermentas, St Leon-Rot, Germany), using random priming. Assay-on-Demand PCR primers and Taqman probe (Applied Biosystems, Foster City, CA) were used for 18S (Hs99999901_s1) and human CYP3A4 (Hs00604506_m1) RT-PCR. The reactions were singleplexed with 18S. The CYP3A4 and housekeeping gene 18S mRNA expression levels were analyzed using an ABI prism 7000 sequence detection system (Applied Biosystems). The relative quantification defines the changes in CYP3A4 gene expression between the different incubations. First, the cycle threshold (ct) value and the Δct values (difference in ct values compared with the control such as DMSO) were determined. Subsequently, the 2-(difference in Δct) method was used to determine the relative changes in CYP3A4 gene expression (Livak and Schmittgen, 2001).
Computational Molecular Docking
Dockings were performed with Autodock 4.2 (Scripps Research Institute, La Jolla, CA) (Morris et al., 1998), as part of Yasara 12.4.1. software (Krieger et al., 2004). The structure of the ligand binding domain of hPXR of Watkins 2003 was used [Protein Data Bank ID 1M13 (Watkins et al., 2003)]. Docking of hyperforin was started from its X-ray position in 1M13. Subsequently, docking of rifampicin was started after superimposing the hPXR-rifampicin complex 1SKX (Chrencik et al., 2005) onto 1M13. The PXR antagonist A-792611 was built from the similar HIV-protease inhibitor in Protein Data Bank ID 3GGV (Degoey et al., 2009), guided by the structure drawing from Healan-Greenberg et al. (2008). The other compounds (ketoconazole, paclitaxel, erlotinib, and milk thistle’s components) were obtained as Sybyl Mol2 files from ChemIDplus Advanced (http://chem.sis.nlm.nih.gov/chemidplus/; National Institutes of Health, Bethesda, MD). The milk thistle’s components silybin A+B, isosilybin A+B, silychristin, and silydianin were modified to follow the stereochemistry as given by Lee et al. (2007), and roughly placed in the hPXR pocket.
Yasara was used for cleaning the molecules with regard to atom types and bond types. After initial minimization and stimulated annealing with the Amber03 force field (Duan et al., 2003) of the ligand and the sidechains of the hPXR residues within 7 Å, 250 rigid docking runs were performed, with ga_pop_size set at 15000.
LanthaScreen TR-FRET PXR Competitive Binding Assay
A LanthaScreen TR-FRET PXR competitive binding assay was conducted according to the manufacturer’s (Invitrogen) protocol. In brief, the assay was performed in black noncoated, low-volume and round-bottomed 384-well Corning plates. The compounds were diluted in TR-FRET PXR assay buffer and 10 μl/well of these dilutions were dispensed in quaduplicate with the following end concentrations: 100 μg/ml milk thistle and 100 μM of silybin, isosilybin, silydianin, silychristin and taxifolin. In addition, 100 μM of the positive controls ketoconazole and A-792611 were tested, accompanied by known PXR agonists 100 μM rifampicin, 100 μM clotrimazole, 100 μM hyperforin, and 1 μM SR12813. Second, 5 μl/well of Fluormone PXR green at a final concentration of 40 nM was added. At last, 5 μl/well of the following mix was added: dithiothreitol, terbium-labeled anti-glutathione S-transferase antibody, and hPXR-LBD at final concentrations of 50 μM, 10 nM, and 5 nM, respectively. The plate was incubated for 1 hour at room temperature, protected from light. The PerkinElmer Envision plate reader was used to measure TR-FRET (PerkinElmer, Waltham, MA). Table 4 gives an overview of the applied settings. For the TR-FRET ratios, the emission signal at 520 nm was divided by the emission signal at 495 nm.
Settings of the PerkinElmer Envision plate reader
Statistical Analysis
Data were analyzed by using the two-sample t test and were considered statistically significant when P < 0.05. All statistical determinations were performed with SPSS software (version 16.0; SPSS Inc., Chicago, IL).
Results
CAM Screen
The potential inhibition of PXR-mediated CYP3A4 induction by 14 of the most commonly used CAM (β-carotene, Echinacea, garlic, green tea, Ginkgo biloba, ginseng, grape seed, milk thistle, saw palmetto, St. John’s Wort, valerian, and vitamins B6, B12, and C) (McCune et al., 2004; Sparreboom et al., 2004; Werneke et al., 2004; Gupta et al., 2005; Tascilar et al., 2006) was determined in LS180 cells, transfected with plasmids expressing hPXR, CYP3A4, and Renilla. As shown in Fig. 1, 100 μg/ml milk thistle extract is the strongest inhibitor of PXR-mediated CYP3A4 induction by rifampicin, because this CAM significantly inhibited CYP3A4 induction by 83%. Furthermore, extracts of Echinacea and vitamin B12 moderately inhibited this induction by 24 and 18%, respectively. Compared with the 32 and 49% inhibition of CYP3A4 induction by the well-known PXR antagonists ketoconazole and A-792611, the degree of inhibition of CYP3A4 induction is high by milk thistle and is moderate by Echinacea and vitamin B12.

CAM screen for possible inhibition of PXR-mediated CYP3A4 induction by rifampicin using reporter gene. LS180 cells were transfected with the pCDG-hPXR expression vector, pGL3-CYP3A4-XREM reporter construct, and pRL-TK control plasmid. After 24 hours of transfection, the LS180 cells were incubated with 0.3% DMSO or the CYP3A4 model inducer rifampicin (10 µM) in combination with the negative control DMSO, CAM, or the positive controls ketoconazole and A-792611. The luciferase activities were measured after 24 hours of incubation. Data are means ± S.D. from a representative experiment performed in triplicate. The fold inductions compared with 0.3% DMSO are shown (comparison with the fold induction by rifampicin: *P < 0.05; **P < 0.01). DMSO +, results after incubation without rifampicin; Rif +, results after incubation with rifampicin in combination with DMSO, CAM, or positive controls.
Standardized Milk Thistle Extract
Since 100 μg/ml standardized milk thistle extract was shown to be a strong inhibitor of PXR-mediated CYP3A4 induction, milk thistle was further investigated by incubating transfected LS180 cells with three concentrations of milk thistle: 50, 75, and 100 μg/ml. In addition, besides the model inducer rifampicin also the anticancer drugs erlotinib and paclitaxel were used as inducers to determine whether milk thistle can also inhibit CYP3A4 induction caused by anticancer drugs.
CYP3A4 Reporter Gene Assay.
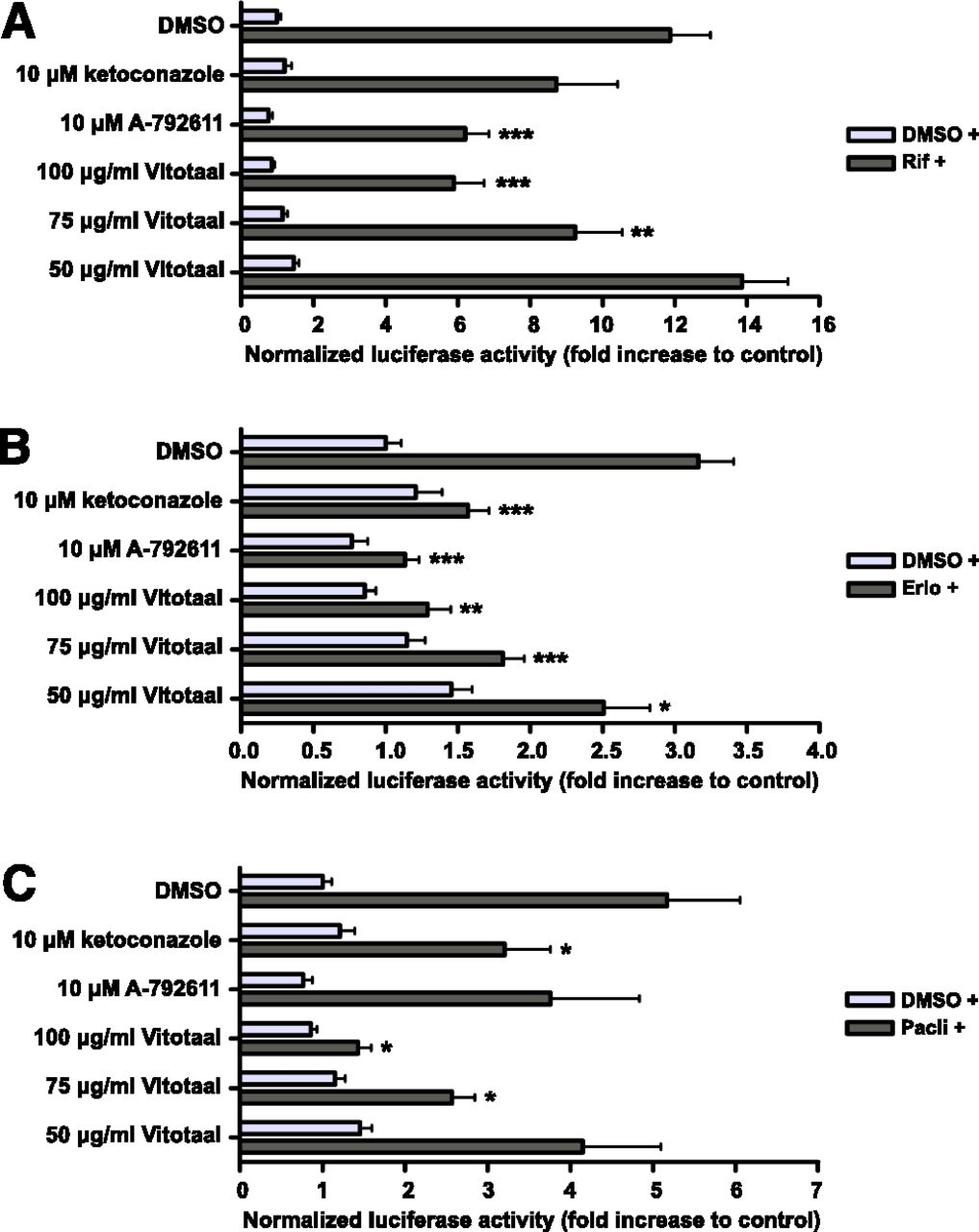
A solution of 75 and 100 μg/ml standardized milk thistle extract significantly inhibited rifampicin-mediated CYP3A4 induction by 38 and 74%, respectively (Fig. 2A). At concentrations of 75 and 100 μg/ml, the standardized milk thistle extract also significantly inhibited erlotinib-mediated CYP3A4 induction by 66 and 79%, respectively, and paclitaxel-mediated CYP3A4 induction by 62 and 83% (Fig. 2, B and C). Compared with ketoconazole and A-792611, milk thistle is a strong inhibitor of CYP3A4 induction caused by rifampicin and both anticancer drugs.

Inhibition of PXR-mediated CYP3A4 induction by the standardized milk thistle extract using reporter gene. Transfected LS180 cells were incubated with 0.3% DMSO or the CYP3A4 inducers rifampicin (10 µM), erlotinib (20 μM), or paclitaxel (20 μM) in combination with the negative control DMSO, the standardized milk thistle extract, or the positive controls ketoconazole and A-792611. The luciferase activities were measured after 24 hours of incubation. Data are means ± S.D. from a representative experiment performed in triplicate. The fold inductions compared with 0.3% DMSO are shown (comparison with the fold induction by the inducer: *P < 0.05; **P < 0.01; ***P < 0.001). CYP3A4 induction by rifampicin (A), erlotinib (B), and paclitaxel (C). DMSO +, results after incubation without CYP3A4 inducer; Rif +, Erlo + or Pacli +, results after incubation with rifampicin, erlotinib or paclitaxel in combination with DMSO, milk thistle or positive controls.
CYP3A4 qRT-PCR Assay.
The potential of milk thistle to inhibit PXR-mediated CYP3A4 induction was also investigated in nontransfected LS180 cells at the mRNA expression level, using qRT-PCR analysis.
A solution of 100 μg/ml standardized milk thistle extract significantly inhibited rifampicin-mediated CYP3A4 induction by 80% (Fig. 3). These results are in accordance with the inhibition of PXR-mediated CYP3A4 induction by the standardized milk thistle extract, determined using the CYP3A4 reporter gene assay (Fig. 2).

Inhibition of PXR-mediated CYP3A4 induction by the standardized milk thistle extract using qRT-PCR. LS180 cells were incubated with 0.3% DMSO or the CYP3A4 model inducer rifampicin (10 µM) in combination with the negative control DMSO, the standardized milk thistle extract, or the positive controls ketoconazole and A-792611. The mRNA expression levels were measured after 24 hours of incubation using singleplexed qRT-PCR. Data are means ± S.D. from a representative experiment performed in triplicate, with the exception of the incubation with ketoconazole combined with rifampicin. The fold inductions compared with 0.3% DMSO are shown (comparison with the fold induction by rifampicin: *P < 0.05). DMSO +, results after incubation without rifampicin; Rif +, results after incubation with rifampicin in combination with DMSO, milk thistle, or positive controls.
Commercial Milk Thistle Products: Vitotaal and Siliphos
The standardized milk thistle extract was shown to be a strong inhibitor of rifampicin-, erlotinib-, and paclitaxel-mediated CYP3A4 induction. However, cancer patients do not use this formulation but a variety of commercial milk thistle products. Regarding the differences in the composition between standardized extracts and extracts of commercial milk thistle products, it is important to determine the effect of commercial milk thistle on CYP3A4 induction.
CYP3A4 (Reporter Gene Assay).
The commercial product Vitotaal also significantly inhibited rifampicin-mediated CYP3A4 induction by 22 and 50% at concentrations of 75 and 100 μg/ml, respectively (Fig. 4A). These concentrations of Vitotaal also resulted in a significant inhibition of 43 and 59% of CYP3A4 induction by erlotinib (Fig. 4B). Furthermore, Vitotaal significantly inhibited paclitaxel-mediated CYP3A4 induction by 50 and 75% at concentrations of 75 and 100 μg/ml, respectively (Fig. 4C).

Inhibition of PXR-mediated CYP3A4 induction by Vitotaal using reporter gene. Transfected LS180 cells were incubated with 0.3% DMSO or the CYP3A4 inducers rifampicin (10 µM), erlotinib (20 μM), or paclitaxel (20 μM) in combination with the negative control DMSO, Vitotaal, or the positive controls ketoconazole and A-792611. The luciferase activities were measured after 24 hours of incubation. Data are means ± S.D. from a representative experiment performed in triplicate. The fold inductions compared with 0.3% DMSO are shown (comparison with the fold induction by the inducer: *P < 0.05; **P < 0.01; ***P < 0.001). CYP3A4 induction by rifampicin (A), erlotinib (B), and paclitaxel (C). DMSO +, results after incubation without CYP3A4 inducer; Rif +, Erlo + or Pacli +, results after incubation with rifampicin, erlotinib or paclitaxel in combination with DMSO, Vitotaal or positive controls.
A solution of 100 μg/ml Siliphos significantly inhibited rifampicin-, erlotinib- and paclitaxel-mediated CYP3A4 induction by 29, 50, and 50%, respectively (Fig. 5). In addition, Siliphos significantly inhibited erlotinib-mediated CYP3A4 induction by 34% at a concentration of 75 μg/ml.

Inhibition of PXR-mediated CYP3A4 induction by Siliphos using reporter gene. Transfected LS180 cells were incubated with 0.3% DMSO or the CYP3A4 inducers rifampicin (10 µM), erlotinib (20 μM), or paclitaxel (20 μM) in combination with the negative control DMSO, Siliphos, or the positive controls ketoconazole and A-792611. The luciferase activities were measured after 24 hours of incubation. Data are means ± S.D. from a representative experiment performed in triplicate. The fold inductions compared with 0.3% DMSO are shown (comparison with the fold induction by the inducer: *P < 0.05; **P < 0.01; ***P < 0.001). CYP3A4 induction by rifampicin (A), erlotinib (B), and paclitaxel (C). DMSO +, results after incubation without CYP3A4 inducer; Rif +, Erlo + or Pacli +, results after incubation with rifampicin, erlotinib or paclitaxel in combination with DMSO, Siliphos or positive controls.
CYP3A4 qRT-PCR Assay.
The potential of both commercial milk thistle products to inhibit PXR-mediated CYP3A4 induction was also confirmed in nontransfected LS180 cells, using qRT-PCR analysis.
Both Vitotaal and Siliphos significantly inhibited rifampicin-mediated CYP3A4 induction by 69 and 61% at a concentration of 100 μg/ml respectively, in accordance with the results of the reporter gene assay (Fig. 6).

Inhibition of PXR-mediated CYP3A4 induction by Vitotaal and Siliphos using qRT-PCR. LS180 cells were incubated with 0.3% DMSO or the CYP3A4 model inducer rifampicin (10 µM) in combination with the negative control DMSO, Vitotaal, Siliphos, or the positive controls ketoconazole and A-792611. The mRNA expression levels were measured after 24 hours of incubation using singleplexed qRT-PCR. Data are means ± S.D. from a representative experiment performed in triplicate, with exception of the incubation with ketoconazole combined with rifampicin. The fold inductions compared with 0.3% DMSO are shown (comparison with the fold induction by rifampicin: *P < 0.05). Inhibition of CYP3A4 induction by Vitotaal (A) and Siliphos (B). DMSO +, results after incubation without rifampicin; Rif +, results after incubation with rifampicin in combination with DMSO, Vitotaal, Siliphos, or positive controls.
Milk Thistle’s Components
Overall, the inhibition of PXR-mediated CYP3A4 induction by the standardized milk thistle extract, Vitotaal, and Siliphos is confirmed in two assays: reporter gene and qRT-PCR. However, milk thistle is a mixture of multiple components, including silybin (mixture of diastereomers A+B 1:1), isosilybin (mixture of diastereomers A+B 1:1), taxifolin, silychristin, and silydianin. To identify a potent inhibitor of PXR-mediated CYP3A4 induction, it is important to identify which component is responsible for the observed effect. Therefore, all of milk thistle’s components were investigated in both the reporter gene and qRT-PCR assays.
CYP3A4 (Reporter Gene Assay).
The results of the reporter gene assay showed that milk thistle’s components silybin and isosilybin are responsible for the inhibition of PXR-mediated CYP3A4 induction by milk thistle. Silybin significantly inhibited rifampicin-mediated CYP3A4 induction by 58 and 81% at concentrations of 133 and 200 μM, respectively (Fig. 7A). Compared with silybin, its isomer isosilybin is a stronger inhibitor of PXR-mediated CYP3A4 induction. A solution of 89, 133, and 200 μM isosilybin significantly inhibited CYP3A4 induction by 64, 82, and 88%, respectively (Fig. 7B).

Inhibition of PXR-mediated CYP3A4 induction by milk thistle’s components using reporter gene. Transfected LS180 cells were incubated with 0.3% DMSO or the CYP3A4 model inducer rifampicin (10 µM) in combination with the negative control DMSO, milk thistle’s components, or the positive controls ketoconazole and A-792611. The luciferase activities were measured after 24 hours of incubation. Data are means ± S.D. from a representative experiment performed in triplicate. The fold inductions compared with 0.3% DMSO are shown (comparison with the fold induction by rifampicin: **P < 0.01; ***P < 0.001). (A) silybin (A+B), (B) isosilybin (A+B), (C) taxifolin, (D) silychristin, and (E) silydianin. DMSO +, results after incubation without CYP3A4 inducer; Inducer +, results after incubation with CYP3A4 inducer in combination with DMSO, milk thistle, or positive controls; Rif +, results after incubation with rifampicin in combination with DMSO, milk thistle's components, or positive controls.
Compared with the potent inhibition of rifampicin-mediated CYP3A4 by silybin and isosilybin, the inhibition by 200 μM taxifolin and silychristin was moderate (Fig. 7, C and D). Silydianin showed no inhibition of rifampicin-mediated CYP3A4 induction (Fig. 7E).
In addition, IC50 curves of silybin and isosilybin reveal the potency of these components to inhibit CYP3A4 induction (Fig. 8). The IC50 value of silybin is 135 μM and isosilybin has an IC50 value of 74 μM, which confirms that isosilybin is more potent.

IC50 curves of silybin and isosilybin using reporter gene. Transfected LS180 cells were incubated with CYP3A4 model inducer rifampicin (10 µM) in combination with 2–250 µM silybin and 2–200 µM isosilybin. The luciferase activities were measured after 24 hours of incubation. Data are means ± S.D. from a representative experiment performed in triplicate. The fold inductions compared with 0.3% DMSO are shown.
CYP3A4 qRT-PCR Assay.
The potential of silybin and isosilybin to inhibit PXR-mediated CYP3A4 induction is also confirmed in nontransfected LS180 cells using qRT-PCR analysis. Silybin and isosilybin significantly inhibited PXR-mediated CYP3A4 induction with 43 and 55% at a concentration of 100 μM, respectively. The other components did not inhibit PXR-mediated CYP3A4 induction (Fig. 9).

Inhibition of PXR-mediated CYP3A4 induction by milk thistle’s components using qRT-PCR. LS180 cells were incubated with 0.3% DMSO or the CYP3A4 model inducer rifampicin (10 µM) in combination with the negative control DMSO, the standardized milk thistle extract, silybin A+B, isosilybin A+B, taxifolin, silychristin, silydianin, or the positive controls ketoconazole and A-792611. The mRNA expression levels were measured after 24 hours of incubation using singleplexed qRT-PCR. Data are means ± S.D. from a representative experiment performed in triplicate. The fold inductions compared with 0.3% DMSO are shown (comparison with the fold induction by rifampicin: *P < 0.05; **P < 0.01). DMSO +, results after incubation without rifampicin; Rif +, results after incubation with rifampicin in combination with DMSO, milk thistle, milk thistle's components, or positive controls.
Together, the results confirm that silybin and isosilybin are responsible for the inhibition of PXR-mediated CYP3A4 induction. Besides the use of a reporter gene assay, which is generally accepted to screen for PXR antagonists, the capability of silybin and isosilybin to bind in the PXR pocket was further investigated via computational molecular docking and a TR-FRET PXR competitive binding assay.
Computational Molecular Docking
For the computational molecular docking, the following compounds were tested: known PXR agonists, known PXR antagonists, and the milk thistle’s components. Although PXR is promiscuous, it is able to bind with high to very high affinity to chemically diverse ligands (Watkins et al., 2001).
PXR agonists rifampicin, paclitaxel, and erlotinib showed a different degree of binding affinity toward the PXR pocket. Rifampicin had the highest binding affinity (1.39 pM), followed by paclitaxel (20.7 pM) and erlotinib (3.97 µM) (Table 5). These results are in accordance with the degree of PXR-mediated CYP3A4 induction from the reporter gene assay. Rifampicin-mediated CYP3A4 induction ranged from 9- to 12-fold, paclitaxel-mediated induction was approximately 5-fold, and erlotinib-mediated induction was approximately 3-fold. For the moderately binding agent erlotinib, more extensive docking (Lamarckian genetic algorithm) was used to check for missed poses. This did not lead to a higher affinity.
Binding affinity of different compounds to hPXR and the involvement of amino acid residues
Hyperforin (the active component of St. John’s Wort) is a well-known CAM component that activates PXR, with a predicted binding affinity of 1.03 nM. The reporter gene assay demonstrated the activation of PXR by St. John’s Wort with an 8-fold CYP3A4 induction (Fig. 1).
Known PXR antagonists ketoconazole and A-792611 also differed in their degree of binding affinity toward PXR. Ketoconazole had a lower binding affinity (2.6 nM) than A-792611 (26.45 pM), which is in accordance with the results of the reporter gene assay and qRT-PCR analysis.
The computational molecular docking of milk thistle’s components showed that silybin and isosilybin have the highest binding affinity, followed by silydianin, silychristin, and taxifolin, which is partially in accordance with the results of the reporter gene assay. In the reporter gene assay, silybin and isosilybin were the strongest antagonists and the other components did not convincingly inhibit CYP3A4 induction. For taxifolin, extensive docking did not improve its moderate PXR affinity.
Based on the computational molecular docking, it is difficult to distinguish PXR agonists and antagonists. However, the pharmacophore of novel PXR antagonists (e.g., A-792611) (Healan-Greenberg et al., 2008) suggested that PXR antagonists bind the small binding site close to the outer surface of PXR at the AF-2 (activation function 2) domain, whereas PXR agonists bind in the core of the ligand binding domain. Binding of steroid receptor coactivator-1 to AF-2 is necessary to stabilize PXR and this can be interfered with PXR antagonists (Ekins et al., 2007). The amino acid Phe429A is located in the AF-2 domain of PXR and it is likely that interaction of steroid receptor coactivator-1 with this amino acid stabilizes the active form of PXR during upregulation (Xue et al., 2007).
According to Table 5 and Fig. 10, the known PXR antagonists ketoconazole and A-792611 interact with this particular amino acid. Interestingly, silybin and isosilybin also interact with Phe429A. Based on the similar binding of both milk thistle’s components and the known PXR antagonists, it is plausible that silybin and isosilybin are potent PXR antagonists. This was already demonstrated in the reporter gene and qRT-PCR assays, in which CYP3A4 induction was strongly inhibited by silybin and isosilybin. Both silydianin and taxifolin did not interact with Phe429A and together with the results of the reporter gene assay, these components are probably no PXR antagonists. However, silychristin showed an interaction with Phe429A, but this component has a lower binding affinity toward PXR. Together with the reporter gene results, silychristin is probably a weak PXR antagonist.

Computational molecular docking of known PXR antagonists (ketoconazole and A-792611) and silybin A and isosilybin to the ligand binding domain of PXR. The key amino acids that are involved in the hydrophobic and polar interactions with ketoconazole, A-792611, Silybin, and isosilybin are shown. The figures are generated with http://poseview.zbh.uni-hamburg.de/.
Overall, although the binding affinities predicted by docking have a standard deviation of around 1.5 log units, it is possible to conclude that at least six compounds from milk thistle have affinities for hPXR comparable to ketoconazole. Of all of milk thistle’s components, silybin and isosilybin are probably the most potent PXR antagonists by interfering with the AF-2 domain.
Lanthascreen TR-FRET PXR Competitive Binding Assay
To confirm binding affinities of silybin and isosilybin toward PXR, the direct interaction with PXR was determined with a TR-FRET PXR competitive binding assay. The results of the TR-FRET PXR competitive binding assay are shown in Fig. 11. The TR-FRET emission ratio was reduced for 44% by 100 μg/ml milk thistle, compared with 24 and 27% by the positive controls 100 μM ketoconazole and A-792611, respectively. Of all of milk thistle’s components, both silybin and isosilybin significantly decreased the TR-FRET ratio with 27%. Silychristin and silydianin moderately affected the ratio with 17 and 9%. Taxifolin did not decrease the TR-FRET ratio.

Competitive binding to PXR-LBD in a TR-FRET PXR competitive binding assay. Human PXR-LBD (5 nM) was incubated for 1 hour with Fluormone PXR Green (40 nM) in the presence of DMSO (1% v/v), SR12813 (1 µM), clotrimazole (100 µM), hyperforin (100 µM), ketoconazole (100 µM), A-792611 (100 µM), milk thistle (100 µg/ml), isosilybin A+B (100 µM), silybin A+B (100 µM), taxifolin (100 µM), silychristin (100 µM), and silydianin (100 µM). After 1 hour, TR-FRET was measuring using the PerkinElmer Envision plate reader. The TR-FRET ratio was calculated by dividing the response at 520 nm by the response at 495 nm. Data are means ± S.D. from a representative experiment performed in quadruplicate. The percentage of the mean values compared with 1% DMSO are shown (*P < 0.05; **P < 0.01).
In comparison, the known PXR agonists SR12813, clotrimazole, and hyperforin decreased the TR-FRET emission ratio by 33, 56, and 47%, respectively. It is important to note that the TR-FRET assay was not applicable for rifampicin, because this compound quenched the fluorescence (unpublished data). This problem was also previously described by Shukla et al. (2009) and Lau et al. (2012).
Discussion
Fourteen of the most commonly used CAM (McCune et al., 2004; Sparreboom et al., 2004; Werneke et al., 2004; Gupta et al., 2005; Tascilar et al., 2006) were screened for their ability to inhibit PXR-mediated CYP3A4 induction by rifampicin, erlotinib, and paclitaxel in LS180 cells.
LS180 cells were used in the present study because it was previously demonstrated that these cells are a more suitable cell line to study the induction of CYP3A4 compared with the more frequently used cell line HepG2 (Harmsen et al., 2008). Of all tested CAM, Echinacea and vitamin B12 were shown to moderately inhibit PXR-mediated CYP3A4 induction by rifampicin. Milk thistle was shown to strongly inhibit CYP3A4 induction in the reporter gene assay. In addition to the standardized milk thistle extract, the extracts of the commercial products Vitotaal and Siliphos were investigated. Both the standardized milk thistle extract and the two extracts of the commercial products significantly inhibited PXR-mediated CYP3A4 induction by rifampicin, erlotinib, and paclitaxel at the transcriptional level. However, compared with the standardized milk thistle extract, the extracts of Vitotaal and Siliphos inhibited PXR-mediated CYP3A4 induction to a lesser extent. A plausible explanation for the difference in the degree of inhibition is the unknown compositions and the variability in the content of milk thistle’s components: seven primary flavonolignans (silybin A, silybin B, isosilybin A, isosilybin B, silychristin, isosilychristin, silydianin) and a flavonoid taxifolin (Kroll et al., 2007). Furthermore, the same weight of the standardized milk thistle extract and the extracts of Vitotaal and Siliphos do not contain the same number of milk thistle’s components because the reprocessing methods differ.
Furthermore, all milk thistle products showed a limited inhibition of CYP3A4 transcription in LS180 cells without the concomitant addition of an inducer. A plausible explanation is the presence of basal activity of PXR in LS180 cells, which can also be inhibited by milk thistle.
Since milk thistle is a mixture of multiple components, it is necessary to identify the component(s) responsible for the inhibition of CYP3A4 induction in the search for new PXR antagonists. Therefore, all components of milk thistle were investigated in the reporter gene assay. Results showed that silybin and isosilybin strongly inhibited rifampicin-mediated CYP3A4 induction.
The inhibition of PXR-mediated CYP3A4 induction by the standardized milk thistle extract, extracts of commercial milk thistle products, and their components was not only determined using a CYP3A4 reporter gene assay, but was also determined in a nontransfected cell system using qRT-PCR. In accordance with the results obtained with the reporter gene assay, the milk thistle extract, both commercial extracts, silybin, and isosilybin significantly inhibited the induction of CYP3A4 mRNA expression by rifampicin as well.
The presented results demonstrate that milk thistle is able to prevent CYP3A4 induction and that silybin and isosilybin are responsible for this effect. It is plausible that the inhibition of CYP3A4 induction occurs via the PXR pathway, because PXR is highly expressed in the transfected LS180 cells during the reporter gene assay and because this assay is generally accepted to screen for PXR antagonists (Luo et al., 2002). However, it is not completely certain that milk thistle–mediated inhibition of CYP3A4 induction is only PXR mediated. When the cells are exposed to milk thistle, CYP3A4 induction could also be inhibited via other pathways. Therefore, additional computational molecular docking of milk thistle’s components and a TR-FRET PXR competitive binding assay were performed to confirm that the inhibition of CYP3A4 induction by milk thistle could be PXR mediated. The results showed that silybin and isosilybin have a high binding affinity toward PXR and both directly interact with PXR. More importantly, in the computational molecular docking, both components showed to interact with the amino acid Phe429A in the AF-2 site similar to the known PXR antagonists ketoconazole and A-792611. Therefore, it is plausible that silybin and isosilybin are PXR antagonists, which is in accordance with the results from the reporter gene assay and qRT-PCR. Silybin and isosilybin can probably also inhibit the PXR-mediated induction of other cytochrome P450 enzymes and drug transporters (e.g. P-glycoprotein). Further investigation is needed to confirm this.
Based on the structure of the active components silybin and isosilybin, a potent PXR-inhibiting agent can be designed to prevent induction of CYP3A4 in cancer patients. Designing a potent (iso)silybin-derived PXR antagonist may be important because of the low bioavailability of milk thistle. In a clinical study with six cancer patients, a 14-day supplementation of 200 mg milk thistle extract resulted in low plasma levels of silybin ranging between 0.0249 and 0.257 μM (van Erp et al., 2005). The use of a hydrophobic silybin-phosphatidylcholine complex (silipide), however, resulted in higher plasma levels of silybin in healthy volunteers (0.29–0.53 μM) (Gatti and Perucca, 1994). When a potent PXR-inhibiting agent is designed with a higher bioavailability, it can possibly reach plasma levels of the determined IC50 values of 74 and 135 μM to prevent CYP3A4 induction by anticancer drugs in patients. In cases in which this (iso)silybin-derived agent inhibits CYP3A4 induction in patients, decreased plasma levels of anticancer drugs can be prevented. However, this should be evaluated in the clinic.
In conclusion, milk thistle’s components silybin and isosilybin prevent PXR-mediated CYP3A4 induction at the transcriptional level, as demonstrated by reporter gene and qRT-PCR assays. Both silybin and isosilybin have a high binding affinity toward PXR according to computational molecular docking and a TR-FRET PXR competitive binding assay. Thus, silybin and isosilybin might be suitable candidates to design potent PXR antagonists to prevent drug–drug interactions via CYP3A4 in cancer patients or patients that use other drugs metabolized by CYP3A4.
Acknowledgments
The authors thank Pasi Halonen, Ben Morris and Boris Rodenko from the NKI Robotics and Screening Center for their technical support with the Lanthascreen experiments.
Authorship Contributions
Participated in research design: Mooiman, Maas-Bakker, Meijerman.
Conducted experiments: Mooiman, Maas-Bakker.
Performed data analysis: Mooiman, Moret.
Wrote or contributed to the writing of the manuscript: Mooiman, Maas-Bakker, Moret, Beijnen, Schellens, Meijerman.
Footnotes
- Received January 11, 2013.
- Accepted May 14, 2013.
This research was supported by the Dutch Cancer Society [Grant UU 2007-3795].
Abbreviations
- A-792611
- (S)-1-[(1S,3S,4S)-4-[(S)-2-(3-benzyl-2-oxo-imidazolidin-1-yl)-3,3-dimethyl-butyrylamino]-3-hydroxy-5-phenyl-1-(4-pyridin-2-yl-benzyl)-pentylcarbamoyl]-2,2-dimethyl-propyl-carbamic acid methyl ester
- AF-2
- activation function 2
- CAM
- complementary and alternative medicine
- ct
- cycle threshold
- DMSO
- dimethylsulfoxide
- PXR
- pregnane X receptor
- qRT-PCR
- quantitative real-time polymerase chain reaction
- SR12813
- [tetraethyl 2-(3,5-di-tert-butyl-4-hydroxyphenyl)ethenyl-1,1-bisphosphonate]
- TR-FRET
- time-resolved fluorescence resonance energy transfer
- Copyright © 2013 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}