


Abstract
The molecular basis of perinatal changes occurring in major UDP-glucuronosyltransferase (UGT) family 1 isoforms and in UGT2B1, a relevant isoform belonging to family 2, was analyzed in rat liver. Nonpregnant, pregnant (19–20 days of pregnancy), and two groups of postpartum animals corresponding to early and middle stages of lactation (2–4 and 10–12 days after delivery, respectively) were studied. UGT activity determined in UDP-N-acetylglucosamine-activated microsomes revealed that bilirubin, p-nitrophenol, and ethynylestradiol (17β-OH and 3-OH) but not androsterone and estrone glucuronidation rates, were decreased in pregnant rats. Decreased enzyme activities returned to control values after delivery.p-Nitrophenol, androsterone, and estrone conjugation rate increased in postpartum rats. Western blot analysis performed with anti-peptide-specific (anti-1A1, 1A5, 1A6, and 2B1) antibodies revealed decreased levels of all family 1 isoforms and UGT2B1 during pregnancy. In postpartum animals, protein level recovered (1A5 and 2B1) or even increased (1A1 and 1A6) with respect to control rats. Northern blot analysis suggested that expression of UGT proteins is down-regulated at a post-translational level during pregnancy and that increased levels of 1A1 and 1A6 observed in postpartum rats were associated to increased mRNA. To establish whether prolactin is involved in up-regulation of UGT1A1 and 1A6 postpartum, ovariectomized rats were treated with 300 μg of ovine prolactin per day for 7 days. The data indicated that prolactin was able to increase expression of UGT1A6 (protein and mRNA) but not 1A1. Thus, prolactin is the likely mediator of the increased expression of UGT1A6 observed in maternal liver postpartum.
UDP-glucuronosyltransferases (UGTs) represent a superfamily of enzymes that catalyze the conjugation of glucuronic acid to both endogenous compounds, including bilirubin, bile acids, and steroid and thyroid hormones, and exogenous compounds, including food additives, drugs, and environmental pollutants (for review, see Tephly and Burchell, 1990). Based on the nucleotide and amino acid sequences, UGT isoforms in mammals are grouped into two major families termed 1 and 2. Enzymes belonging to family 1, including a bilirubin cluster (UGT1A1 and UGT1A5) and a phenol cluster (UGT1A6 and UGT1A7), are formed by alternative splicing of an isoform-specific exon encoding a unique N-terminal region with common exons 2 to 5 encoding an identical C-terminal region. In contrast, UGT family 2 isoforms are each derived from an individual gene. The activity of UGT is affected by many factors, including enzyme inducers, aging, diet, diseases, and hormones (for review, see Tephly and Burchell, 1990;Miners and Mackenzie, 1991; Burchell et al., 1994). Regulation may occur either at the gene transcription level, resulting in changes in mRNA and protein levels, or at the level of post-translational processing. Because of association of UGT with the lipid environment (Zakim and Dannenberg, 1992), restriction of the cosubstrate UDP-glucuronic acid (UDPGA) to access the enzyme active site (Berg et al., 1995; Bossuyt and Blanckaert, 1995) and protein-protein interactions derived from oligomer formation (Peters et al., 1984;Ikushiro et al., 1997; Meech and Mackenzie, 1997), enzyme activity toward a specific substrate may also depend on the functional state of UGT.
Many studies have been conducted to determine UGT activity in experimental animals during pregnancy. This subject is of particular interest because changes in metabolizing enzyme activities in maternal liver may change the risk of exposure by the fetus. During pregnancy, significant hormonal changes take place, mainly in sex steroids levels, that may substantially affect disposition of potentially toxic compounds. Most of the studies performed in rats reported a decrease in liver UGT activity (expressed per milligram of microsomal protein) compared with nonpregnant controls, affecting major family 1 substrates such as bilirubin and phenol derivatives and some family 2 substrates such as estradiol (Halac and Sicignano, 1969; Neale and Parke, 1973;Vore and Soliven, 1979; Muraca et al., 1984; Borlakoglu et al., 1993). It is reasonable to speculate that factors, such as those modulating the functional state of UGT, may be affected in such a way that they similarly decrease the activity of all the isoforms tested. We previously reported that substantial changes in microsomal lipid composition and membrane fluidity occur perinatally but, in contrast to what is expected, they positively modulate UGT activity (Luquita et al., 1994). Consequently, a decrease in the expression of UGTs is more likely involved in down-regulation of UGT activities during pregnancy. At present, no studies have been conducted to clarify the molecular basis of UGT regulation during pregnancy.
We reported that hepatic UGT activity toward planar phenols is increased in female rats during lactation, particularly at the late stage of lactation (19–21 days after delivery) (Luquita et al., 1994). Using a polyclonal nonspecific antibody developed against a phenol-conjugating isoform, we detected an increased content of UGT protein in ovariectomized rats in response to ovine prolactin treatment, suggesting that this hormone may be involved in the regulation of UGT expression postpartum (Luquita et al., 1996). We postulated that UGT induction, together with an increased bile secretory function in lactating rats (Liu et al., 1992), may be an adaptive response to increase the biliary excretion of toxic compounds, thus preventing their secretion into breast milk. Taken together, the evidence suggests a differential regulation of UGT in pregnancy and postpartum. Immediately after delivery, levels of progesterone and estrogens are dramatically decreased, and other hormones, i.e., prolactin, are greatly increased. In consequence, differences in UGT activities in maternal liver between pregnant and lactating rats are most reasonably associated with the various hormonal actions. The activity and protein expression of the different UGT isoforms, particularly those involving family 1 substrates, have not been determined in postpartum rats or in response to prolactin administration.
In the present study we analyzed the molecular basis of changes occurring in UGT family 1 isoforms (1A1, 1A5, 1A6, and 1A7) and in UGT2B1, a relevant isoform belonging to family 2, in rat liver at late pregnancy and postpartum. For this purpose, enzyme activity toward the classical substrates of the different isoforms and protein and mRNA levels were systematically determined. The role of prolactin on activity and expression of UGTs from family 1 was also analyzed.
Materials and Methods
Chemicals.
[9,11-3H]Androsterone (56.5 Ci/mmol), [2,4,6,7-3H]estrone (76.5 Ci/mmol), and 17α-[6,7-3H (N)]ethynylestradiol (49.1 Ci/mmol) were from PerkinElmer Life Science Products (Boston, MA). Bilirubin, UDPGA (ammonium salt), UDP-N-acetylglucosamine (UDP-N-AG),p-nitrophenol, d-saccharic acid 1,4-lactone, and nonlabeled steroids (androsterone, estrone, and ethynylestradiol) were purchased from Sigma Chemical Co. (St. Louis, MO). Bromocriptine (2-Br-α-ergocriptine methane sulfate) was a gift of Sandoz Research Institute (East Hanover, NJ). Ovine prolactin (NIDDK-ovine prolactin-19; AFP-9221A) was kindly provided by National Institute of Diabetes and Digestive and Kidney Diseases, National Hormone and Pituitary Program, National Institute of Child Health and Human Development, and U.S. Department of Agriculture. All other reagents were of the highest grade commercially available.
Animals.
Female Sprague-Dawley rats (Harlan Industries, Indianapolis, IN) were used throughout. All procedures involving animals were conducted in accordance with National Institutes of Health guidelines for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee of the University of Kentucky. The pregnant and postpartum rats were timed according to the first day that sperm were detected (day 0). The rats had free access to Purina Rat Chow and water and were maintained on a 12-h automatically timed light and dark cycle. Nonpregnant rats (180–210 g) served as controls, and rats at 19 to 20 days of pregnancy (360–400 g) and at 2 to 4 days (240–270 g) and 10 to 12 days (250–280 g) postpartum [pp (2–4 days) and pp (10–12 days) groups] were used as late-pregnant and early- and mid-lactating rats, respectively. Litter size in postpartum animals ranged from 8 to 10 pups.
Two groups of ovariectomized rats weighing 190 to 230 g were implanted with osmotic minipumps (Alzet 2001; Alza, Palo Alto, CA) attached to an intravenous catheter as described by Liu et al. (1992). The minipumps were filled with solvent (0.4 M NaHCO3, 1.6% glycerol, 0.02% sodium azide) (control group) or with solvent plus ovine prolactin to yield an infusion rate of 300 μg of ovine prolactin per day for 7 days (prolactin group). The minipumps were immersed in saline at 37°C to ensure that flow had started. To suppress endogenous prolactin secretion, all the rats were implanted with bromocriptine pellets (7.5 mg, 10 day-release) subcutaneously at the time of implantation of the minipumps.
Enzyme Assays.
All animals were killed by decapitation between 9 and 11 AM, to avoid possible effects of diurnal variations. Liver samples were collected (Luquita et al., 1994) and a portion snap frozen in liquid nitrogen for RNA analysis. Liver homogenate (25% w/v) was prepared in 0.15 M Tris-HCl buffer, pH 7.4 and the corresponding microsomal fractions were obtained by ultracentrifugation at 105,000g for 1 h at 4°C as previously described (Siekevitz, 1962). Protein concentration in microsomal preparations was measured using the Lowry method (Lowry et al., 1951) with bovine serum albumin as standard.
UGT activities were evaluated in microsomes activated with UDP-N-AG (2.0 mM final concentration). Assay conditions for the determination of glucuronidation rate of p-nitrophenol and bilirubin were as previously described (Sánchez Pozzi et al., 1994; Catania et al., 1995). Glucuronidation rate of the neutral steroids androsterone and estrone were determined according to Rao et al. (1977) and conjugating activity of the synthetic derivative of estradiol ethynylestradiol (3-OH and 17β-OH conjugation) was determined under the same incubation conditions (1.0 mM final concentration). In all cases, high performance liquid chromatography was performed to separate the corresponding steroid glucuronides. The mobile phase, consisting of methanol/0.1 M KH2PO4, pH 4.5 (60:40, v/v) was the same as previously used for bile salts separation (Tietz et al., 1984). Detection of radioactive unconjugated and conjugated steroids was accomplished with an IN/US β-RAM flow-through detector (Pine Brook, NJ). The peaks corresponding to glucuronides were confirmed by including a sample preincubated with β-glucuronidase. An ethynylestradiol 17β-glucuronide standard (generous gift of Dr. Brian Burchell, Department of Biochemical Medicine, Ninewells Hospital, Dundee, UK) was used to discriminate between 17β- and 3-derivatives of ethynylestradiol.d-Saccharic acid 1,4-lactone (2 mM) was systematically included in all the incubation media to inhibit enzymatic hydrolysis of glucuronides. Reactions were initiated with the addition of UDPGA to the mixtures.
Western Blot Analysis.
Polyclonal anti-peptide antibodies that specifically recognize the 1A1, 1A5, 1A6, and 1A7 isoforms belonging to UGT family 1 as well as an antibody developed against a peptide common to all isoforms of the same group (1A) (Ikushiro et al., 1995) and a specific antibody against isoform 2B1 (UGT family 2) (Ikushiro et al., 1997) were used in Western blot studies.
Western analyses were performed with microsomal preparations using an amount of protein (15 μg) in the gels that was found to give a densitometric signal in the linear range of the response curve for all antibodies (data not shown). Preparations were loaded onto 10% SDS-polyacrylamide gel (Laemmli, 1970) and subjected to electrophoresis. After electrotransfer onto nitrocellulose membranes (Protran; Scheleicher & Schuell, Keene, NH), the blots were blocked at least for 2 h at 4°C with Tris-buffered saline containing 0.1% Tween 20 and 5% nonfat dry milk and then incubated overnight with the primary antibodies 1A (1:4000), 1A1 (1:4000), 1A5 (1:1000), 1A6 (1:1000), or 1A7 (1:500). The immune complex was detected by incubation with alkaline phosphatase-linked anti-rabbit secondary antibody (1:2000; Sigma Chemical Co.) for 1 h. Immunoreactive bands were detected by the alkaline phosphatase color reaction using 5-bromo-4-chloro-3-indolyl phosphate and nitro blue tetrazolium and quantified by densitometry (Shimadzu CS-9000; Shimadzu, Kyoto, Japan).
Northern Blot Analysis.
Probes consisting of 293-, 308-, and 317-base pair cDNA fragments were amplified from rat liver RNA by reverse transcription-polymerase chain reaction (Access TR-PCR System; Promega, Madison, WI) using the same primers and conditions reported inEmi et al. (1995) to exons B1 (UGT1A1), B5 (UGT1A5), and A1 (UGT1A6), respectively. Exon-specific oligonucleotide primers were synthesized by Bio-Synthesis (Lewisville, TX). In preliminary experiments, the probes thus obtained were able to detect specific induction of different UGT isoforms in response to treatment with clofibrate (UGT1A1 and 1A5) and methylcholanthrene (UGT1A6), as was previously described (Emi et al., 1995). The content of mRNA encoding the family 2 isoform UGT2B1 was analyzed using a full-length rat probe (Mackenzie, 1986), which was generously provided by Dr. Peter Mackenzie (Flinders University of South Australia, Bedford Park, Australia). A single-stranded 26-mer oligoprobe to 28S rRNA (Barbu and Dautry, 1989) was synthesized by Integrated DNA Technologies (Coralville, IA).
Total RNA was isolated from liver samples frozen in liquid nitrogen by a guanidine thiocyanate lysis procedure (Chomczynski and Sacchi, 1987). Total RNA (15 μg) was denatured, electrophoresed through a 1.2% agarose/formaldehyde gel, transferred to a nylon membrane (Hybond-N; Amersham Pharmacia Biotech UK Limited, Little Chalfont, Buckinghamshire, England) overnight by capillary blotting in 6× SCC and baked at 80°C for 2 h. To analyze the content of mRNA encoding UGTs, the membranes were prehybridized in 5× SSC, 0.02% SDS, 0.1% N-laurylsarcosine, and blocking reagent (catalog no. 1096176; Roche Molecular Biochemicals, Mannheim, Germany) at 68°C for at least 30 min. Hybridization was performed at 68°C for 16 h after addition of labeled cDNAs (UGT1A1, 1A5, 1A6, and 2B1). The membranes were then washed in 2× SSC, 0.1% SDS twice at room temperature for 5 min, followed by washing in 0.1× SSC, 0.1% SDS twice at 68°C for 15 min under constant agitation. UGT probes were labeled with digoxigenin-dUTP by a random-primed methodology and detected with an alkaline phosphatase-conjugated antibody followed by chemiluminescent reaction (kit 1585614; Roche Molecular Biochemicals). Differences in RNA loading were corrected after reprobing the stripped blots with 28S RNA. For this purpose, the membranes were prehybridized in 5× SSC, 0.1% SDS, 0.5% dextran sulfate (mol. wt. = 500,000) and blocking reagent (catalog no. RPN 5770; Amersham Pharmacia Biotech, Piscataway, NJ) at 42°C for at least 30 min. Hybridization was performed at 42°C for 2 h after addition of the labeled oligoprobe. The membranes were then washed in 5× SSC, 0.1% SDS twice at room temperature for 5 min, followed by washing in 1× SSC, 0.1% SDS twice at 42°C for 15 min under constant agitation. The 28S oligonucleotide was labeled with fluorescein-dUTP using a 3′-end labeling kit (RPN5776) and detected with an alkaline phosphatase-conjugated antibody followed by chemiluminescent reaction (kit RPN3510; Amersham Pharmacia Biotech). In all cases the blots were exposed to Bio-Max MR-2 film (Sigma Chemical Co.) for various time periods until the autoradiographic signal was optimal. The hybridization bands were then quantified by densitometry (Shimadzu CS-9000; Shimadzu). The densitometry was done in the linear range of the film.
Statistical Analysis.
Data on enzyme activities and densitometric analysis of Western and Northern studies were presented as means ± standard deviation. Statistical analysis was performed using one-way analysis of variance followed by Bonferroni test (pregnancy and postpartum studies) or Student's t test (prolactin studies). Values of p < 0.05 were considered to be statistically significant.
Results
Expression of UGTs in Maternal Rat Liver
UGT Activities.
Enzyme activities toward the different substrates were determined in the presence of UDP-N-AG, which is considered a physiological activator of UGTs (Berg et al., 1995;Bossuyt and Blanckaert, 1995). In addition, under this experimental condition, no substantial disturbance of the membrane environment that could affect catalytic activity of UGTs is expected. Figure1 shows activity values for bilirubin,p-nitrophenol, and ethynylestradiol conjugated in position 3-OH, substrates primarily associated with isoforms of the UGT1 family, and for androsterone, estrone, and ethynylestradiol conjugated in position 17β-OH as substrates that react preferentially with isoforms of the UGT2 family (Ebner et al., 1993; Mackenzie et al., 1997). All substrates associated with UGT1 as well as ethynylestradiol in position 17β-OH exhibited a decreased conjugating activity (55% in average) in pregnant rats, whereas androsterone and estrone conjugation did not change. The data clearly indicate that pregnancy differentially affected conjugation of substrates associated with UGT family 2 isoforms.

UGT activities in maternal rat liver. UGT activities were determined in the presence of the physiological activator UDP-N-AG (2 mM). pp (2–4 days) and pp (10–12 days) represent postpartum rats 2 to 4 and 10 to 12 days after delivery, respectively. Conjugating activity is expressed as nanomoles per minute per milligram of microsomal protein (p-nitrophenol); or as picomoles per minute per milligram of microsomal protein (bilirubin, ethynylestradiol 17β-OH, ethynylestradiol 3-OH, androsterone, and estrone). Data are mean values ± standard deviation, where N = 4 to 6. aSignificantly different from control rats (p < 0.05). bSignificantly different from pregnant rats (p < 0.05).cSignificantly different from pp (2–4 days) rats (p < 0.05).
After delivery, UGT activities toward family 1 substrates gradually returned to control values or even increased forp-nitrophenol (about 40% over controls) by mid-lactation. Androsterone and estrone glucuronidation rates were also increased in postpartum rats, the average increase being 50% with respect to controls by mid-lactation. Thus, changes in the activity of the different UGTs postpartum did not seem to be associated to a particular family, but to specific isozymes belonging to both UGT major families.
Immunoblotting Analysis.
Characterization of expression of the different UGT isoforms by means of specific anti-peptide antibodies showed a decrease in the intensity of immunoreactive bands in pregnant rats for all the isoforms tested (Fig.2). The data clearly indicate that the decreased level of UGT protein in microsomes is the main cause of decreased activities reported above. Within 2 to 4 days after delivery, most of family 1 isoforms as well as UGT2B1 tended to increase their levels, and within 10 to 12 days of delivery, they approached or exceeded values in control rats. In fact, protein content corresponding to 1A1 and 1A6 isoforms was significantly increased with respect to controls in the pp (10–12 days) group. Under the current experimental conditions, the 1A7 isoform was not detected in any of the groups analyzed (data not shown).

UGT levels in maternal rat liver. Equal amounts of microsomal protein (15 μg) were loaded in all lanes. The densitometric analysis is presented as mean values ± standard deviation of relative areas expressed in arbitrary units (N = 3). aSignificantly different from control rats (p < 0.05).bSignificantly different from pregnant rats (p < 0.05). cSignificantly different from pp (2–4 days) rats (p < 0.05).
At least two different isoforms of UGT belonging to family 2 (i.e., 2B1 and 2B3) are involved in conjugation of estradiol and testosterone in position 17β-OH (Mackenzie et al., 1997). Assuming that these same isoforms participate in the glucuronidation of ethynylestradiol at the 17β-OH position, the decrease in content of the 2B1 isoform observed during pregnancy (Fig. 2) may explain down-regulation of UGT activity toward ethynylestradiol (Fig. 1). After delivery, both the expression of UGT2B1 and glucuronidation of ethynylestradiol followed a parallel recovery and gradually returned to control levels along the lactating period. Glucuronidation of androsterone and estrone showed a different pattern of variation along the different perinatal stages compared with ethynylestradiol 17β-OH conjugation (Fig. 1), indicating differential regulatory features for the different family 2 isoforms in maternal liver. Although pregnancy did not affect enzyme activity toward androsterone and estrone, postpartum rats exhibited a significant increase, particularly at mid-lactation. Additional studies are necessary to confirm whether an increase in expression of the isoforms involved in glucuronidation of these neutral steroids (e.g., UGT2B2) is responsible for the increased enzyme activity in microsomal membranes.
Analysis of mRNA Encoding UGTs.
Figure3 shows liver content of mRNA encoding the major UGT1 isoforms and the 2B1 isoform. Interestingly, no change was observed in mRNAs from pregnant rats in contrast to what was observed for protein levels. Within 10 to 12 days after delivery, UGT1A1 and 1A6 mRNA content significantly increased with respect to normal females (40 and 120%, respectively), agreeing well with increased level of protein. Figure 3 also shows that the levels of mRNA encoding UGT1A5 and 2B1 did not change in maternal liver postpartum.

UGT mRNA levels in maternal rat liver. Three major UGT family 1 isoforms and UGT2B1 were analyzed. To correct for differences in total RNA loading and transfer among the lanes, the content of 28S rRNA was also estimated. An additional study was performed with two additional animals per group (data not shown). The densitometric analysis of UGTs mRNA performed on four rats per group in total, is shown on the right. Data represent arbitrary units relative to the corresponding 28S values. aSignificantly different from control rats (p < 0.05).bSignificantly different from pregnant rats (p < 0.05). cSignificantly different from pp (2–4 days) rats (p < 0.05).
Effect of Prolactin on Expression of UGTs
UGT Activities.
Because postpartum rats exhibited an increase in the expression of the isoforms involved in conjugation of bilirubin and p-nitrophenol, it was of interest to establish whether the lactogenic hormone prolactin is involved. Prolactin was administered at a dose that was shown to maximally increase the expression of Na+-taurocholate cotransport polypeptide (ntcp) (Liu et al., 1995) and p-nitrophenol UGT (Luquita et al., 1996) in the rat liver and to increase the P1 subunit of glutathione in the rat intestine (Luquita et al., 1999), thus accounting for increases observed in postpartum rats. Figure4 shows the activity of UGT toward bilirubin and p-nitrophenol in ovariectomized control and ovine prolactin-treated rats. Only p-nitrophenol conjugation was affected by hormone administration, exhibiting a 60% increase with respect to controls.

Effect of prolactin on UGT activities. UGT activities were determined in the presence of the physiological activator UDP-N-AG (2 mM) in ovariectomized controls and in ovine prolactin-treated rats.p-Nitrophenol conjugating activity is expressed as nanomoles per minute per milligram of microsomal protein and bilirubin-conjugating activity as picomoles per minute per milligram of microsomal protein. Data are mean values ± standard deviation, where N = 4 to 5. aSignificantly different from control rats (p < 0.05).
Immunoblotting Analysis.
Figure5 shows the effect of ovine prolactin on the level of the isoforms of UGT involved in bilirubin (UGT1A1 and 1A5) and p-nitrophenol (UGT1A6) conjugation. Only the content of UGT1A6 was increased by the hormone (about 50% over controls), in agreement with the increase in activity of p-nitrophenol conjugation. Neither UGT1A1 nor UGT1A5 was affected by ovine prolactin. As expected, the bands detected with anti-UGT1A, which mainly reflects the level of the isoforms involved in bilirubin conjugation (Ikushiro et al., 1995), were not altered by the hormone. UGT1A7 was not detected in any of the groups analyzed (data not shown).
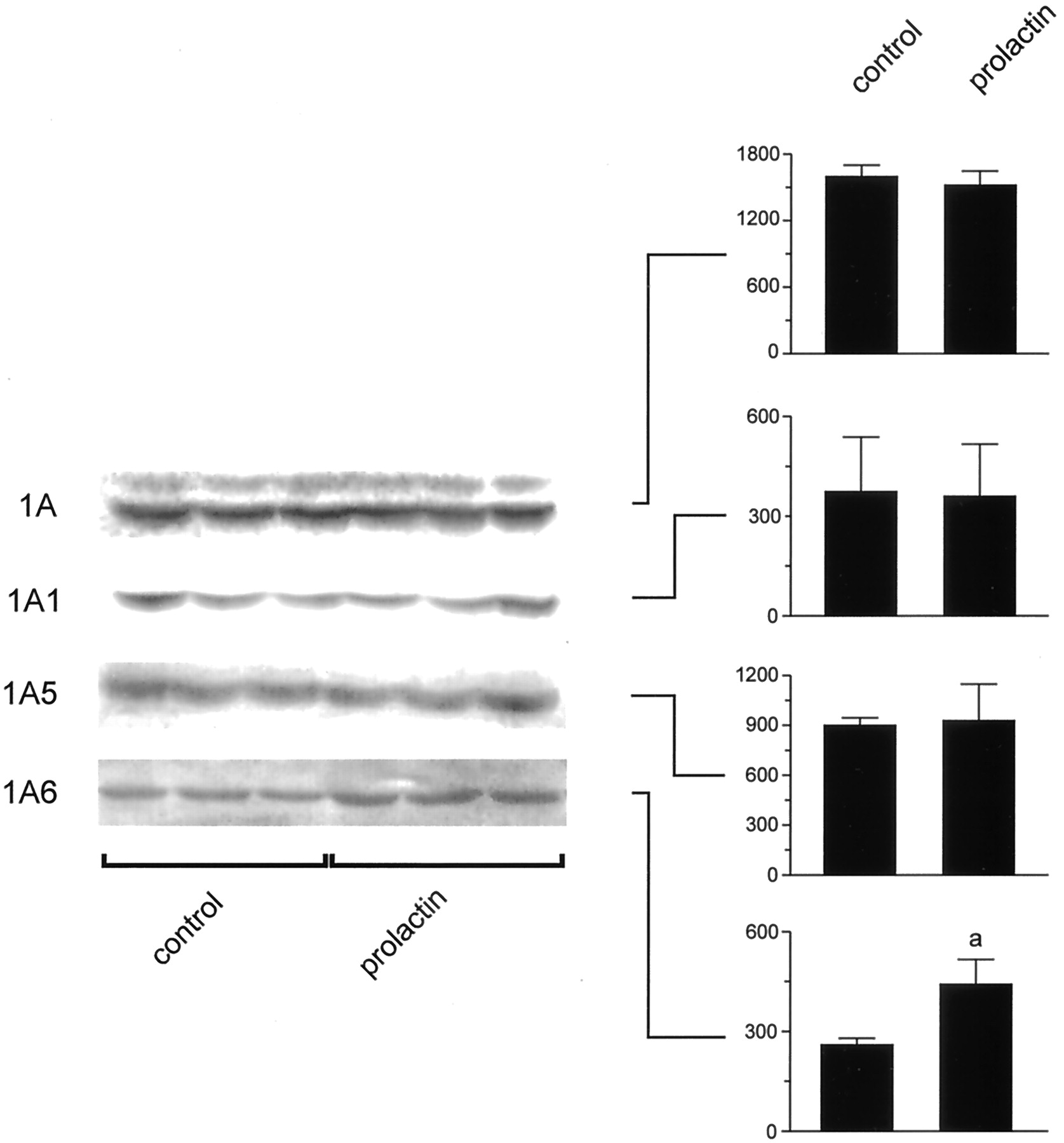
Effect of prolactin on UGT protein levels. Equal amounts of microsomal protein (15 μg) were loaded in all lanes. The densitometric analysis is presented as mean values ± standard deviation of relative areas expressed in arbitrary units (N = 3). aSignificantly different from control rats (p < 0.05).
Analysis of mRNA Encoding UGTs.
Figure6 shows the level of mRNA encoding UGT1A1 and 1A6. Only UGT1A6 mRNA was increased in response to ovine prolactin administration (about 80% over controls). The content of mRNA encoding UGT1A5 was not affected by the hormone (data not shown).

Effect of prolactin on UGT mRNA levels. To correct for differences in total RNA loading and transfer among the lanes, the content of 28S rRNA was estimated. The densitometric analysis of UGTs mRNA performed on three rats per group in total, is shown on the right. Data represent arbitrary units of mRNAs densitometry relative to the corresponding 28S values. aSignificantly different from control rats (p < 0.05).
Discussion
Most studies of UGT activity toward several endogenous and exogenous substrates report down-regulation of UGT-mediated reactions in liver in pregnancy (Halac and Sicignano, 1969; Neale and Parke, 1973; Vore and Soliven, 1979; Muraca et al., 1984; Borlakoglu et al., 1993). Regulation of UGT activity in microsomes is multifactorial, and depends not only on protein expression, but also on functional properties of the corresponding catalytic unit. Using polyclonal anti-peptide antibodies that specifically recognize the 1A1, 1A5, 1A6, and 1A7 isoforms, we here demonstrate for the first time down-regulation of the expression of UGT family 1 isoforms as a main cause of decreased UGT activity in maternal liver during pregnancy. Expression of UGT2B1, an important member of UGT family 2, is also decreased in pregnant rats, in agreement with impaired conjugation of ethynylestradiol in position 17β-OH.
Analysis of liver content of mRNA encoding UGT1 isoforms and UGT2B1 by Northern blotting revealed that pregnancy did not substantially affect their levels (Fig. 3). In consequence, an impairment in gene transcription is unlikely the cause of protein down-regulation. Other factors, such as impairment of protein synthesis, redistribution of protein molecules into an additional membrane compartment (apart from the endoplasmic reticulum), or an increase in protein degradation may be involved. Additional studies are necessary to clarify our understanding of the mechanisms affecting UGTs regulation post-translationally. It is interesting to note that expression of the multidrug-resistant protein Mrp2, the main protein involved in canalicular secretion of conjugated compounds (Keppler et al., 1997), is also down-regulated in female rats during pregnancy (Cao et al., 2001). In these rats, however, mRNA encoding Mrp2 is preserved. Phase II enzymes acting coordinately with Mrp2-mediated secretion of conjugated compounds across the apical domain of epithelial cells may represent an important strategy to protect cells from chemical injury. The molecular basis for the dissociation between protein and mRNA levels occurring during pregnancy for both UGT and Mrp2 is not known. Treatment of male rats with 5 mg of ethynylestradiol per kilogram of body weight for 5 days decreased Mrp2 protein level in liver plasma membranes and was able to mimic dissociation between protein and mRNA (Trauner et al., 1997). Treatment of female rats with twice this dose of ethynylestradiol decreased UGT activity toward estradiol (3-OH and 17β-OH groups) in a similar magnitude as was observed in pregnancy (Connors and Vore, 1988). However, in contrast to what was reported for pregnant rats, administration of estradiol or progesterone increased bilirubin glucuronidation (Muraca et al., 1983). The same hormones added to primary hepatocytes culture did not change the level of mRNA encoding major isoforms of UGT belonging to family 1 and 2 (Li et al., 1999). When administered simultaneously to ovariectomized rats, estradiol and progesterone also did not decrease bilirubin UGT activity (Muraca et al., 1984). Thus, hormonal treatments commonly used to mimic hormonal changes occurring during pregnancy are not able to reproduce alterations in UGT activity, indicating that complex and unknown regulatory factors are involved.
The current data also indicate that during postpartum, UGT activities recovered and even increased with respect to control females. Particularly, p-nitrophenol-conjugating activity showed a significant increase in the pp (10–12 days) group (Fig. 1). This is in agreement with the previous finding in lactating rats at the late stage of lactation (19–21 days postpartum) (Luquita et al., 1994). The current study clearly demonstrates that increased expression of UGT1A6 is the likely explanation for the increase in planar phenol conjugation observed in these rats. In fact, two major isoforms belonging to UGT family 1, i.e., 1A6 and 1A7, are responsible for planar phenol conjugation in the rat (Miners and Mackenzie, 1991; for review, seeBurchell et al., 1994). Using 1A6- and 1A7-specific antibodies, it was previously demonstrated that 1A6, but not 1A7 isoform, is constitutively expressed in rat liver (Ikushiro et al., 1995). In turn, expression of UGT1A7 is more relevant in intestine, probably accounting for conjugating activity toward planar phenols in this tissue. In agreement with the previous observation, we did not find significant expression of 1A7 isoform in liver from normal females and in addition, neither pregnant nor postpartum animals exhibited any significant level of the 1A7 protein. The current study also demonstrates that an enhancement in the level of mRNA encoding UGT1A6 is involved in the increased protein expression postpartum.
Using a polyclonal nonspecific antibody we previously demonstrated an increased content of UGT protein in ovariectomized rats in response to ovine prolactin treatment, suggesting that this hormone may be involved in the regulation of UGT expression postpartum (Luquita et al., 1996). In the second part of the current study, we demonstrated that prolactin was able to specifically increase the expression of UGT1A6 isoform, thus accounting for the increase inp-nitrophenol-conjugating activity. The increase in the level of UGT1A6 mRNA observed in these animals indicates increased gene transcription and/or stabilization of mRNA. Interestingly, ovine prolactin was not able to mimic the increase in expression of UGT1A1 observed in lactating rats. It is possible that other, unknown factors, alone or in combination with prolactin, may be involved in up-regulation of UGT1A1 postpartum. The mechanism of action of prolactin on the liver tissue has been well characterized for protein-mediated transport of taurocholate at the sinusoidal level. In fact, it has been shown that prolactin increases the mRNA encoding the ntcp in the rat liver (Liu et al., 1995) and that this occurs via transcriptional regulation of the ntcp promoter by this hormone (Ganguly et al., 1997). Thus, prolactin acts via the long form of the hormone receptor in the hepatocyte to increase phosphorylation and translocation of a signal transducer and activator of transcription, Stat5, to the nucleus where it binds to γ-interferon-activated sequence elements in the ntcppromoter to increase gene transcription. Further studies are necessary to determine whether a similar mechanism is involved in prolactin-mediated increase in the expression of UGT1A6 in maternal rat liver postpartum.
The analysis of regulation of bilirubin UGT in lactating rats revealed a dissociation between protein and mRNA corresponding to UGT1A1, the main isoform involved in pigment glucuronidation, and enzyme activity. Particularly, it is noteworthy that even when UGT1A1 expression was up-regulated (Figs. 2 and 3) in postpartum animals 10 to 12 days after delivery, microsomal bilirubin glucuronidation was not affected (Fig.1). This dissociation may be tentatively attributed to differences between control and postpartum rats in either protein activity per molecule or protein turnover. The former possibility would imply that postpartum rats might express a number of immunologically reactive but biologically inactive enzyme. Differences in catalytic activity of UGT could result from complex interaction among different monomeric UGTs to form active oligomers or from a particular influence of microsomal membrane environment in postpartum animals, since they critically affect the functional state of UGTs (Peters et al., 1984; Zakim and Dannenberg, 1992; Ikushiro et al., 1997; Meech and Mackenzie, 1997).
In conclusion, we report a decrease in expression of UGT family 1 isoforms as a main cause of down-regulation of UGT-mediated reactions involving bilirubin and planar phenols during pregnancy. Down-regulation of UGT proteins likely occurs at the post-translational level, since mRNA levels were unchanged in control versus pregnant rats. After delivery, protein level from all isoforms gradually returned to control values or even increased (1A1 and 1A6) in association with increased mRNA levels. Prolactin administration was able to mimic the increase in UGT expression observed postpartum only for the 1A6 isoform.
Acknowledgments
We thank Drs. Liyue Huang, Raquel Chan, Daniel Gonzalez, Silvina Fellitti, Lucrecia Alvarez, Cristian Magni, Silvina Pessino, and Javier Palatnik for technical assistance and valuable suggestions.
Footnotes
-
This work was supported by Research Grants from Consejo Nacional de Investigaciones Cientı́ficas y Técnicas, Universidad Nacional de Rosario, and Subsecretarı́a de Investigación y Tecnologı́a, Ministerio de Salud de la Nación, Argentina; and by U.S. Public Health Service Grants GM55343 and NS31220.
- Abbreviations:
- UGT
- UDP-glucuronosyltransferase
- UDPGA
- UDP-glucuronic acid
- UDP-N-AG
- UDP-N-acetylglucosamine
- pp
- postpartum
- SSC
- standard saline citrate
- ntcp
- Na+-taurocholate cotransport polypeptide
- Mrp
- multidrug-resistant protein
- Received January 17, 2001.
- Accepted March 13, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}