


Abstract
The masculine profile of growth hormone (GH) secretion characterized by episodic bursts (∼200–300 ng/ml plasma) every 3.5 to 4 h, separated by interpulse periods devoid of detectable hormone, was restored at various peak heights to hypophysectomized, thyroxine-supplemented male rats to determine the minimum signaling amplitudes of the hormone pulse required to maintain male-like expression levels of gender-dependent hepatic cytochrome P-450s (CYP P-450s). Restoration of the pulse to as little as 2.5% of normal elevated CYP2C11 (the predominant isoform in male liver) protein and dependent catalytic activities to ∼50% of normal, whereas transcript concentrations increased to 150% of physiologic. Renaturalizing the masculine plasma GH profile to 5% of normal was sufficient to increase CYP2C11 protein and catalytic activity to intact levels while further elevating mRNA to ∼200% of normal (subsequently declining to intact concentrations with physiologic pulses). In dramatic contrast, CYP2C7 (mRNA and protein) declined to barely detectable levels following hypophysectomy and remained completely unresponsive to GH until replaced with the physiologic masculine profile. The repressive effects of the episodic GH profile on CYP2A2 and CYP3A2 expression similarly required replacement of near physiologic pulse amplitudes. Exhibiting an intermediate response to the masculine profile, restoration of 25% of the normal pulse amplitude was sufficient to significantly elevate CYP2A1 and CYP2C6 expression levels in hypophysectomized rats. These findings illustrate the importance of the pulse amplitudes (in addition to the interpulse periods) in the circulating masculine GH profile as differential signals regulating the expression and/or repression of each sex-dependent hepatic P-450 isoform in the rat.
Rat liver contains at least a dozen sex-dependent isoforms of cytochrome P-4503 (CYP P-450) that are regulated by the gender-dependent profiles of circulating growth hormone (GH) (Legraverend et al., 1992a; Waxman, 1992). Male rats secrete GH in episodic bursts (∼200–300 ng/ml plasma) every 3.5 to 4 h. Between the peaks, GH levels are undetectable. In females, the hormone pulses are more frequent and irregular and are of lower magnitude than those in males, whereas the interpulse concentrations of GH are always measurable (Legraverend et al., 1992a; Shapiro et al., 1995).
In the rat, P-450 responses to GH regulation are almost as variable as the number of GH-dependent isoforms. That is, expression of the major female-specific CYP2C12 (as well as the non-P-450 5α-reductase) is dependent on the feminine profile of continuous GH secretion. Exposure to the masculine profile of episodic hormone release, as well as the absence of the hormone from the circulation (e.g., hypophysectomy), completely prevents expression of CYP2C12 (Ram and Waxman, 1990;Legraverend et al., 1992b). In a somewhat similar vein, female-predominant CYP2C7 expression is also dependent on the feminine GH profile and is completely suppressed in the hypophysectomized rat. However, exposure to the masculine profile allows expression of CYP2C7 at 25 to 40% of normal female levels (Ram and Waxman, 1990; Westin et al., 1990; Legraverend et al., 1992a). Expression of the major male-specific CYP2C11 requires the episodic “on/off” masculine profile of GH secretion. Although the feminine pattern of continuous hormone secretion blocks CYP2C11 expression, total GH depletion from the circulation allows CYP2C11 expression at 20 to 30% of intact male levels (Morgan et al., 1985; Janeczko et al., 1990; Legraverend et al., 1992b). After hypophysectomy, female-predominant CYP2A1 (male/female; ∼1:3) concentrations decline but remain above male levels and are restored to intact female-like levels with continuously administered GH (Waxman et al., 1990a; Yamazoe et al., 1990). Although the expression levels of CYP2C7, CYP2C11, CYP2C12, and CYP2A1 are greatest when exposed to their gender-dependent GH profiles, other isoforms are optimally expressed in the absence of GH. Male-specific CYP2A2 and CYP3A2 are maximally expressed in the hypophysectomized rat, disappear when GH is secreted constantly, but are only partially suppressed under the influence of episodic GH (Waxman et al., 1988, 1995a). Male-specific CYP2C13 is optimally expressed when exposed to the masculine hormone profile or under conditions of no GH, whereas the feminine GH profile completely suppresses CYP2C13 (Legraverend et al., 1992a,b). Although there are additional examples, it becomes clear that the expression or suppression of each isoform of P-450 is likely to be regulated by a different “signal” or perhaps, a differential sensitivity to the “signal”, in the sexually dimorphic GH profiles. These signals may be recognized by the hepatocyte in the frequencies and/or durations of the pulse and interpulse periods. Alternatively, perhaps the hepatocyte can monitor the mean plasma concentration of the hormone. In the latter case, with the selective-GH deficient monosodium glutamate (MSG)-treated rat, we reported that a 70 to 85% reduction in the feminine GH profile has little, if any, effect on the levels of gender-dependent P-450 isoforms normally expressed in the female liver (Waxman et al., 1990b; Pampori and Shapiro, 1994a). More recently, with the euthyroid, hypophysectomized rat (Pampori and Shapiro, 1996), we observed that a renaturalized feminine GH secretory pattern of only 2 to 3% of its normal level (the lowest values we examined) remained completely suppressive of male-specific CYP2A2, CYP2C11, CYP2C13, and CYP3A2 expression. In contrast, the female-dependent isoforms exhibited a variable responsiveness to the restored hormonal profile. Female-predominant CYP2A1 and CYP2C6 were restored to prehypophysectomy levels when the feminine GH profile was renaturalized to just 3% of normal. Although this level of GH restored female-specific CYP2C12 and 5α-reductase to ∼40 and ∼70% of normal, respectively, restoration to intact expression levels required a circulating feminine profile of 12 to 25% of normal. Female-predominant CYP2C7 appeared to be the least GH-sensitive isoform. Although exhibiting a commensurate increase in mRNA and protein concentrations with increasing levels of GH, the feminine GH profile had to be restored to near 100% (i.e., physiologic) to induce normal female-like expression levels of CYP2C7.
Although these studies have identified the concentration in the continuous feminine GH profile as the intrinsic signaling element regulating the characteristic expression of hepatic P-450 isoforms observed in the female rat, the present study has examined the importance of the sizable pulses in the episodic masculine GH profile in regulating the expression of sex-dependent hepatic P-450s found in the male rat.
Materials and Methods
Animals.
Animals were housed in the University of Pennsylvania Laboratory Animal Resources facility under the supervision of certified laboratory animal medicine veterinarians and were treated according to a research protocol approved by the University's Institutional Animal Care and Use Committee. Male rats [Crl:CD(SD)BR] were hypophysectomized by the vendor (Charles River Laboratories, Wilmington, MA) between 7 and 8 weeks of age and were observed in our facilities for 5 weeks. The effectiveness of the surgery was verified by the lack of weight gain over this period and the absence of pituitaries or fragments at necropsy at the end of the study (i.e., ∼100 days old) in ∼85 to 90% of the initial cohort.
Hormone replacement experiments with rat GH (rGH) (1.8 I.U./mg) were begun when hypophysectomized male rats were ∼13 weeks of age. Periodic injections via a chronic indwelling right atrial catheter implant and controlled by an external syringe pump (MacLeod and Shapiro, 1988; Pampori et al., 1991a) were administered as 3-min pulses at 4-h intervals at doses from 0 to 40 μg of rGH per kilogram body weight per injection, as specified in the tables and figure legends, for 7 consecutive days. The amount of rGH in each pulse was chosen to replicate 0 to 100% of the normal masculine pulse parameters. To verify the effectiveness of the pumping apparatus, concurrent blood samples (25 μl) were obtained at 15-min intervals from four rats in each rGH-treatment group (as well as from intact rats). Eight-hour plasma rGH profiles were determined with a radioimmunoassay with a sensitivity of 2 to 3 ng/ml. Procedural details and statistical validation of the assay have been reported previously (Shapiro et al., 1989).
The atrial catheterizations were performed 4 to 5 days before initiation of the rGH treatments. At the time of surgery, all the hypophysectomized rats were s.c. implanted with osmotic pumps (Alza Corp., Palo Alta, CA) set to continuously deliver, for 14 days, thyroxine at a dosage (0.8 μg/h/kg b.wt.) that produced the euthyroidism (Emerson et al., 1989) required to maintain normal concentrations of NADPH-CYP P-450 reductase, a microsomal enzyme required for the expression of P-450 catalytic activity (Ram and Waxman, 1992).
Animals were decapitated within 2 h of the last administered pulse; the livers were quickly removed and perfused with ice-cold saline. Each liver was quickly minced; a portion reserved for mRNA determination was plunged into liquid nitrogen and subsequently stored at −70°. The remaining minced liver was used for microsomal preparation.
RNA Analysis.
Total hepatic RNA was isolated with a single-step guanidinium thiocyanate method (Chomczynski and Sacchi, 1987). Ten micrograms of RNA was electrophoresed under formaldehyde-denaturing conditions on 1% agarose and transferred to GeneScreen nylon membranes (DuPont-New England Nuclear, Boston, MA). The Northern blots were probed and reprobed with32P-labeled oligonucleotide probes, with hybridization and high stringency washing conditions as described previously (Waxman, 1991). The nucleotide sequence of oligonucleotide probes for CYP2A1, CYP2A2, CYP2C6, CYP2C7, CYP2C11, CYP2C12, CYP2C13 (Waxman, 1991), and CYP3A2 (Ram and Waxman, 1991) have been reported. The consistency of RNA loadings between samples was confirmed by ethidium bromide staining of 18S and 28S ribosomal RNAs and was verified with an 18S oligonucleotide probe (Ramsden et al., 1993). The hybridized mRNA signals were quantified by scanning the autoradiographs and normalized to the 18S rRNA signals in each lane.
Western Blots.
Hepatic microsomes were prepared from individual rat livers (Shapiro and Szczotka, 1984) and then assayed for individual P-450s by Western blotting and/or by measurement of their selective catalytic activities (Waxman, 1991; Agrawal et al., 1995). Briefly, 10 μg of microsomal protein was electrophoresed on 0.75-mm-thick sodium dodecyl sulfate-polyacrylamide (7.5%) gels and electroblotted onto nitrocellulose filters. The blots were probed with monoclonal anti-rat CYP2C11 (Oxford Biomedical Research, Oxford, MI) and anti-rat CYP2C12/13 (kindly provided by Dr. Marika Rönnholm, Huddinge University Hospital, Huddinge, Sweden) mouse IgG, polyclonal anti-rat CYP2C7 (kindly provided by Dr. Stelvio M. Bandiera, The University of British Columbia, Canada), and anti-rat CYP3A1/2 (Human Biologics, Phoenix, AZ) rabbit IgG, and detected with an enhanced chemiluminescence kit (Amersham, Arlington Heights, IL) (Pampori et al., 1995).
The specificity of the antibodies has been discussed elsewhere (Pampori and Shapiro, 1996). Briefly, antibodies against CYP2C7 and CYP2C11 have been found to be highly specific with no detectable cross-reactivities. Antibodies to CYP2C12 strongly react with CYP2C13 protein. However, not only is CYP2C13 a male-specific isoform (in contrast to the female-specificity of CYP2C12), but its location on the blot is easily distinguished from CYP2C12. Although CYP3A1 and CYP3A2 proteins are recognized by the anti-rat CYP3A1/2, the former is basically an inducible, GH-independent isoform only marginally expressed constitutively, whereas the latter is a major male-specific isoform.
Catalytic Activity.
Testosterone metabolites, including 2α- and 16α-, 7α-, 15α- and 6β-hydroxylases, reflective of the activity levels of CYP2C11, CYP2A1, CYP2A2, and CYP3A2 proteins, respectively (Schenkman, 1992), and female-specific testosterone 5α-reductase (coincidental with CYP2C12) were assayed according to our methods as described previously (Pampori et al., 1991b; Agrawal et al., 1995). Male-predominant, multi-P-450-dependent microsomal hexobarbital hydroxylase was measured as reported in Shapiro and Szczotka (1984).
Statistics.
The characteristics of plasma GH pulses were analyzed with the aid of the Cluster analysis computer program (Veldhuis and Johnson, 1986) as we reported previously (MacLeod and Shapiro, 1988; Agrawal and Shapiro, 1996). All data, including those obtained from the Cluster analysis program, were subjected to ANOVA, and differences were determined with t statistics and the Bonferroni procedure for multiple comparison.
Results
Hormone Delivery Systems.
Whereas almost all reports examining the effects of GH replacement on P-450 expression do not verify the resulting plasma profiles of the hormone, we considered it essential to do so in the present study. In addition to actually measuring the restored circulating GH profiles in the rGH-replaced hypophysectomized rats (Fig. 1 and Table1), we also evaluated the accuracy and precision of the GH delivery system. Initially, each morning when the pump syringes were replaced with new sterile syringes containing freshly prepared rGH, the remaining volume in the spent syringes were measured as a rough determination of the actual amount of hormone administered during the past 24 h. With the exception of two syringes (of some 500 used) for which the pumps malfunctioned during the evening and the effected rats removed from the study, all other spent syringes were found to contain the expected residual volumes. Subsequently, to verify that the pumps contained and delivered the correct dosage of rGH, hormone levels in the residual volumes of the spent syringes were measured by radioimmunoassay and found to contain 99 ± 8% (mean ± S.D.) of the expected values.

Plasma levels of circulating rGH obtained from individual undisturbed catheterized intact and euthyroid hypophysectomized (HYPOX) rGH-replaced male rats at 15-min intervals for eight consecutive hours. Masculine-like plasma growth hormone profiles were restored to HYPOX rats by administering the hormone in six daily pulses, once every 4 h, through an external pump attached to an indwelling atrial catheter (MacLeod and Shapiro, 1988;Pampori et al., 1991a). The various concentrations of rGH (μg/kg b.wt.) infused per pulse are indicated (center). Depicted next to each infusion rate are the resulting circulating profiles normalized by subtracting plasma values obtained from hypophysectomized rats. ●, values determined by radioimmunoassay. Similar findings were obtained from three to four additional animals in each treatment group. (Note the different scales on the y-axes.)
Analysis of restored growth hormone pulses in hypophysectomized male rats
To evaluate the effectiveness of the osmotic pump delivery of thyroxine, we measured the volume of residual hormone in each pump at the time of necropsy and compared our empirical findings with the expected values. The calculated mean ± S.D. residual volume of thyroxine in all the osmotic pumps used in the study was 99 ± 6% of the expected value.
Plasma GH Replacement Profiles.
In agreement with previous observations (Shapiro et al., 1995), intact (control) male (Fig. 1) and female (data not shown) rats exhibited sexually dimorphic patterns of GH secretion described in the introduction. Episodic infusion from 1 to 40 μg rGH/kg b.wt./pulse by our pump apparatus produced masculine-like circulating profiles of GH characterized by a regular pulse every 4 h interrupted by undetectable concentrations of the hormone for ∼3 h (Fig. 1). Statistical analysis of the pulses produced by episodic infusion of the 40-μg rGH dose indicated that the amplitudes (height), durations (width), and areas of the renaturalized peaks were indistinguishable from the naturally occurring pulses (Table 1). In fact, there was a proportional relationship between the doses of rGH infused and the amplitudes and areas of the resulting plasma GH peaks. (In contrast, although each reduction in the dose of administered rGH resulted in a commensurate decline in the widths of the plasma GH peaks, the relationship was nonproportional.) When the administered pulse of rGH was reduced from 40 to 20 μg, the mean plasma peak amplitude and area (120 ± 19 ng/ml and 1.89 ± 0.41 μg/min/ml, respectively) declined to ∼50% of normal. A further 50% reduction in GH replacement to 10 μg rGH/kg b.wt./pulse produced circulating hormone peaks that were ∼25% of normal. Reduction in the concentration of the infused rGH pulse to 4 μg resulted in mean plasma peak amplitudes and areas (23 ± 4 ng/ml and 0.34 ± 0.06 μg/min/ml, respectively) declining to ∼10% of normal. A decrease in the administered rGH pulse to 2 μg produced circulating peak characteristics that were ∼5% of normal, and an additional 50% reduction in the infused pulse to 1 μg resulted in plasma GH peaks that were ∼2.5% of normal. GH replacement at the rates of 0.4, 0.2, 0.1, 0.04, and 0.02 μg rGH/kg b.wt./pulse produced circulating GH peaks that were below the statistical sensitivity of the radioimmunoassay. However, because the higher concentrations of GH infusion resulted in proportional and predictable plasma hormone levels, we thought it reasonable to extrapolate (by linear regression) plasma rGH peak levels for the hypophysectomized males administered pulses of 0.4, 0.2, 0.1, 0.04, and 0.02 μg rGH/kg b.wt. to ∼1, 0.5, 0.25, 0.1, and 0.05% of normal peak levels, respectively (Table 1).
Hepatic CYP2C11.
The male specificity of CYP2C11 was illustrated by its expression in intact male liver and its absence in female liver (Fig. 2). With the disappearance of the masculine pattern of episodic GH secretion in the hypophysectomized male rat, expression of the isoform (i.e., mRNA, protein, and highly specific CYP2C11-dependent testosterone 2α-hydroxylase and less-specific testosterone 16α-hydroxylase) was reduced to a baseline of ∼25% of the normal male levels. Restoration of the masculine plasma GH pulse to 0.05, 0.1, 0.25, 0.5, and 1.0% of normal had no inductive effects on CYP2C11 expression in the hypophysectomized rats. Restoration of the pulse to as little as 2.5% of normal significantly (P < .01) elevated CYP2C11 protein to ∼45% of normal, catalytic activities to ∼60% of normal and mRNA to ∼150% of normal. Renaturalization of the masculine plasma GH profile to 5% of normal was sufficient to stimulate CYP2C11 protein and dependent catalytic activities to intact (control) levels, whereas CYP2C11 mRNA was further elevated to near twice normal levels. CYP2C11 mRNA peaked at >200% of normal when GH pulses were restored to 10% of intact levels. Thereafter, mRNA levels declined (although remaining significantly above control levels) as rGH pulses increased to 25 and 50% of normal, and finally decreasing to normal expression levels when the GH peak was restored to physiologic (100% of the control pulse). In this regard, there was a small but significant (P < .01) overexpression of CYP2C11-dependent testosterone 16α-hydroxylase activity and CYP2C11 protein when the circulating GH was restored to 10 and 25% of normal, respectively.

Relative hepatic CYP2C11 mRNA, protein and catalytic activity levels in intact female (♀) and intact male (♂) rats and euthyroid hypophysectomized (HYPOX) rGH-replaced male rats. The levels of rGH replacement are presented as a percentage of the normal masculine episodic plasma pulse illustrated in Fig. 1 and determined inResults (Table 1). Relative CYP2C11 mRNA and protein levels determined by laser densitometry of actual Northern autoradiographs and Western enhanced chemiluminescence autoradiographs and microsomal CYP2C11-dependent testosterone 2α-hydroxylase (T 2αOH) and testosterone 16α-hydroxylase (T 16αOH) levels of at least six different livers for each treatment group (means ± S.D.). ND, not detected. a, P < .01 compared with the intact male rats; b, P < .01 compared with the vehicle-treated (0%) hypophysectomized male rats.
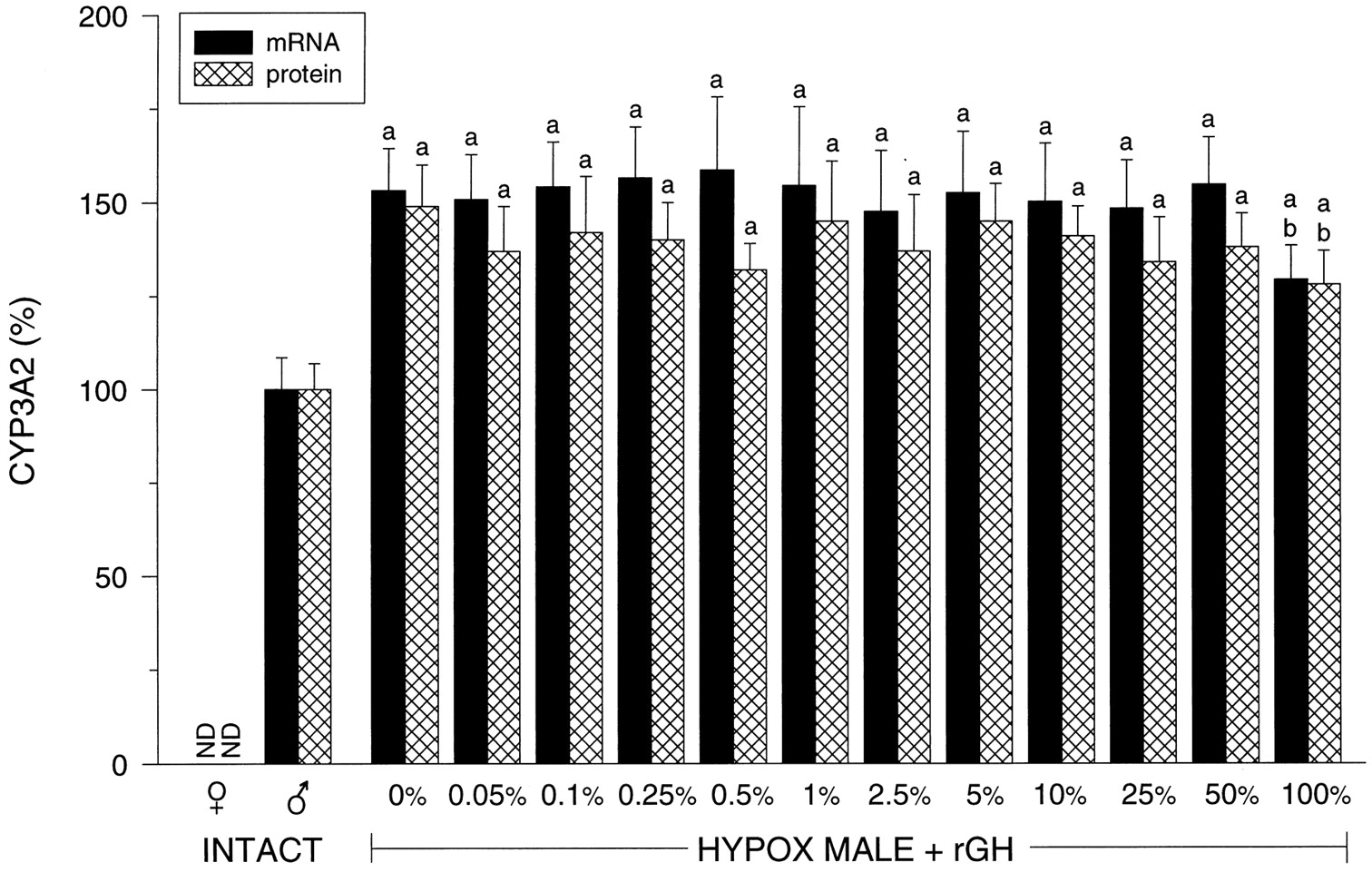
Hepatic CYP3A2.
CYP3A2 is a male-specific isoform whose mRNA and protein expression were undetectable in intact female liver (Fig.3) The complete elimination of circulating GH by hypophysectomy resulted in an overexpression of the isoform (i.e., 40 to 50% above normal) that remained consistently unresponsive to the suppressive effects of the masculine GH profile until the pulse was restored to the 100% level. CYP3A2-dependent testosterone 6β-hydroxylase activity was in agreement with mRNA and protein levels (data not shown).
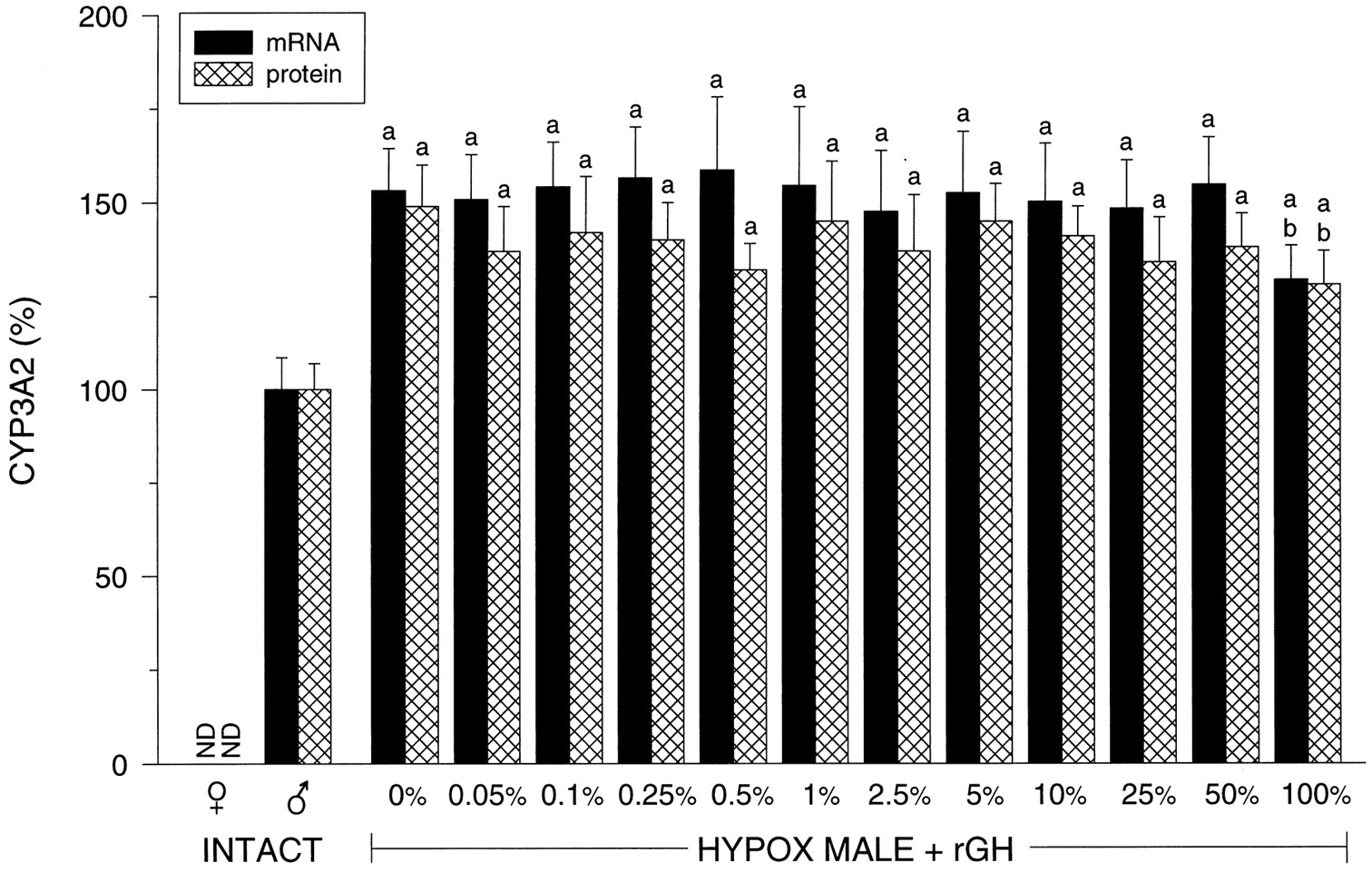
Relative hepatic CYP3A2 mRNA and protein levels in intact female (♀ ) and intact male (♂) rats and euthyroid hypophysectomized (HYPOX) rGH-replaced male rats. The levels of rGH replacement are presented as a percentage of the normal masculine episodic plasma pulse illustrated in Fig. 1 and determined inResults (Table 1). Relative CYP3A2 mRNA and protein levels determined by laser densitometry of actual Northern autoradiographs and Western enhanced chemiluminescence autoradiographs of at least six different livers for each treatment group (means ± S.D.). ND, not detected; a, P < .01 compared with the intact male rats; b, P < .01 compared with the vehicle-treated (0%) hypophysectomized male rats.
Hepatic CYP2A2.
Like CYP3A2, CYP2A2 is a male-specific isoform completely suppressed by the feminine continuous secretory GH profile, but overexpressed, in comparison to intact males, in hypophysectomized rats (Fig. 4) GH depletion produced an only ∼20% borderline significant increase in the isoform that persisted with all replacement doses of rGH except at the 100% level, which reduced CYP2A2 mRNA to intact male concentrations. Although CYP2A2-dependent testosterone 15α-hydroxylase activities tended to agree with CYP2A2 mRNA levels, the differences between treatments were too small to be statistically validated (data not shown).

Representative Northern blots of hepatic CYP2A2, 2C13, 2C12, and 2C6 mRNAs in intact female (♀) and intact male (♂) rats and euthyroid hypophysectomized (HYPOX) rGH-replaced male rats. The levels of rGH replacement are presented as a percentage of the normal masculine episodic plasma pulse illustrated in Fig. 1 and determined in Results (Table 1). Although not presented, relative CYP mRNA levels determined by laser densitometry of actual Northern autoradiographs of at least six different livers for each treatment group are referred to in Results. The same Northern blots reanalyzed with a 32P-labeled oligonucleotide probe specific for 18S rRNA was used as a control to indicate equal loading of the RNA in all lanes.
Hepatic CYP2C13.
Because male-specific CYP2C13 was maximally expressed when either exposed to the masculine GH profile (intact males) or under conditions of no GH (hypophysectomized males) it is not surprising that none of the replacement doses of rGH had any effect on CYP2C13 mRNA levels (Fig. 4). In contrast, CYP2C13 mRNA expression was completely suppressed in control females. Although not presented, CYP2C13 protein levels were in agreement with mRNA values.
Hepatic CYP2C12.
Being obligatorily dependent on the feminine GH profile for its expression, CYP2C12 is presently the only female-specific hepatic isoform in the rat. Accordingly, CYP2C12 mRNA (Fig. 4), protein, and testosterone 5α-reductase type I (data not shown) were completely suppressed in livers of intact males and all hypophysectomized males, irrespective of rGH treatment. Only the livers of intact females expressed CYP2C12.
Hepatic CYP2C6.
CYP2C6 was found to be female-predominant with males expressing ∼60% of the hepatic concentration of the transcript found in females (Fig. 4). When male rats were hypophysectomized, their hepatic CYP2C6 mRNA levels more than doubled, now being 40 to 50% greater than concentrations found in intact females (Fig. 4). Episodic GH replacement had no effect on the overexpressed transcript in the hypophysectomized males until the dose restored the rGH pulse to 25% of normal. However, restoration of GH pulses at 25 and 50% of normal suppressed CYP2C6 mRNA only to female-like levels. It required normal (i.e., 100%) rGH pulses to reduce CYP2C6 mRNA to the lower male-like concentrations.
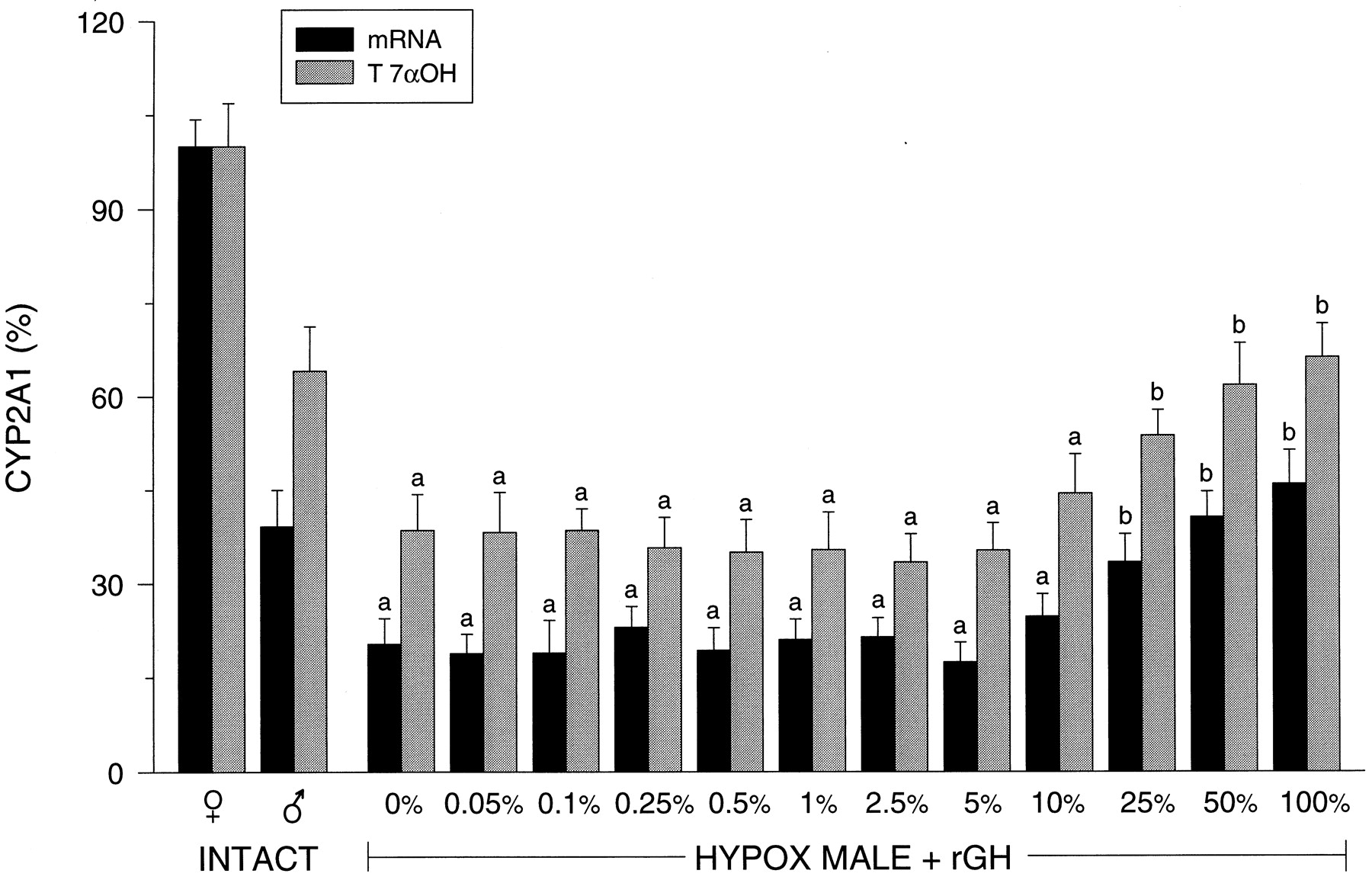
Hepatic CYP2A1.
Similar to CYP2C6, CYP2A1 was found to be female-predominant, with males expressing lower hepatic concentrations of the transcript (∼60% less) and CYP2A1-dependent testosterone 7α-hydroxylase (∼40% less) than females (Fig.5). Hypophysectomy of male rats resulted in a 50% decline in CYP2A1 expression with the isoform remaining unresponsive to rGH infusion until plasma pulses of the hormone were elevated to 25% of normal. However, it was not until GH levels were restored to 50% of normal that CYP2A1 was expressed at intact male-like levels. [Although consistent with our earlier findings (Pampori and Shapiro, 1996) of a hypophysectomy-induced, gender-independent decline in hepatic CYP2A1 mRNA, protein concentrations have been reported, for inexplicable reasons, to increase or change little following GH depletion (Waxman et al., 1989;Yamazoe et al., 1990).]

Relative hepatic CYP2A1 mRNA and catalytic activity levels in intact female (♀) and intact male (♂) rats and euthyroid hypophysectomized (HYPOX) rGH-replaced male rats. The levels of rGH replacement are presented as a percentage of the normal masculine episodic plasma pulse illustrated in Fig. 1 and determined inResults (Table 1). Relative CYP2A1 mRNA levels determined by laser densitometry of actual Northern autoradiographs and microsomal CYP2A1-dependent testosterone 7α-hydroxylase (T 7αOH) levels of at least six different livers for each treatment group (means ± S.D.). a, P < .01 compared with the intact male rats; b, P < .01 compared with the vehicle-treated (0%) hypophysectomized male rats.
Hepatic CYP2C7.
CYP2C7 is another female-predominant isoform whose mRNA and protein levels in females were expressed at three to four times that observed in males (Fig.6). In the absence of GH, CYP2C7 expression in males was reduced to barely detectable. Restoration of the masculine episodic plasma GH profile at all pulse levels, except at 100%, were ineffective in inducing CYP2C7 mRNA or protein expression. GH pulses restored to the normal masculine amplitude (i.e., 100%) resulted in a commensurate induction of the isoform to the physiologic intact masculine level.

Relative hepatic CYP2C7 mRNA and protein levels in intact female (♀) and intact male (♂) rats and euthyroid hypophysectomized (HYPOX) rGH-replaced male rats. The levels of rGH replacement are presented as a percentage of the normal masculine episodic plasma pulse illustrated in Fig. 1 and determined inResults (Table 1). Relative CYP2C7 mRNA and protein levels determined by laser densitometry of actual Northern autoradiographs and Western enhanced chemiluminescence autoradiographs of at least six different livers for each treatment group (means ± S.D.). a, P < .01 compared with the intact male rats; b, P < .01 compared with the vehicle-treated (0%) hypophysectomized male rats.
Hepatic Hexobarbital Hydroxylase.
Hepatic microsomes from intact male rats contained ∼5 times the activity of multi-P-450-dependent hexobarbital hydroxylase than that observed in intact females (Fig. 7). Hypophysectomy reduced the activity of the monooxygenase in males to female-like levels. Restoration of the masculine plasma GH profile had no inductive effect until the hormone pulse was restored to 2.5% of the normal level, producing a 100% increase in hexobarbital hydroxylase over hypophysectomy concentrations. Renaturalizing GH pulses to 5.0% or greater completely restored masculine levels of the monooxygenase.

Relative hepatic hexobarbital hydroxylase activity levels in intact female (♀) and intact male (♂) rats and euthyroid hypophysectomized (HYPOX) rGH-replaced male rats. Various concentrations of rGH were infused through an atrial catheter to replicate 0 to 100% normal masculine pulse parameters as illustrated in Fig. 1 and determined in Results (Table 1). Values are presented as means ± S.D. with an n ≥ 6. a, P < .01 compared with the intact male rats. b, P < .01 compared with the vehicle-treated (0%) hypophysectomized male rats.
Miscellaneous Microsomal Measurements.
Microsomal protein (milligrams per gram liver) in intact males (17.5 ± 1.2; mean ± S.D.) was dramatically reduced (11.0 ± 1.1) following hypophysectomy. Whereas replacement of the episodic GH profile at 25 to 50% of normal increased microsomal protein concentrations to female-like levels (14.1 ± 1.0), only the complete restoration of the pulse to normal (i.e., 100%) could elevate hepatic protein concentrations in the hypophysectomized males (16.7 ± 1.0) to intact levels.
Hepatic microsomal enzymes responsible for the metabolism of testosterone to 7β-hydroxytestosterone and 16-ketotestosterone were both found to be female predominant (2:3; M:F). Hypophysectomy resulted in a 50 to 60% decrease in the enzyme activities of males. Whereas the lowest replacement dose of rGH to induce significant increases in the enzyme levels was between 25 and 50% of normal, it was only the 100% replacement dose that restored the enzyme activities to intact male-like concentrations (data not shown).
Body and Organ Weights.
Hypophysectomy was associated with a small (i.e., 1 to 2 g) weekly decline in body weight (Table2). Restoration of the masculine GH profile to 5% of normal was the lowest dose of the hormone to stimulate a significant increase in body weight gain. Replacement of the GH pulse at 25 to 50% of normal was required to restore body weight gain to intact masculine levels.
Body weight gains and organ weights of hypophysectomized male rats receiving renaturalized masculine profiles of circulating GH
Relative liver weights were depressed by hypophysectomy and were not restored to that found in intact males until GH pulses were renaturalized to 25 to 50% of normal. Like liver weights, relative kidney weights were reduced ∼20% following hypophysectomy in the male. The loss in renal weight could not be corrected until masculine GH profiles were restored to 50% of normal.
Although it is well known that testicular and seminal vesicle weights are primarily dependent on gonadotropin and androgen stimulation, respectively, our findings indicate some stimulatory effects by GH (Table 2). As anticipated, hypophysectomy resulted in a profound loss in testicular and seminal vesicle weights. Although not capable of restoring the organ weights to presurgical levels, restoration of the GH pulse to ∼25% of normal significantly increased testes and seminal vesicle relative weights that were maximized, albeit incompletely, by 100% replacement of the pulse.
Although containing barely trace amounts of adrenocorticotropin contamination (A. F. Parlow, Scientific Director, National Hormone and Pituitary Program, personal communication), the ∼45% decrease in adrenal weights following hypophysectomy were completely restored when circulating GH pulses were renaturalized to 50% of normal. Lower doses had no effect.
Discussion
Whereas the hypophysectomized rat has proven to be a useful model4 in our studies to identify the fundamental signaling elements in the plasma GH profile (which could be considered the initial signal, albeit extrahepatic, in the signal transduction pathway regulating P-450 expression), such signaling studies require that certain procedures, which are often overlooked, be followed.
Having found that plasma GH concentrations either too low to be measurable (Pampori and Shapiro, 1996) or as presented herein, on the borderline of assay sensitivity, are capable of maintaining the sexually dimorphic hepatic P-450 profiles without influencing body weight gain, it is essential that the completeness of the hypophysectomy be confirmed (Pampori and Shapiro, 1996). Clearly, residual pituitary fragments insufficiently large to affect growth could still alter P-450 expression.
Because each isoform of P-450 is regulated by different signals in the circulating GH profile, it is important to know the resulting plasma profiles produced by administered GH. Accordingly, the often-reported observation that two daily s.c. injections of GH can masculinize hepatic P-450s (Legraverend et al., 1992a; Waxman, 1992) may have led to the erroneous conclusion that it is the elevated, short-lived peaks in the plasma hormone profile of the male rat that signals masculinization of the isoforms. To the contrary, monitored s.c. injections of GH were found to produce low-amplitude, very long-lived plasma plateaus that had no resemblance to the endogenous pulse (Pampori et al., 1991a). Unfortunately, almost all studies examining the effects of GH replacement on P-450 expression neglect monitoring the renaturalized plasma GH profiles, making it difficult to know the kind of plasma profile to which the hepatocyte was exposed.
When trying to identify what might be very subtle signals in the sexually dimorphic GH patterns regulating expression of individual P-450 isoforms, it seems prudent to use species-specific GH (in this case, rGH) instead of the much more widely administered human and bovine GHs, which are not necessarily equally effective (MacGeoch et al., 1985; Morgan et al., 1985; Waxman et al., 1990a).
In spite of these drawbacks, earlier reports (see the introduction) proposed that the episodic or intermittent nature of the masculine GH profile was responsible for the phenotypic expression levels of CYP2A1, CYP2A2, CYP2C6, CYP2C7, CYP2C11, CYP2C12, CYP2C13, and CYP3A2 characteristic of the male rat. Subsequently, by using hypophysectomized rats with renaturalized circulating GH (Waxman et al., 1991; Shapiro et al., 1993), we were able to identify a minimum 2.5-h interpulse period, devoid of detectable GH, as the regulating signal attributed to the episodic or intermittent masculine GH profile.
Studies that examine the role of the masculine GH profile by administering one or two daily s.c. injections of pharmacologic doses of human or bovine GH (hGH, bGH) to hypophysectomized rats must base their conclusions on the ability of the exogenous hormone to reconstitute, to varying degrees, the male-like pattern of hepatic P-450 isoforms without reference to the physiologic plasma GH profiles. Even allowing for similar pharmacokinetics of rGH, bGH, and hGH, the mode of hormone administration (i.e., s.c. injections) precludes the possibility of reestablishing any profile resembling the natural masculine hormonal profile (Pampori et al., 1991a). This inability to restore the physiologic secretory patterns may explain why s.c. injections of ∼25 μg hGH/kg b.wt. were ineffective in inducing detectable concentrations of hepatic CYP2C11 protein in hypophysectomized rats (Morgan et al., 1985). It is likely that s.c. administration of this ∼25-μg hGH dose produced a plasma pulse that was even lower than that produced in the present study by the inductive 1-μg dose of the hormone. Similarly, twice daily s.c. injections of either ∼330 or ∼880 μg hGH/kg b.wt. were ineffective in restoring hepatic CYP3A2 protein levels in hypophysectomized rats, and were only minimally effective at the higher dose, in reducing CYP2A2 protein concentrations (Waxman et al., 1988). Last, daily s.c. injections of ∼500 to 600 μg hGH/kg b.wt. were observed to be ineffective in restoring male-like mRNA levels of hepatic CYP2C6 and 2C7 to hypophysectomized rats (Westin et al., 1990; Legraverend et al., 1992b).
Interestingly, those sexually dimorphic isoforms reported to be least responsive or even unresponsive to s.c. administered GH exhibited similar sensitivities to the renaturalized profiles (i.e., they were the most dependent on the higher, near normal pulse amplitudes for expression). In this regard, CYP2C7 appears to be the isoform least responsive to GH, requiring physiologic masculine as well as feminine (Pampori and Shapiro, 1996) plasma hormone profiles for normal sexually dimorphic expression5. Exhibiting a greater sensitivity to GH than CYP2C7, restoration of male-like expression levels of hepatic CYP2A1, CYP2A2, CYP2C6, and CYP3A2 still required considerable GH pulse amplitudes that could approach physiologic. In contrast, CYP2C11, the predominant isoform expressed in male liver (Morgan et al., 1985) and CYP2C12, the predominant isoform in female liver (MacGeoch et al., 1985) displayed a high sensitivity to GH requiring only 2 to 3% of the normal masculine and feminine (Pampori and Shapiro, 1996) profile, respectively, for the induction of substantial levels.
Clearly, the mechanism(s) mediating GH-dependent expression/suppression of some dozen constituent hepatic P-450 isoforms in the rat is complex. To illustrate, although a plasma GH concentration of 2 ng/ml or only 1% of the normal masculine pulse amplitude corresponds to half-maximal saturation of the membrane GH receptor (Leung et al., 1987), it would appear that GH induction of some isoforms (e.g., CYP2C7), as well as suppression of others (e.g., CYP2A2 and CYP3A2) is more than a matter of saturating the receptor. Moreover, it has been reported (Gebert et al., 1997) that signal transducer and activator of transcription 5b (Stat5b), a liver-expressed latent cytoplasmic transcription factor that is tyrosine phosphorylated by Janus kinase 2 (JAK2) and then undergoes nuclear translocation is selectively activated by the masculine episodic GH profile. Accordingly, it has been proposed (Waxman et al., 1995b) that this JAK2/Stat5b signal transduction pathway regulates expression of male-dependent, GH-responsive proteins (e.g., P-450s) in liver. Whereas this episodic growth hormone-driven intracellular mechanism might regulate some metabolic reactions, it would be difficult to explain how a subnormal GH pulse could selectively restore CYP2C11 transcription without affecting expression of other similar GH-dependent isoforms. That is, presumably a subnormal hormone pulse induces CYP2C11 expression by activating the transcription factor Stat5b. Why then aren't the other isoforms (e.g., CYP2A1 and CYP2C7) known to be responsive to the masculine GH profile also induced?
More than just responsive to nominal masculine GH pulse amplitudes, CYP2C11 exhibited a unique overexpression when exposed to several subnormal pulse amplitudes in the circulating profile6. Although not as dramatic as the 200% overexpression in transcript levels, CYP2C11 protein and dependent-catalytic activities also were increased to levels significantly above normal. Multi-P-450 [including CYP2C11 (Ryan and Levin, 1990)]-dependent hexobarbital hydroxylase levels also were suggestive of an above normal induction in response to subnormal pulse amplitudes (Shapiro et al., 1989). The reason(s) for this unique overexpression of CYP2C11 in response to subphysiologic pulse heights in the masculine GH profile is unknown, but the finding is consistent with our earlier reports with low dose MSG-treated rats exhibiting a 90% reduction in plasma masculine GH pulse amplitudes (Pampori and Shapiro, 1994b), hypophysectomized rats with renaturalized subnormal GH profiles (Pampori and Shapiro, 1994b), and subsequent studies in naturally occurring male dwarf rats exhibiting a >90% decline in their circulating GH pulse heights (Legraverend et al., 1992a).
The present findings demonstrate the importance of the pulse amplitudes in the circulating masculine GH profile as signals regulating the expression, repression, and even overexpression of sex-dependent hepatic P-450 isoforms in the rat7. Moreover, we have observed, for the first time, a differential dependence of the isoforms to varying pulse heights. However, the necessity of hepatocyte-recognizable pulses in the masculine GH profile does not preclude the requirement for the 2.5-h GH-devoid interpulse signal. Recently, we infused supraphysiologic pulses of rGH to hypophysectomized, thyroxine-replaced rats to produce masculine circulating GH profiles with pulses four times higher than normally found in intact males (N. A. Pampori and B. H. S., unpublished data). With a ∼1000 ng/ml pulse every 4 h, the decay rate of the hormone in the blood was retarded resulting in GH-devoid interpulse durations of <2.5 h and a commensurate 90% reduction (i.e., feminization) in hepatic CYP2C11 mRNA, protein, and dependent-testosterone 2α-hydroxylase. Clearly, to masculinize expression of the hepatic P-450s, the hepatocyte must be able to discriminate a GH profile characterized by a sufficient interpulse period devoid of GH but interrupted by recognizable pulses, albeit at a different height for each isoform.
In the rat, the gender-dependent hepatic isoforms of P-450 can be classified into three groups according to their dependence on GH for expression. The female-specific P-450s, containing the single isoform CYP2C12, exhibit the most restrictive dependence on GH. Expression is solely dependent on the continuous GH profile. The absence of GH (e.g., hypophysectomy) or exposure to the episodic masculine GH profile is completely ineffective in inducing CYP2C12 expression. Less restrictive are the male-specific isoforms (CYP2A2, CYP2C11, CYP2C13, and CYP3A2) characterized by their extreme sensitivity to the repressive effects of the continuous feminine GH profile. In contrast to the exclusive dependence of CYP2C12 on the feminine circulating profile, the male-specific isoforms are expressed, to varying degrees, in GH's absence or when exposed to their masculine GH profile. Exhibiting the least restrictive dependence on GH regulation are the female-predominant isoforms (e.g., CYP2A1, CYP2C6, CYP2C7, and CYP2E1) expressed in both sexes, but to a greater degree in female liver. (Curiously, there appear to be no male-predominant hepatic P-450s in the rat.) These female-predominant isoforms can be expressed by either the feminine or masculine GH profiles (the former being more effective) and in most cases, with the exception of CYP2C7, in the absence of the hormone. Adding another layer of complexity are our observations that each isoform exhibits a different sensitivity to the expressive and/or repressive signaling elements in both the masculine and feminine GH secretory profiles. Whether this prodigious permutation in GH dependence of the P-450s is beneficial or is even expressed under physiologic or pathologic conditions is unknown, but may be probed by determining if the different hormone-signaling elements regulate the expression and/or repression of each isoform by activating a different signal transduction pathway or by different “strands” in the signal transduction web.
Acknowledgments
We appreciate the generosity of Drs. Marika Rönnholm, Agneta Mode, and Jan-Åke Gustafsson in supplying the antibody to rat CYP2C12 and Dr. Stelvio M. Bandiera in supplying the antibody to rat CYP2C7. Materials used to assay rat GH were obtained through the National Hormone and Pituitary Program and Dr. A. F. Parlow. We also thank Alka Agrawal for excellent technical assistance.
Footnotes
-
Send reprint requests to: Bernard H. Shapiro, Laboratories of Biochemistry, School of Veterinary Medicine, University of Pennsylvania, 3800 Spruce St., Philadelphia, PA 19104-6084. E-mail:shapirob{at}vet.upenn.edu
-
↵1 This work was supported by National Institutes of Health Grants GM45758 and HD16358.
-
↵2 Current address: Department of Drug Metabolism, Merck Research Laboratories, P.O. Box 2000, RY80-A9, Rahway, NJ 07065-0900.
-
↵4 A comparison of the benefits and limitations of the growth hormone-deficiency models in studies of P-450 regulation has been addressed elsewhere (Shapiro et al., 1993, 1995).
-
↵5 Normal expression levels of hepatic CYP2C7 require both growth hormone and retinoids [i.e., vitamin A and its metabolites (Westin et al., 1997)]. Because retinoid metabolism is also growth hormone-dependent (Westin et al., 1997), it is possible that the subnormal replacement levels of the hormone were insufficient to generate the obligatory concentrations of retinoid metabolites needed for normal restoration of CYP2C7.
-
↵6 We reported a similar overexpression of hepatic female-dependent CYP2C12 in low dose MSG-treated female rats exhibiting a 75 to 85% permanent reduction in the continuous feminine growth hormone profile (Pampori and Shapiro, 1994a). This overexpression, however, was not observed in the hypophysectomized female rat infused with subphysiologic feminine plasma growth hormone profiles (Pampori and Shapiro, 1996).
-
↵7 The gender-dependent isoforms may not be the only P-450s regulated by the pulse amplitudes as the repressive effects of the masculine plasma growth hormone profile on phenobarbital induction of nonconstituent CYP2B1 and CYP2B2 are also directly proportional to the pulse amplitudes (Shapiro et al., 1994).
-
↵3 The terms sex-dependent, sex-predominant or dominant, and sex-specific are often used indiscriminately. We use sex- or gender-dependent to imply that expression levels are dependent on the existence of gender; sex- or gender-predominant indicates expression levels, regardless of magnitude, are consistently greater in one gender; and sex- or gender-specific implies that expression is basically restricted to only one gender.
- Abbreviations:
- CYP P-450
- cytochrome P-450
- GH
- growth hormone
- MSG
- monosodium glutamate
- rGH
- rat growth hormone
- hGH
- human growth hormone
- bGH
- bovine growth hormone
- Stat5b
- signal transducer and activator of transcription 5b
- JAK2
- Janus kinase 2
- Received June 28, 1999.
- Accepted September 14, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}