


Abstract
Organic cation transporters (OCT1–3) play an important role in renal elimination of many drugs. The goals of this study were to 1) identify a cell culture model which constitutively expressed OCT2 that could be used to study the characteristics and regulation of this transporter, and 2) to study the mechanisms by which xenobiotics and hormones regulate the activity of OCT2. We characterized the endogenous organic cation transporter (OCT) activity in Madin-Darby canine kidney (MDCK) cells. The activity was localized to the basolateral membrane and was pH and membrane potential-dependent. The uptake of the model organic cation, tetraethylammonium, was saturable (Km, 19.5 ± 4.6 μM;Vmax, 350 ± 19.4 pmol/mg of protein/10 min) and was inhibited by known OCT inhibitors (e.g., cimetidine and quinidine). A cDNA fragment (711 base pairs) isolated by reverse transcriptase-polymerase chain reaction (RT-PCR) was greater than 83% identical to OCT2 cDNAs from mammalian species; no OCT1 or OCT3 was detected by RT-PCR, suggesting that OCT2 may be the primary basolateral OCT in MDCK. OCT2 mRNA levels were increased significantly following exposure of MDCK to the steroid hormones, dexamethasone (2.0-fold), hydrocortisone (2.4-fold), and testosterone (1.8-fold) with comparable increases in activity. Other compounds tested, including the cytochrome P450 inducers, rifampicin, phenobarbital, and phenytoin, and the OCT substrates, verapamil and metformin, had no inducing effects. Collectively, these data indicate that MDCK can serve as a useful and convenient tool in screening candidate drugs for interaction with OCT2 and for studying the regulation of this transporter. Furthermore, our data demonstrate that steroid hormones induce the transcription of OCT2 in the kidney.
Under physiological pH, many clinically used drugs such as antihistamines, antiarrhythmics, antibiotics, β-adrenoceptor blocking agents, cytostatics, and sedatives are organic cations. Although structurally diverse, these compounds are secreted across renal epithelia via common active transport mechanisms that have been proposed as a two-step process: an electrogenic facilitated diffusion step via organic cation transporters (OCTs) at the basolateral membrane and an organic cation-proton exchange mechanism at the apical membrane that may be mediated by OCTN2 (Pritchard and Miller, 1993; Zhang et al., 1998a;Dresser et al., 2001). In recent years, significant progress has been made in understanding the molecular and cellular characteristics of the OCT group. Presently, three members of the OCT group, OCT1, OCT2, and OCT3, have been cloned from various species and functionally characterized in heterologous expression systems (Zhang et al., 1998a;Koepsell et al., 1999; Burckhardt and Wolff, 2000; Dresser et al., 2001).
Xenobiotics frequently regulate proteins involved in their disposition or detoxification. These proteins are also often under the regulatory influence of endogenous hormones. Although there are a number of examples of such regulation by xenobiotics or hormones, most of these concern drug-metabolizing enzymes, most notably the cytochrome P450 (CYP) family. Although P-glycoprotein (Pgp) has been demonstrated to be coordinately regulated with certain drug-metabolizing enzymes such as CYP3A4 by a wide array of xenobiotics (Thorgeirsson et al., 1991;Schuetz et al., 1996), the regulation of other drug transporters including OCTs by xenobiotics or hormones has received little attention. With the cloning and characterization of the OCTs, it is now possible to study mechanisms involved in their regulation. Recently,Urakami et al. (1999, 2000) reported that in vivo administration of testosterone increased the activity and mRNA levels of rOCT2 in the kidney whereas estradiol moderately reduced it, providing the first evidence of hormonal regulation of OCTs.
The goals of this study were to 1) identify a cell culture model that can be used to study the functional characteristics and mechanisms involved in the regulation of OCT2, and 2) to elucidate specific mechanisms by which xenobiotics or hormones regulate the activity of OCT2. In the present study, we characterized the localization, kinetics, electrogenicity, and pH dependence of OCT activity in Madin-Darby canine kidney (MDCK) cells. Using RT-PCR and sequence analysis, we determined that OCT2 was likely to be the primary OCT expressed in MDCK cells. Our data suggest that steroid hormones induce the transcription of OCT2 in MDCK cells.
Experimental Procedures
Materials.
Cell culture supplies were purchased from the Cell Culture Facility at the University of California (San Francisco, CA). Original stocks of MDCK cells were obtained from American Type Culture Collection (Manassas, VA).14C-Tetraethylammonium (14C-TEA) (55 and 2.40 mCi/mmol) was purchased from Moravek Biochemicals (Brea, CA) and PerkinElmer Life Science Products (Boston, MA), respectively. Unlabeled chemicals were supplied by either Sigma (St. Louis, MO) or Fisher Scientific (Pittsburgh, PA). Primers were synthesized by Invitrogen (Carlsbad, CA).
Cell Culture.
MDCK cells (passages 10–40) were grown in 100 × 20 mm plates in minimum Eagle's medium supplemented with 0.292 g/liter glutamine, 1.0 g/liter glucose, 2.2 g/liter NaHCO3, 100 IU/ml penicillin, 100 μg/ml streptomycin, and 10% (v/v) fetal bovine serum. Cells were maintained at 37°C in a humidified atmosphere (95% air, 5% CO2). Cultures were split 1:10 every 5 days. For transport studies, cells were polarized by growth on Transwell filters (0.4-μm pore size, 12-well plate; Costar, Cambridge, MA) at a confluent density for 7 days with regular media changes as described elsewhere (Mangravite et al., 2001).
14C-TEA Uptake Measurements.
All uptake experiments were performed on polarized cell monolayers. Briefly, monolayers were washed two times at room temperature in uptake buffer (145 mM NaCl, 3 mM KCl, 1 mM CaCl2, 0.5 mM MgCl2, 5 mM d-glucose, 5 mM HEPES, pH 7.4). Uptake was initiated by application of uptake buffer containing14C-TEA (with or without inhibitor) to either the apical or basolateral chambers. Blank uptake buffer was added to the opposite side. Although initial experiments showed that the transport rate of 14C-TEA was linear at room temperature for up to 60 min, incubation times of less than 10 min were used for all subsequent experiments to approximate the initial transport rate. Uptake was stopped by aspirating the reaction mixture and washing the filters three times with ice-cold buffer. Membranes were air dried, removed from plastic support, and radioactivity was counted using a Beckman scintillation counter (Beckman Coulter, Inc., Fullerton, CA). Data were normalized to protein count as determined by using the Bio-Rad protein assay (Bio-Rad, Hercules, CA).
Characterization of Endogenous OCT in MDCK Cells.
The OCT in MDCK cells was functionally localized to the basolateral and/or apical membrane grossly by uptake measurements. Trans-stimulation studies were also conducted as described previously (Zhang et al., 1998b). Similar to those described previously (Zevin et al., 1998), valinomycin (1 μM) dissolved in ethanol was added to the physiologic buffer (140 mM NaCl, 5.4 mM KCl, 1.8 mM CaCl2, 0.8 mM MgSO4, 5.0 mMd-glucose, and 25 mM HEPES) or to a depolarizing buffer (145 mM KCl, 1.8 mM CaCl2, 0.8 mM MgSO4, 5.0 mM d-glucose, and 25 mM HEPES) to investigate the effect of membrane potential on the OCT activity. For control cells, an equal amount of ethanol was added to the uptake medium. To study the effect of pH on OCT activity, the uptake of 14C-TEA was determined at pH 5.5, 6.5, 7.4, and 8.5, respectively.
Kinetic and Inhibition Studies.
Transport kinetics were characterized by measuring uptake of 14C-TEA (5 μM) at 10 min in the presence of varying concentrations of unlabeled TEA (0–500 μM) with or without the OCT inhibitor quinidine (200 μM). The inhibitory effects of selected compounds on OCT activity were also examined by measuring uptake of 14C-TEA in the presence of varying concentrations (0.2–500 μM) of compounds. Data were fit as described under Data Analysis.
RT-PCR and Sequence Analysis.
Total RNA was extracted from MDCK cells using TRIzol reagent (Invitrogen). For RT-PCR analysis, 1.0-μg total RNA was reverse-transcribed and amplified using SuperScript one-step RT-PCR with Platinum Taq (Invitrogen) under the following conditions: 1) initial cycle of 50°C for 30 min and 94°C for 2 min; 2) 25 to 38 cycles of 94°C for 15 s, 57°C for 30 s and 72°C for 1.5 min; and 3) extension cycle of 72°C for 15 min. Because dog OCT has not been cloned, the OCT primers used in this study were designed using consensus regions from OCT1, OCT2, or OCT3 from mammalian species (Table 1). The mRNA level of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) in MDCK cells was also determined as an internal standard for OCT mRNA quantification. For OCT sequence analysis, 38 PCR cycles were used to obtain sufficient RT-PCR product. However, the number of PCR cycles for mRNA quantification was empirically determined with different amounts of total RNA from the same sample and with the same amount of total RNA from different samples to ensure that amplification did not reach saturation and that products would be reflective of mRNA levels in cells. In most cases, 28 to 33 cycles were optimal for OCT2, whereas 25 to 30 cycles were optimal for GAPDH. ScnImage software (Scion Corporation, Frederick, MD) was employed to quantify mRNA levels.
Oligonucleotide primers for OCT and GAPDH in RT-PCR analysis
RT-PCR products were electrophoresed through 1% agarose gels. For sequence analysis, size-selected DNA fragments were extracted, subcloned into pGEM-T (Promega, Madison, WI), and sequenced by the Biomolecular Resource Center (University of California San Francisco). Sequences were compared with all published OCT cDNA sequences using BLAST (http://www.ncbi.nlm.nih.gov/BLAST/).
Regulation Studies.
Regulation was studied by treating monolayers for 72 h with xenobiotic or hormone prior to uptake of14C-TEA (50 μM). Compounds were used at a concentration of 10 μM unless otherwise noted. Controls contained an equivalent volume of solvent. After 72 h, cells were washed three times with PBS, and 14C-TEA uptake was measured. For those compounds with significant effects,trans-stimulation studies were carried out as described previously (Zhang et al., 1998b).
Data Analysis.
Each data point was determined in duplicate or triplicate for each experiment unless indicated. The data are presented as mean ± standard deviation unless indicated. All experiments were repeated at least once using a different cell passage. To obtain the Km andVmax in the kinetic studies, the data were fit to the Michaelis-Menten equation: V =Vmax ·S/(Km + S), where Vmax is the maximum transport rate, Km is the Michaelis-Menten constant at which the transport rate (V) is one-half theVmax, and S is the concentration of TEA. Kaleidagraph version 3.0 (Abelbeck/Synergy, Reading, PA) was used to fit the data. IC50 values were estimated by fitting the data to the following equation: V =V0/[1 + (I/IC50)n] where V is the uptake rate of 14C-TEA in the presence of inhibitor, V0 is the uptake rate of 14C-TEA in the absence of inhibitor, I is the concentration of inhibitor, andn is the Hill coefficient. Unpaired Student's ttest was applied to analyze data, when appropriate. A pvalue of <0.05 was considered statistically significant.
Results
Characterization of Endogenous OCT in MDCK Cells.
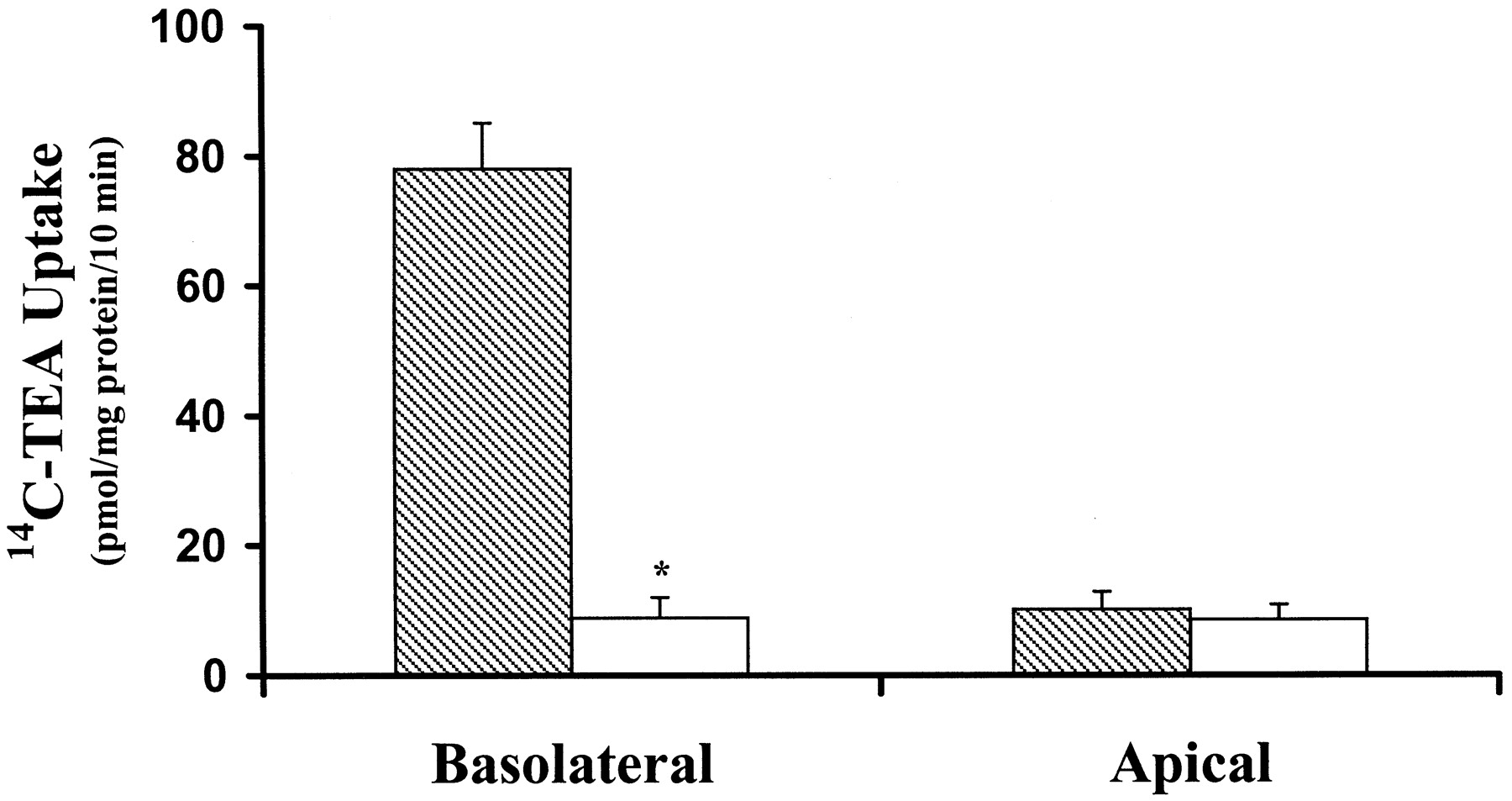
The transport of the model organic cation, TEA, was detectable across both the basolateral and apical membranes of MDCK cells. However, the uptake of 14C-TEA was considerably greater across the basolateral membrane (78.3 ± 7.1 pmol/mg of protein/10 min versus 10.1 ± 2.7 pmol/mg of protein/10 min across the apical membrane,p < 0.05), and was inhibitable by quinidine (Fig.1). Basolateral uptake of14C-TEA was also significantly inhibited by the organic cations (500 μM) N-methylnicotinamide, hemicholinium, quinidine, guanidine, cimetidine, and TEA but not byd-glucose or the organic anion, PAH (Fig.2). These data suggest that an OCT is present on the basolateral membrane of MDCK cells.

Functional localization of endogenous OCT activity in MDCK cells. Uptake of 14C-TEA (5 μM) across the basolateral or apical membrane was measured in the absence (filled bars) or presence (open bars) of quinidine (200 μM). Data are presented as the mean ± S.D. (n = 3). Asterisks indicate significant difference from basolateral uptake in the absence of quinidine (p < 0.05).

Inhibition of 14C-TEA uptake across the basolateral membranes of MDCK cells. Uptake of 14C-TEA (5 μM) was measured in the absence (control) or presence of various compounds (500 μM). NMN, N-methylnicotinamide; HC-3, hemicholinium. Data represent the mean ± S.D. (n = 3). Asterisks indicate significant difference from control (p < 0.05).
In cells preloaded with 250 μM unlabeled TEA, a significanttrans-stimulation of 14C-TEA uptake was observed (29%), whereas the same concentrations of preloaded cimetidine and decynium-22 were found to significantly inhibit14C-TEA uptake (40 and 86%, respectively; Fig.3), suggesting that cimetidine and decynium are trans-inhibitors, or possibly slowly dissociating cis-inhibitors, of the basolateral OCT in MDCK cells. These effects of preloaded TEA, cimetidine, and decynium-22 were previously observed in HeLa cells transiently transfected with an organic cation transporter, hOCT1 (Zhang et al., 1998).

Trans-stimulation or inhibition of14C-TEA uptake across the basolateral membrane of MDCK cells. Cells were pretreated with PBS (control) or PBS containing 250 μM unlabeled compound for 20 min at 37°C before 5 min uptake of14C-TEA (5 μM). Data represent the mean ± S.E. (n = 3). Asterisks indicate significant difference from control (p < 0.05).
14C-TEA uptake across the basolateral membrane was linear for at least 60 min and was shown to be significantly inhibited by unlabeled cimetidine (250 μM) at all points during this time period (data not shown). The inhibitable, carrier-mediated uptake of 14C-TEA was sensitive to membrane potential. When the inside negative potential of MDCK cells was abolished with valinomycin (1 μM) and a high extracellular K+concentration, the uptake was reduced by 50% from 37.2 ± 1.5 to 18.7 ± 0.7 pmol/mg of protein/3 min (p < 0.05). In contrast, hyperpolarizing the cells led to a slight but not statistically significant increase (18%) in the uptake (Fig.4A). The effect of extracellular pH on the uptake was also evaluated. 14C-TEA uptake was stimulated with increasing pH under normal membrane potential (Fig.4B). These data suggest that the basolateral transporter is both pH and potential-sensitive.

Effect of membrane potential (A) and extracellular pH (B) on 14C-TEA (5 μM) uptake across the basolateral membrane of MDCK cells. A, cells were incubated for 20 min with PBS (control) or valinomycin (1 μM) in depolarizing or physiologic buffer prior to uptake of 14C-TEA; B, 14C-TEA uptake was measured in uptake buffer at pH 5.5, 6.5, 7.4 (control), or 8.5. Data are presented as mean ± S.D. (n = 3). Asterisks indicate significant difference from control (p < 0.05).
Kinetics of TEA Transport across the Basolateral Membrane of MDCK Cells.
To determine whether TEA uptake across the basolateral membrane of MDCK cells was saturable, initial rates (10 min) of14C-TEA (5 μM) uptake were measured over a range of TEA concentrations (0–500 μM) (Fig.5A). Kinetic behavior, evaluated by Eadie-Hofstee analysis (Fig. 5B), suggests that a single transporter is involved in the basolateral uptake of TEA into MDCK cells. The single-component Michaelis-Menten model [V =Vmax ·S/(Km + S)] was then iteratively fit to the untransformed kinetic data, and a good fit was achieved. Computer modeling derived kinetic parameters were 19.5 ± 4.6 μM (Km) and 350 ± 19.4 pmol/mg of protein/10 min (Vmax), respectively.

Saturation of 14C-TEA uptake rate across the basolateral membrane of MDCK cells. 14C-TEA (5 μM) uptake in the presence of increasing concentrations of unlabeled TEA (0–500 μM) was measured in the absence (active transport + passive diffusion) or presence (passive diffusion) of quinidine (200 μM). Data points represent active transport (“absence − presence”) and are graphed as hyperbolic (A) or Eadie-Hofstee (B) plots. Data were fit by nonlinear regression to the equationV = Vmax ·S/(Km + S).
RT-PCR and Sequence Analysis.
Primers designed to consensus sequences of each OCT were used in RT-PCR to determine which OCT group member was present in MDCK (Table 1). A cDNA fragment was amplified with the primers designed for OCT2. The cDNA fragment was further sequenced as a 711-base pair-long fragment and compared with other published OCT cDNA sequences using BLAST software. The fragment shares at least 83% identity with all the OCT2 cDNA sequences but shares a very low identity (<11%) with those of OCT1 and OCT3, suggesting that a fragment of the canine ortholog of OCT2 had been amplified. This fragment has been reported to GenBank with an accession number ofAY028623. No RT-PCR product was detected with the primers designed for OCT1 and OCT3 (data not shown). This suggests that neither OCT1 nor OCT3 are expressed in MDCK cells. Alternatively but less likely, our primers designed to those OCTs of other species may be unable to detect canine OCT1 and OCT3 mRNA in MDCK cells.
Regulation Studies.
We determined whether estradiol and testosterone as well as several known inducers of drug-metabolizing enzymes and/or the drug transporter Pgp could regulate the activity and expression of endogenous OCT2 in MDCK cells. Among the selected drugs and hormones at the concentration tested, dexamethasone, testosterone, and hydrocortisone significantly enhanced the basolateral14C-TEA uptake (Fig.6), whereas estradiol and phenytoin slightly decreased 14C-TEA uptake by MDCK cells. Neither a trans-stimulation nor atrans-inhibition effect on 14C-TEA uptake was observed with these five compounds (data not shown), excluding trans-effects as a mechanism of their regulatory effects on 14C-TEA uptake. On the other hand, incubation of cells with phenobarbital, verapamil, rifampicin, thyroid T3, and metformin for 72 h did not result in any significant change in 14C-TEA uptake. Dexamethasone, hydrocortisone, and testosterone increased the basolateral14C-TEA uptake in a concentration-dependent manner (Fig. 7). Hydrocortisone (50 μM), which had the most apparent regulatory effect among the three compounds, more than doubled the 14C-TEA uptake (180 ± 15 versus 410 ± 0.67 pmol/mg of protein/10 min). There was no relationship between 14C-TEA uptake and estradiol concentrations (0.1–10 μM). It was not possible to determine the concentration-dependent effects of phenytoin on OCT activity because this compound inhibited proliferation of cells in this study.
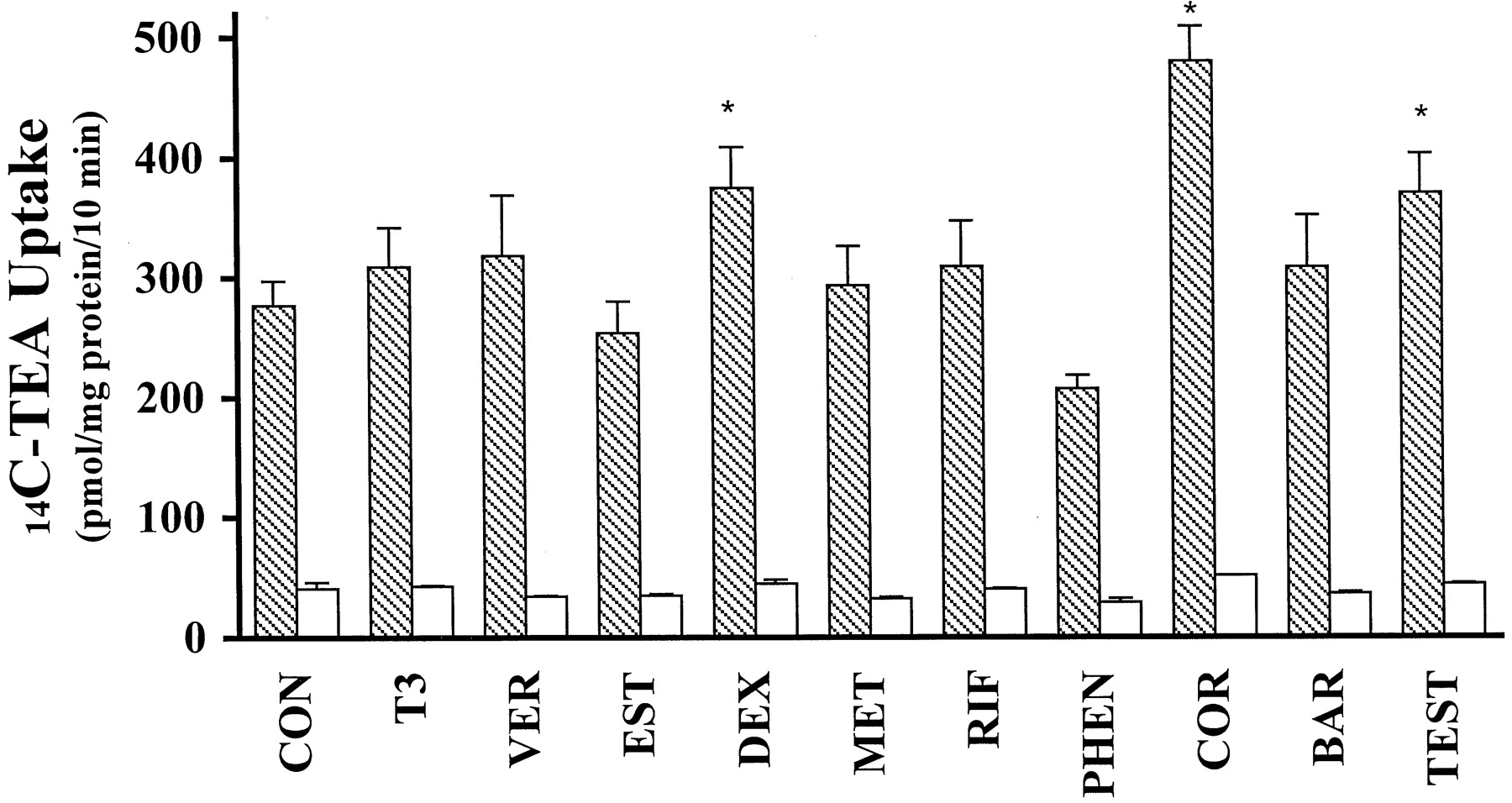
Effects of xenobiotics and hormones on14C-TEA (50 μM) uptake across the basolateral membrane of MDCK cells. Cells were pretreated for 72 h with medium (control) or medium containing 10 μM indicated compound [except thyroid hormone (T3; 100 nM), estradiol (EST; 50 nM), and testosterone (TEST; 1 μM)] prior to the uptake of 14C-TEA (50 μM). Uptake was measured in the absence (filled bars) or presence (open bars) of quinidine (200 μM). Con, control; VER, verapamil; DEX, dexamethasone; MET, metformin; RIF, rifampicin; PHEN, phenytoin; COR, hydrocortisone; BAR, phenobarbital. Data are presented as mean ± S.D. (n = 3). Asterisks indicate significant difference from control (p < 0.05).

Concentration dependence of inductive effects of steroid hormones on 14C-TEA uptake in MDCK cells. Polarized cells were incubated for 72 h in the absence (control) or presence of varying concentrations (0.1–50 μM) of dexamethasone (A), hydrocortisone (B), or testosterone (C). 14C-TEA uptake (50 μM) was measured in the absence (filled bars) or presence (open bars) of quinidine (200 μM). Data are presented as mean ± S.D. (n = 2).
It is possible that the regulatory effects of a compound on OCT2 transport activity may be obscured if the compound also inhibits14C-TEA transport. Therefore, we performed inhibition studies with the compounds shown to regulate organic cation transport activity in MDCK along with compounds known to inhibit OCT transport (Fig. 2). Hydrocortisone, dexamethasone, testosterone, estradiol, and verapamil significantly inhibited the basolateral14C-TEA uptake in a concentration-dependent fashion. The apparent IC50 values are shown in Table 2.
Potencies of various compounds in inhibiting 14C-TEA uptake across the basolateral membrane of MDCK cells
To determine the mechanism responsible for the hormonal or drug-induced regulation of OCT activity in MDCK, we determined the expression of OCT2 mRNA transcripts using RT-PCR (Fig.8). Consistent with its effect on OCT activity, hydrocortisone increased the amount of OCT2 mRNA by a factor of 2.4. Moreover, dexamethasone and testosterone also significantly enhanced OCT2 mRNA expression, whereas no difference in the level of OCT2 mRNA was detected in cells exposed to the other compounds. None of the compounds tested had an apparent effect on GAPDH mRNA expression.

Effects of xenobiotics and hormones on mRNA levels of OCT2 in MDCK cells. Top panel, RT-PCR analysis of canine OCT2 and GAPDH mRNA of MDCK cells treated for 72 h with the indicated compounds. From left to right, lanes 1 and 14 are markers, lanes 12 and 13 are controls, and lanes 2 through 11 correspond to treated samples in the order indicated in the bottom panel (see Fig. 6 legend for abbreviations). RT-PCR samples were electrophoresed through a 1% agarose gel. Bottom panel, density of electrophoresis bands was analyzed using ScnImage and normalized to internal control (GAPDH band). Data represent mean ± S.E. (n = 2).
Discussion
In this study, we characterized endogenous OCT activity in MDCK cells, a polarized epithelial cell line derived from canine kidney and examined regulation of this activity by xenobiotics and steroid hormones. Carrier-mediated uptake of TEA was observed on the basolateral membrane (Fig. 1) and was inhibitable by a broad range of organic cations but not by d-glucose, a sugar, or PAH, an organic anion (Fig. 2). This organic cation-specific transport activity was saturable, potential-dependent, and affected by extracellular pH. In addition, TEA uptake could be trans-stimulated by unlabeled TEA and inhibited by preloaded cimetidine or decynium-22 (Fig. 3). These transport properties are characteristic of the OCT group of transporters. Moreover, PCR amplification of cDNA from MDCK cells resulted in a single OCT product, which was at least 83% identical to all reported OCT2 cDNAs, suggesting that the organic cation transport system characterized in MDCK cells may be the canine ortholog of OCT2.
OCTs are thought to localize to the basolateral membrane of epithelial cells, participating in the first step of a two-part process of cation transport that results in active secretion of organic cations in the kidney (Zhang et al., 1998a). Although there is some evidence to the contrary (Gorboulev et al., 1997; Gründemann et al., 1997), recent studies using heterologous expression systems and direct staining of kidney sections with specific antibodies suggest both rOCT1 and rOCT2 are localized to the basolateral membrane (Urakami et al., 1998; Sweet and Pritchard, 1999; Karbach et al., 2000; Sweet et al., 2000). Our data localizing electrogenic OCT activity to the basolateral membrane of MDCK cells also supports this two-step model of active secretion.
Transport of TEA across the basolateral membrane of MDCK cells was concentration-dependent and saturable with an apparentKm of 19.5 μM and aVmax of 350 pmol/mg of protein/10 min. This apparent affinity is somewhat higher than those observed for OCT2 homologs of rat, pig, and human (reviewed by Dresser et al., 2001), suggesting species-related differences in OCT2 characteristics. However, it is possible that other OCT members, which may contribute to OCT activity in MDCK cells, might not have been detected by the RT-PCR methods that we used.
Recent studies suggested that the activity and mRNA level of rOCT2 in rat kidney are increased by testosterone and moderately reduced by estradiol (Urakami et al., 1999, 2000). Consistent with these results, we observed that both the activity and the mRNA level of OCT2 in MDCK cells are increased by testosterone and slightly reduced by estradiol. Furthermore, we observed that the steroid hormones, dexamethasone and hydrocortisone, also increased OCT2 activity and mRNA level (Fig. 6 and8). These data suggest that OCT2 can be transcriptionally regulated by steroid hormones. Hormonal regulation of OCT2 activity may be an underlying mechanism of the observed gender difference in renal disposition of amantadine, an organic cation, in humans (Wong et al., 1995). MDCK cells have been used for heterologous expression of OCT1 and OCT2 from rat (Urakami et al., 1998). Our data indicate that MDCK cells can be used as a convenient model to further study the regulation of OCT2 as well as to screen candidate drugs for interaction with OCT2. LLC-PK1 cells an original cell line is already available for screening drugs interacting with OCT2 (Dudley et al., 2000). However, the OCT2 in LLC-PK1 cells (OCT2p) is localized to brush membrane (Gründemann et al., 1997), which may be different from the basolateral membrane-located OCT2 in MDCK cells. The OCT2p has been proposed to be another member of the OCT group (Urakami et al., 1998). Potential limitations may still exist when MDCK cell line is used for the above purposes. For example, this is a cell line with all of the limitations of cell lines in terms of alteration of function over time in culture. Moreover, since MDCK cells are derived from the distal tubule of kidney, the transport mechanisms may be different from those in proximal tubule cells that are primarily responsible for drug active secretion in the kidney.
A variety of xenobiotics regulate proteins involved in their physiologic distribution and/or detoxification, most notably the CYP family. Because OCTs are important in the active secretion of cationic xenobiotics, we were interested in examining the potential effect of several compounds on OCT2 activity and mRNA levels in MDCK cells. We chose three compounds (phenytoin, dexamethasone, and rifampicin) documented to coordinately regulate levels of CYP3A4 and Pgp (Schuetz et al., 1996), two compounds that are substrates of OCTs (verapamil and metformin) (Zhang et al., 1998b; and M. Dresser, G. Xiao, M. Leabman Kaushal, A. Gray, and K. Giacomini, unpublished data), and phenobarbital because there is a cis-element for barbiturate-induced expression in the 5′-upstream region of the human OCT2 gene (Gründemann and Schömig, 2000). Of the non-steroids, only phenytoin affected OCT activity in MDCK cells (Fig.6). This effect is probably due to the cytotoxic effect of phenytoin because MDCK cells cultured in medium containing phenytoin (10 μM) did not proliferate well.
In summary, we have determined that OCT activity in MDCK cells appears to be due to the expression of an OCT2 ortholog. MDCK cells provide a useful tool for examining the transcriptional regulation of OCT2 by xenobiotics and hormones. In this study, we observed that OCT2 in MDCK cells was transcriptionally regulated by steroid hormones.
Footnotes
-
↵1 Current address: Sugen Inc., 230 East Grand Avenue, South San Francisco, CA 94080-4811.
-
This work was supported by a Grant GM 36780 from the National Institutes of Health. L.M.M. was also supported by Pharmaceutical Chemistry, Pharmacology and Toxicology Training Grant GM 07175. Y.S. is a recipient of a Merck Sharp & Dohme International Fellowship in Clinical Pharmacology.
- Abbreviations:
- OCT
- organic cation transporter
- rOCT
- rat organic cation transporter
- CYP
- cytochrome P-450
- Pgp
- P-glycoprotein
- MDCK
- Madin-Darby canine kidney
- RT-PCR
- reverse transcriptase-polymerase chain reaction
- TEA
- tetraethylammonium
- GAPDH
- glyceraldehyde-3-phosphate dehydrogenase
- PAH
- p-aminohippuric acid
- PBS
- phosphate-buffered saline
- Received April 27, 2001.
- Accepted June 29, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}