


Abstract
Gugulipid is an extract of the guggul tree, Commiphora mukul, that is used to treat hyperlipidemia in humans. The lipid-lowering activity is found in the stereoisomers and plant sterols Z-guggulsterone and E-guggulsterone. The molecular basis for the lipid-lowering action of guggulsterone has been suggested to be antagonism of the farnesoid X receptor, a member of the nuclear receptor superfamily of ligand-activated transcription factors. To determine whether guggulsterone has the ability to function as an agonist of other nuclear receptor family members, we screened a panel of these proteins for their ability to transactivate reporter genes. Here, we show that guggulsterones activate the estrogen receptor α isoform, progesterone receptor, and pregnane X receptor. Concentration-response analysis using reporter gene assays indicate that guggulsterones activate these three receptors with EC50 values in the low micromolar range. Furthermore, we show that guggulsterone-mediated activation of the pregnane X receptor induces the expression of CYP3A genes in both rodent and human hepatocytes. Protein interaction assays indicate that guggulsterones interact directly with pregnane X receptor, thereby modulating interaction with protein cofactors. We introduce a novel method to screen herbal remedies for their ability to activate pregnane X receptor. Pregnane X receptor activation is known to cause herb-drug interactions, and our data suggest that gugulipid therapy should be used cautiously in patients taking prescription medications that are metabolized by CYP3A family members. Moreover, our data suggest the need for additional studies of guggulsterones agonist activity against estrogen receptor α isoform and the progesterone receptor.
Among the many herbal remedies available over-the-counter, gugulipids are made from the gum resin of a tree from India called Commiphora mukul. Gugulipid therapy has been used in Ayurvedic medicine to treat hyperlipidemia for nearly 3000 years (Dev, 1999). Experiments using animals and human clinical trials indicate that treatment with gugulipid produces favorable serum lipid profiles (Kuppurajan et al., 1978; Baldwa et al., 1981; Singh et al., 1990). The active ingredient in gugulipid is the cis- and trans-stereoisomers of 4,17(20)-pregnadiene-3,16-dione, referred to as E-guggulsterone and Z-guggulsterone, respectively (Beg et al., 1996).
Recent experiments suggest that guggulsterones lower hepatic cholesterol through antagonism of the farnesoid X receptor (FXR; NR1H4) nuclear receptor superfamily member. Induction of FXR-target gene expression by FXR ligands is abolished in mice when guggulsterones are coadministered (Urizar et al., 2002; Wu et al., 2002). Other research shows that guggulsterones enhance the action of specific FXR agonists toward bile salt excretory protein gene expression in vivo and may function as a selective FXR modulator that positively regulates expression of a subset of FXR-target genes (Cui et al., 2003). A double blind, randomized, placebocontrolled study using standardized guggul extract in hypercholesterolemic patients showed that short-term (8-week) treatment did not significantly lower serum cholesterol (Szapary et al., 2003). However, cholesterol-feeding studies show that guggulsterone treatment produces significantly lower hepatic cholesterol levels in wild-type mice compared with FXR knockout littermates (Urizar et al., 2002).
Although guggulsterones modulate FXR activity, biochemical data suggest that they also may function as pregnane X receptor (PXR; NR1I2) agonists (Urizar et al., 2002; Wu et al., 2002). This is important because patients taking prescription drugs that self-medicate with gugulipid herbal remedies may induce herb-drug interactions by upregulating CYP3A4 expression (Willson and Kliewer, 2002). The identification of the active ingredient in the herbal remedy St. John's wort as a PXR-activator exemplifies the prototypical herb-drug interaction paradigm (Moore et al., 2000). People taking St. John's wort are at risk of altering the serum levels of coadministered medications that are CYP3A4 substrates, such as cyclosporin (Ruschitzka et al., 2000). It is therefore important to determine whether herbal remedies modulate the expression of CYP3A4 through PXR activation.
PXR is activated by a variety of drugs, xenobiotics, bile acids, and bile acid precursors (Bertilsson et al., 1998; Blumberg et al., 1998; Lehmann et al., 1998; Goodwin et al., 1999; Staudinger et al., 2001b; Xie et al., 2001; Dussault et al., 2003; Goodwin et al., 2003). PXR knockout mouse models show that PXR functions as a master regulator of the inducible expression of CYP3A family members (Xie et al., 2000; Staudinger et al., 2001a,b). The CYP3A4 gene encodes the most abundant drug-metabolizing cytochrome P450 in human liver and is responsible for the biotransformation of nearly 60% of all clinically prescribed drugs (Maurel, 1996; Guengerich, 1999). The administration of PXR activators should thus be avoided in patients prescribed medications that are metabolized by CYP3A4.
The CYP3A4 gene has two PXR/RXR heterodimer binding sites separated by approximately 8000 base pairs, and an hepatocyte nuclear factor 4α (HNF4α; NR2A1) binding site in its promoter (Goodwin et al., 1999). Removal of the intervening DNA sequences places these PXR-response elements in tandem and produces a highly responsive xenobiotic response element (XREM). This artificial CYP3A4 promoter has been placed upstream of the luciferase reporter gene and is termed XREM-LUC (Goodwin et al., 1999). The XREM-LUC reporter gene has been used to assay activation of human and rodent PXR in CV-1 cells using transient transfection (Goodwin et al., 1999). Overexpression of HNF4α confers maximal transactivation capacity to XREM-LUC in nonhepatic cell lines (Tirona et al., 2003).
Here, we show that guggulsterones function as agonists for at least three nuclear receptor proteins: the estrogen receptor α isoform (ERα; NR3A1), the progesterone receptor (PR; NR3C3), and the PXR (NR1I2). We show that gugulipid and guggulsterones induce CYP3A gene expression both in cell-based assays and in cultured hepatocytes through PXR activation. In vitro assays suggest a direct interaction of guggulsterone with the PXR ligand-binding domain. We introduce a novel high-throughput assay that uses the secreted placental alkaline phosphatase (SPAP) gene under the control of XREM for use as a tool in screening herbal remedies for PXR activation. These data suggest that patients taking prescription medications metabolized by the CYP3A4 pathway should avoid gugulipid therapy. Our data suggest that additional studies of guggulsterones' agonist activity against the ERα and PR are warranted.
Materials and Methods
Plasmids. The full-length mammalian expression vectors encoding human PXR (Lehmann et al., 1998) and mouse PXR (Kliewer et al., 1998) were described previously. The pGEX-PXR ligand-binding domain (LBD) fusion protein also was described previously (Kliewer et al., 1998). The mammalian expression vector encoding human HNF4α was described previously (Wisely et al., 2002). The steroid receptor activator 1 (SRC-1) expression vector was described previously (Onate et al., 1995). The XREM-SPAP reporter gene was constructed using PCR primers to amplify XREM and insert it into the pTAL-SEAP (BD Biosciences Clontech, Palo Alto, CA) using KpnI and BglII restriction sites.
Nuclear Receptor Panel Screen. CV-1 cells were maintained and transiently transfected as described previously (Moore and Kliewer, 2000). The following receptors were transfected as Gal4-LBD chimera constructs (PPARα, PPARδ, PPARγ, retinoic acid receptor α, vitamin D receptor, TRα, and TRβ) with a UAS(5X)-tk-Luciferase reporter as described previously (Parks et al., 1999). Liver X receptors α and β and FXR also were transfected as Gal4-LBD chimera constructs as described above, except in these transfections an expression plasmid encoding the coactivator SRC-1 was included as described previously (Moore et al., 2002). The remaining receptors (glucocorticoid receptor, PR, androgen receptor, mineralocorticoid receptor, RXRα, PXR, ERα, and ERβ) were tested using full-length receptor expression constructs and the reporter genes used were pLM253-MMTV-Luciferase for GR, PR, AR, and MR, pCRBPII-tk-SPAP for RXR, XREM-Luciferase for PXR, and ERE(2X)-tk-SPAP for ERα and ERβ; Goodwin et al., 2000). The ERα and ERβ assays were run in T47D cells instead of CV-1. The cells were treated with 10 μM Z-guggulsterone.
Maintenance and Treatment of PXR Knockout and Wild-Type Mouse Populations. Generation of the PXR knockout (PXR-KO) mice was described previously (Staudinger et al., 2001b). Adult male wild-type mice and PXR-KO mice were maintained on standard laboratory chow and were allowed food and water ad libitum. All animal experiments described in this manuscript were conducted in accordance with accepted standards of humane animal care.
Gugulipid Extract Preparation. A commercial capsule preparation of gugulipid extract was obtained from the General Nutrition Corporation (Pittsburgh, PA). Gugulipid from one capsule (600 mg) was transferred to an amber glass vial and extracted with 1 ml of absolute ethyl alcohol (Aaper Alcohol and Chemical, Shelbyville, KY) by gentle agitation for 30 min. The extract was centrifuged at 300g for 5 min, and the alcohol layer was transferred to a clean centrifuge tube and recentrifuged at 425g for 30 min. Ethanol extracts were decanted and stored in amber glass vials at –20°C.
RNA Isolation for Northern Blot and Quantitative-Polymerase Chain Reaction (Q-PCR) Analysis. Total RNA was isolated from cultured hepatocytes using a commercially available reagent (TRIzol, Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. For Northern blot analysis, total RNA (15 μg) was resolved on a 1% agarose/2.2 M formaldehyde denaturing gel and transferred to nylon membrane (Hybond N+, Amersham Biosciences Inc., Piscataway, NJ). Blots were hybridized with 32P-labeled cDNAs corresponding to the cDNA sequences for mouse Cyp3a11 (bases 69–1609; GenBank NM 007818), human CYP3A4 (bases 790–1322; GenBank NM 017460), and β-actin (BD Biosciences Clontech). For Q-PCR analysis, total RNA was further purified using RNA-EZ columns (QIAGEN, Valencia, CA) after DNaseI (Sigma-Aldrich, St. Louis, MO) treatment.
PXR Transactivation Assays. CV-1 Cells were maintained in DMEM containing 10% fetal bovine serum supplemented with l-glutamine and antibiotics. On the first day, cells were plated in 96-well plates at a density of 20,000 cells/well. After 24 h, cells were transfected using LipofectAMINE (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. The PXR transactivation assays used transfection mixtures containing 110 ng of total DNA per well and were composed of XREM-LUC or XREM-SPAP (25 ng), pSG5-PXR (5 ng), pSG5-HNF4α (3 ng), pSV-β-galactosidase (40 ng), and pBluescript (40 ng). After 6 h, transfection mixtures were removed and replaced with phenol red free DMEM/F-12 (Invitrogen) containing 10% charcoal-stripped delipidated fetal bovine serum (Sigma-Aldrich). Transfected cells were treated 24 h later for 48 h with PCN (10 μM), rifampicin (10 μM), ethanol extracts of gugulipid, or Z-guggulsterone (1, 5, 10, and 20 μM). Luciferase and β-galactosidase reporter gene activities were determined using the Dual-Light kit (Applied Biosystems, Foster City, CA). After heat inactivation (65°C for 20 min), alkaline phosphatase activity in cell culture media was determined using the SEAP reporter gene assay kit (Roche Applied Science, Indianapolis, IN).
Hepatocyte Isolation, Maintenance, and Treatment. Unless otherwise specified, all chemicals and reagents were purchase from Sigma-Aldrich. Mouse hepatocytes were isolated using a modification of the two-step perfusion as described previously (LeCluyse et al., 1996). Briefly, the liver was cleared with wash medium (0.15 M sodium chloride, 5 mM potassium chloride, 0.5 mM EGTA, 50 mM HEPES, and 2 mM glucose) at a flow rate of 16 ml/min for 4 min. The liver was digested for 8 min at a flow rate of 8 ml/min with digestion medium containing 0.5 M sodium chloride, 5 mM potassium chloride, 6 mM calcium chloride, 100 mM HEPES, 0.07% type I collagenase (Invitrogen) and 2 mM glucose. The suspension was filtered through a 100-μm cell strainer (BD Biosciences Clontech). The cells were pelleted at 129g for 5 min at 4°C and washed twice in DMEM supplemented with 5% fetal bovine serum, 100 nM dexamethasone, 1 μM insulin, and antibiotics. Cell viability was determined using trypan blue staining. Generally, 70 to 90% viability was achieved. Mouse hepatocytes were plated in 12-well collagen-coated cell culture plates (BD Biosciences Clontech) at a density of 1 × 105 live cells/ml. After 2 h, DMEM was aspirated and replaced with 1 ml of William's medium E (Invitrogen) supplemented with 100 nM dexamethasone, antibiotics, and insulin-transferrin-selenium + (ITS+) (Invitrogen).
Human hepatocytes were purchased from In Vitro Technologies (Baltimore, MD) and were cultured as described previously (Moore et al., 2000). Two days postplating, hepatocytes were treated for 48 h with either purified guggulsterones (10 μM) (Steraloids, Newport, RI), dexamethasone (10 μM), gugulipid (1:4000 diluted in ethanol), rifampicin (10 μM), or PCN (10 μM) as described in the legend for Fig. 3. Unless otherwise specified, all compounds were added to the culture medium as 1000× stocks in DMSO.

Guggulsterone and gugulipid induces the expression of CYP3A family members in cultured hepatocytes through activation of PXR. A, purified RNA from wild-type or PXR-KO mouse hepatocytes treated with vehicle (0.1% DMSO), PCN (10 μM), or three doses of E-guggulsterone (1, 10, and 20 μM) was subjected to Q-PCR analysis using probe sets specific for Cyp3a11 and β-actin. The data are expressed as fold-induction relative to the wild-type vehicle experimental group and are reported as the mean ± S.E. of three independent experiments. Asterisks indicate statistically significant difference in Cyp3a11 gene expression levels between wild-type and PXR knockout hepatocytes within the respective experimental group (P < 0.05). B, total RNA (15 μg/lane) isolated from cultured human hepatocytes treated with vehicle (0.1% ethanol), E-guggulsterone (10 μM), gugulipid extract (1:16,000-fold dilution in ethanol), or rifampicin (10 μM) was resolved on a 3.7% formaldehyde 1% agarose gel. After transfer to positively charged nylon, Northern blot analysis was performed using a cDNA probe for CYP3A4. C, total RNA (15 μg/lane) isolated from cultured mouse hepatocytes treated with vehicle (0.1% DMSO), PCN (10 μM), dexamethasone (10 μM), Z-guggulsterone (10 μM), or E-guggulsterone (10 μM) was resolved on a 3.7% formaldehyde 1% agarose gel. After transfer to positively charged nylon, Northern blot analysis was performed using a cDNA probe for mouse Cyp3a11. The Northern blots were stripped and subsequently reprobed with a β-actin cDNA probe. Fold-induction values were calculated by normalizing relative intensities of the CYP3A gene expression levels to β-actin expression as determined using Kodak Image Analysis software.
Q-PCR Analysis of Mouse Cyp3a11. One microgram of DNaseI-treated RNA was reverse transcribed using random primers following the manufacturer's instructions (Promega, Madison, WI). Equal amounts of cDNA were used in real-time quantitative-PCR reactions using the SmartCycler (Cepheid, Sunnyvale, CA). Reactions included 200 nM fluorogenic probe and 150 nM primers specific for β-actin and Cyp3a11. The fluorogenic probe and primer sets were designed using the Primer3 program. BioSearch Technologies (Novato, CA) synthesized the fluorogenic probes. The sequences (5′ to 3′) for the primers and probes are as follows: Cyp3a11, forward primer (CAAGGAGATGTTCCCTGTCA), fluorogenic probe (FAM-AGAAGGCAAAGAAAGGCAAGCCTG-BHQ1), reverse primer (CCACGTTCACTCCAAATGAT); and β-actin, forward primer (CAAGATCATTGCTCCTCCTG), fluorogenic probe (ROX-CAGCAGATGTGGATCAGCAAGCAG-BHQ2), reverse primer (TAACAGTCCGCCTAGAAGCA). Cycling conditions were 95°C for 2 min followed by 45 cycles of 95°C for 15 s, 60°C for 30 s, and 72°C for 30 s using the SmartCycler system. Fold-induction was calculated as described previously (Schmittgen et al., 2000).
GST Pull-Down Assay. GST-PXR.1LBD, GST-p75 (Bilderback et al., 1999), and GST proteins were expressed in BL21DE3 cells, and bacterial extracts were prepared by sonication. Protein lysis buffer contained 10 mM Tris, pH 8.0, 50 mM KCl, 10 mM dithiothreitol, and 1% Nonidet P-40. Lysates were centrifuged at 4000 rpm for 20 min. Glycerol was added to the supernatant to a final concentration of 10%, and lysates were stored at –80°C. Radiolabeled SRC-1 was generated using the TNT rabbit reticulocyte system (Promega). Coprecipitation reactions included 100 μl of lysate containing GST-PXR-LBD, GST-p75 fusion proteins, or control GST; 100 μl of incubation buffer (50 mM KCl, 40 mM HEPES, pH 7.5, 5 mM β-mercaptoethanol, 1% Tween 20, and 1% nonfat dry milk); 5 μl of [35S]SRC-1 and PCN (10 μM); and E-guggulsterone (10 μM), or control DMSO (0.1% DMSO). The mixtures were incubated for 25 min at 4°C with gentle mixing before the addition of 15 μl of glutathione-Sepharose 4B beads (Amersham Biosciences Inc.) that had been extensively washed with phosphate-buffered saline. The mixtures were incubated with gentle mixing at 4°C for an additional 20 min. The beads were pelleted at 2000 rpm in a Microfuge and washed three times with protein lysis buffer containing PCN, E-guggulsterone, or control DMSO. After the last wash, the beads were resuspended in 25 μl of 2× SDS-polyacrylamide gel electrophoresis sample buffer containing 1 mM dithiothreitol. Samples were heated at 100°C for 5 min and loaded onto a 10% SDS-polyacrylamide gel electrophoresis gel. The gel was dried and autoradiography was performed overnight at –80°C. Band intensities were determined using the Kodak 1D Image Analysis software program (Eastman Kodak, Rochester, NY).
Computer Software and Statistical Analysis. The Microsoft Excel spreadsheet program was used to perform statistical analysis (t test). The GraphPad Prism program (GraphPad Software Inc., San Diego, CA) was used to fit the concentration-response relationship curves and determine EC50 values. Quantitation of the northern blotting and GST-pulldown assays was performed using Kodak 1D Image Analysis software.
Results
As part of a systematic study of the potential effects of purified guggulsterones against nuclear receptor family members, a panel of 18 nuclear receptors was screened in agonist mode for their ability to respond to treatment with 10 μM Z-guggulsterone. Guggulsterone treatment activated three nuclear receptor family members above 3-fold, including the ERα isoform, PR, and PXR (Table 1). Eleven-point concentration-response experiments were performed with these three receptors to determine the EC50 values for Z-guggulsterone. Additionally, the percentage of maximal activation at 10 μM concentrations of Z-guggulsterone was determined for purposes of comparison. Guggulsterone treatment activated PR with an EC50 value of approximately 5.9 μM and produced approximately 52% maximal activation at a concentration of 10 μM compared with progesterone at 1 μM (data not shown). Strikingly, Z-guggulsterone activated ERα with an EC50 value of approximately 4.6 μM and produced approximately 137% maximal activation at a concentration of 10 μM compared with β-estradiol at 100 nM (data not shown).
Guggulsterone activates PXR, PR, and ERα
Effects of guggulsterone on nuclear receptor activity. Transactivation by guggulsterone was assessed across a broad panel of nuclear receptors. Transfected cells were treated with 10 μM Z-guggulsterone for 24 h, and reporter activity was measured and normalized to appropriate internal control for each assay. Effect of guggulsterone is expressed as fold-activation relative to a vehicle (dimethyl sulfoxide) control (n = 3). The data represent the mean of assays performed in triplicate ± S.D. and are normalized to β-galactosidase activity.
We examined whether ethanol extracts of gugulipid activate mouse and human PXR using the XREM-LUC reporter gene and CV-1 cells in transient transfection assays. This analysis revealed that gugulipid extract induced XREM-LUC reporter gene activity in a concentration-dependent manner in the presence of mouse or human PXR (Fig. 1A). Additionally, increasing concentrations of Z-guggulsterone partially antagonized full activation of both mouse and human PXR by PCN and rifampicin, respectively (Fig. 1B). Full concentration-response analysis showed that purified Z-guggulsterone activated mouse PXR with a half-maximal effective concentration (EC50) of approximately 1.4 μM (Fig. 2A), whereas human PXR was activated with an EC50 value of approximately 2.4 μM (Fig. 2B). Z-Guggulsterone produced approximately 50% maximal activation at a concentration of 10 μM compared with 10 μM PCN or 10 μM rifampicin against mouse and human PXR, respectively (data not shown).

A, gugulipid extract activates human and mouse PXR. CV-1 cells were transfected with plasmids encoding human PXR or mouse PXR, the β-galactosidase reporter gene, and the XREM-LUC reporter gene. Twenty-four hours post-transfection, concentration-response analysis was performed using 2-fold serial dilutions of gugulipid extract (diluted 1:1000 in ethanol). Cells were treated for 24 h, and luciferase activity was determined. B, CV-1 cells were transfected with plasmids encoding human or mouse PXR, the β-galactosidase reporter gene, and the XREM-LUC reporter gene. Twenty-four hours later, cells were cotreated with 10 μM PXR activators (10 μM rifampicin for human PXR or 10 μM PCN for mouse PXR) and increasing concentrations of Z-guggulsterone (1, 5, 10, and 20 μM) for 24 h. The data represent the mean of assays performed in triplicate ± S.E. and are normalized to β-galactosidase activity and expressed as fold-activation over vehicle control.
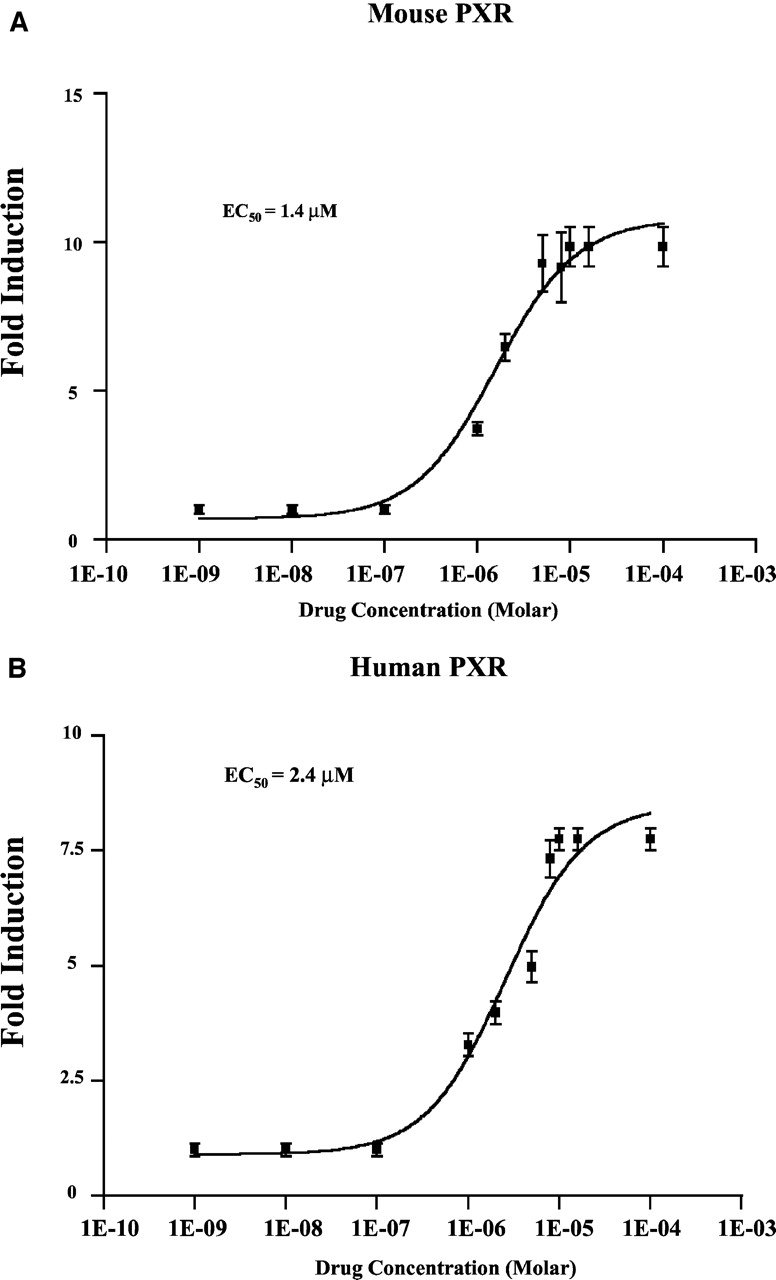
Guggulsterone concentration-response analysis. CV-1 cells were transfected with plasmids encoding either mouse (A) or human PXR (B), together with the β-galactosidase reporter gene, and the XREM-LUC reporter gene. Concentration-response analysis was performed using dilutions of Z-guggulsterone (diluted 1:1000 in DMSO). Cells were subsequently assayed for luciferase activity. The data represent the mean of assays performed in triplicate ± S.E. and are normalized to β-galactosidase activity. EC50 values were obtained by fitting the data to sigmoidal curve. The data are expressed as fold-activation over vehicle control.
To determine whether PXR is required for the induction of the mouse Cyp3a11 gene, real-time Q-PCR was performed using a probe that specifically detects Cyp3a11. Treatment of cultured hepatocytes isolated from wild-type mice with increasing dosages of E-guggulsterone resulted in a dose-dependent increase of Cyp3a11 gene expression, whereas Cyp3a11 gene expression was largely unaffected by any dosage of E-guggulsterone tested using hepatocytes isolated from PXR-KO mice (Fig. 3A). To determine whether guggulsterone treatment of cultured human and rodent hepatocytes can induce the expression of surrogate markers of PXR activation, we performed Northern blotting experiments using radiolabeled cDNA probes that detect the steady-state expression levels of human and rodent CYP3A family members, respectively. This analysis revealed that both E-guggulsterone and gugulipid extract induced the expression of human CYP3A family members (Fig. 3B). Similarly, cultured mouse hepatocytes exhibited marked induction of PXR-target gene expression after treatment with either E- or Z- guggulsterone (Fig. 3C).
To determine whether guggulsterones directly interacts with PXR, we performed pulldown assays with bacterially expressed glutathione S-transferase-PXR-ligand-binding domain fusion protein (GST-PXR). [35S]SRC-1 interacted minimally with the GST-PXR fusion protein in the absence of ligand (Fig. 4A, lane 4), whereas the addition of PCN or purified E-guggulsterone stimulated the interaction between GST-PXR and [35S]SRC-1 (Fig. 4A, lanes 5 and 6). As a control, [35S]SRC-1 protein did not interact with either GST alone or GST-p75 neurotrophin (Fig. 4A, lanes 2 and 3). Quantitative analysis of three independent experiments showed that E-guggulsterone stimulated interaction with [35S]SRC-1 (Fig. 4B). Very similar results were obtained using Z-guggulsterone with [35S]SRC-2 and [35S]SRC-3 protein interaction experiments (data not shown).

Guggulsterone modulates PXR interaction with nuclear receptor cofactors. A, GST-pulldown assays were performed with [35S]SRC-1 synthesized in vitro (lane 1), bacterially expressed GST alone (lane 2), GST-p75 (lane 3), or GST-PXR LBD (lanes 4–6). [35S]SRC-1 was mixed with GST, GST-p75, or GST-PXR LBD in the presence of vehicle (0.1% DMSO), PCN (10 μM), or E-guggulsterone (10 μM). B, quantitative analysis of three independent experiments was performed using Kodak Image Analysis software. The data are plotted as relative intensities of the [35S]SRC-1 band compared with the vehicle control and are reported as the mean ± S.E. of three independent experiments.
To create a high-throughput assay that can be used to screen a large number of herbal mixtures or discrete chemical compounds for PXR activation, we fused XREM to the SPAP reporter gene. We chose the SPAP reporter gene because it encodes a gene product that can be assayed using the media of cultured cells because it is the heat-stable and secreted form of alkaline phosphatase. The use of 96-well plates for cell culture and reporter gene assays ensures that this system has the potential to rapidly test large numbers of herbal mixtures or discrete chemical compounds for their ability to induce hepatic CYP3A4 expression at the transcriptional level through activation of PXR, while minimizing both the expense and the amount of reagents required.
Cotransfection of PXR and HNF4α together with the XREM-SPAP reporter gene into CV-1 cells elicits maximal reporter gene expression and enables accurate dose-response analysis of herbal mixtures and discrete compounds (Tirona et al., 2003). To validate this reporter gene system, we determined the EC50 value of rifampicin against human PXR to be approximately 800 nM, a value in close agreement with previously published results obtained using the CAT reporter gene (Lehmann et al., 1998). Full dose-response analysis of Z-guggulsterone using the XREM-SPAP reporter gene indicated activation of human PXR with an EC50 value of 1.2 μM, a value in close agreement with that which we obtained using the XREM-LUC reporter gene assay (Fig. 5).

XREM-SPAP as a tool for screening additional herbal remedies for PXR activation. A concentration-response analysis was performed using the XREM-SPAP reporter gene in the presence of mouse and human PXR. Twenty-four hours after transfection, CV-1 cells were treated with increasing concentrations of E-guggulsterone for an additional 24 h. The cell culture media was then assayed for SPAP activity as described under Materials and Methods. The data represent the mean of assays performed in triplicate ± S.E. and are normalized to β-galactosidase activity. EC50 values were obtained by fitting the data to sigmoidal curve and are reported as the mean ± S.E. of three independent experiments.
Discussion
Guggulsterones are plant sterols derived from the gum resin of the tree C. mukul. Guggulsterones are contained in over-the-counter herbal remedies as a complex mixture of compounds termed gugulipid and are used for their serum lipid-lowering properties (Nityanand et al., 1989). Guggulsterones have been shown to be the active ingredient in gugulipid (Beg et al., 1996). Guggulsterones also exhibit thyroid-stimulating activity (Tripathi et al., 1984) and inhibit platelet aggregation (Mester et al., 1979). Recent studies indicate that guggulsterones may lower cholesterol through antagonism of FXR, a member of the nuclear receptor superfamily of ligand-activated transcription factors.
A systematic screen with a panel of yeast protein GAL4 nuclear receptor ligand-binding domain fusions was used to identify purified Z-guggulsterone as an FXR-antagonist (Urizar et al., 2002; Wu et al., 2002). Our discovery that guggulsterones can function as agonists for at least three nuclear receptors raises the intriguing possibility that, in addition to antagonism of FXR, the molecular basis of the lipid-lowering properties of gugulipid therapy may be multifactorial. Previous studies of guggulsterone-mediated activation of PXR used the GAL4-ligand-binding domain format (Urizar et al., 2002; Wu et al., 2002). Our findings extend those results and show that purified guggulsterones activate full-length PXR, ERα, and PR nuclear receptor superfamily members using target-gene specific reporter gene assays in transfected cells. Guggulsterone activates these receptors with EC50 values in the low micromolar range, concentrations achieved in serum during gugulipid therapy (Arya, 1988). Moreover, we show that both gugulipid and purified guggulsterones induce expression of the CYP3A family members in both rodent and human cultured hepatocytes through PXR activation. This is an important observation in the light of the recent finding that the active ingredient in St. John's wort, hyperforin, is a potent PXR agonist that induces herb-drug interaction through induction of CYP3A4 enzyme activity in patients on combination therapy (Moore et al., 2000; Wentworth et al., 2000).
Gugulipid is an herbal remedy that is available over-the-counter and is not subject to the rigorous testing that formal drug candidates undergo. These studies further extend the range of human and rodent PXR ligands to include purified guggulsterones. Together, our data suggest the CYP3A drug-metabolizing pathway may be induced in patients during gugulipid therapy, which is likely to alter the disposition of any coadministered medications that are metabolized by CYP3A enzymes. Our findings further support the notion that herbal remedies should be tested for their ability to activate PXR to identify potential herb-drug interactions. The medium-throughput 96-well plate assay that we present here is likely to be helpful and may serve as an important predictor of herb-drug interactions. Moreover, our data suggest that studies of guggulsterone's agonist activity against the ERα and PR nuclear receptor family members are warranted. It is interesting to note that whereas E-guggulsterone activates ERα, it does not activate the ERβ isoform. Finally, our data suggest that it may be possible to identify and develop safer guggulsterone analogs that retain their lipid-lowering activity but that do not activate PXR and promote drug metabolism.
Footnotes
-
This work was supported by National Institutes of Health COBRE Grant RR17708-01.
-
DOI: 10.1124/jpet.103.064329.
-
ABBREVIATIONS: FXR, farnesoid X receptor; PXR, pregnane X receptor; HNF, hepatocyte nuclear factor; XREM, xenobiotic response element; ER, estrogen receptor; PR, progesterone receptor; SPAP, secreted placental alkaline phosphatase; LBD, ligand-binding domain; PCR, polymerase chain reaction; PPAR, peroxisome proliferator-activated receptor; KO, knockout; Q-PCR, quantitative-polymerase chain reaction; DMEM, Dulbecco's modified Eagle's medium; DMSO, dimethyl sulfoxide; GST, glutathione S-transferase; RXR, retinoic X receptor; SRC, steroid receptor coactivator; TR, thyroid receptor; PCN, 5-pregnen-3β-ol-20-one-16α-carbonitrile.
- Received December 12, 2003.
- Accepted April 1, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}