


Abstract
Oxazepam is a commonly used 1,4-benzodiazepine anxiolytic drug that is polymorphically metabolized in humans. However, the molecular basis for this phenomenon is currently unknown. We have previously shown that S-oxazepam glucuronide, the major oxazepam metabolite, is selectively formed by UDP-glucuronosyltransferase (UGT) 2B15, whereas the minor Roxazepam glucuronide is produced by multiple UGTs other than UGT2B15. Phenotype-genotype studies were conducted using microsomes and DNA prepared from the same set of 54 human livers. Sequencing of the UGT2B15 gene revealed three nonsynonymous polymorphisms, D85Y, T352I, and K523T, with variant allele frequencies of 0.56, 0.02, and 0.40, respectively. D85Y genotype showed a significant effect (p = 0.012) on S-oxazepam glucuronidation with lower median activities in 85Y/Y livers (49 pmol/min/mg protein) compared with 85D/D livers (131 pmol/min/mg), whereas 85D/Y livers were intermediate in activity (65 pmol/min/mg). There was also a significant trend (p = 0.049) for higher S-oxazepam activities in the two 352T/I livers (135 and 210 pmol/min/mg) compared with the remaining 352T/T livers (median, 64 pmol/min/mg). Conversely, K523T genotype had no apparent effect on oxazepam glucuronidation (p > 0.05). Donor gender also significantly influenced S-oxazepam glucuronidation with higher median activities in male (65 pmol/min/mg) compared with female (39 pmol/min/ mg) livers (p = 0.042). R-Oxazepam glucuronidation was not affected by either genotype or gender (p > 0.05). In conclusion, gender and D85Y genotype are identified as major determinants of S-oxazepam glucuronidation by human liver and may explain in part polymorphic oxazepam glucuronidation by human subjects.
Glucuronidation, catalyzed by the UDP-glucuronosyltransferases (UGTs), is one of the major metabolic pathways responsible for elimination of potentially harmful xenobiotic and endobiotic compounds in mammals. UGT2B15 is one of more than 16 human UGT isoforms that have been discovered to date. Although originally identified as an enzyme with high intrinsic activity for glucuronidation of androgenic steroids (Chen et al., 1993), subsequent studies with recombinant UGT2B15 indicate that this enzyme is also likely to have a significant role in the metabolism of drugs and other xenobiotics (Green et al., 1994). Pharmaceutical substrates of UGT2B15 that have been identified include oxazepam, E-4-hydroxytamoxifen, 5-hydroxyrofecoxib, eugenol, 8-hydroxyquinoline, phenolphthalein, 4′-hydroxyphenytoin, and nandrolone (Green et al., 1994; Court et al., 2002; Nishiyama et al., 2002; Kuuranne et al., 2003; Zhang et al., 2003). Oxazepam, a commonly used 1,4-benzodiazepine anxiolytic drug, is of considerable interest because it is cleared primarily by glucuronidation and has been used extensively as an in vivo probe of drug glucuronidation in people (Greenblatt et al., 1980, 1984; Greenblatt, 1981; Sonne, 1993). Furthermore, the glucuronidation of this drug by human subjects is known to be polymorphic (Patel et al., 1995a), although the molecular mechanisms responsible for this variability are not well understood.
A previous study of oxazepam biotransformation in human subjects by Patel et al. (1995a) identified S-oxazepam glucuronide as the major oxazepam metabolite in human plasma and urine (Fig. 1). Smaller amounts of R-oxazepam glucuronide were also found in both plasma and urine with S/R-oxazepam glucuronide ratios averaging 3.5 and 3.9, respectively. Intriguingly, in 2 of 11 (18%) subjects evaluated, S/R ratios in the urine and plasma clearance of oxazepam were both very low compared with values in the remaining nine subjects (over 3-fold difference). This finding was considered by the investigators to be consistent with the presence of a slow metabolizer phenotype in the population. Further in vitro studies (Patel et al., 1995a) with human liver microsomes (HLMs) verified S-oxazepam glucuronide as the major metabolite (S/R ratios averaging 4.0) and also identified 4 of 37 (11%) livers with relatively low oxazepam glucuronidation activities (S/R metabolite ratios <2.0). Enzyme kinetic analysis of S-oxazepam glucuronidation suggested that these low activities were associated with higher Km values and lower Vmax values for the four atypical livers compared with the remaining livers. Consequently, slow S-oxazepam glucuronidation was hypothesized to result from relatively lower amounts of a high-affinity S-oxazepam glucuronidating enzyme in those livers, perhaps as a consequence of polymorphism in the gene encoding this particular (as yet unidentified) enzyme.

Pathways responsible for the metabolism of oxazepam by glucuronidation in humans.
Recent work in our laboratory (Court et al., 2002) indicates that UGT2B15 is the principal enzyme responsible for stereoselective glucuronidation of oxazepam in human liver (Fig. 1). Specifically, of 12 different recombinant UGTs evaluated, UGT2B15 was the only isoform to show significant S-oxazepam glucuronide formation, whereas R-oxazepam glucuronide was formed by multiple UGTs, including UGT1A9 and UGT2B7. Furthermore, Km values for S-oxazepam glucuronidation by recombinant UGT2B15 (29–35 μM) were similar to values determined for HLMs (43–60 μM). Finally, we showed that S-oxazepam glucuronidation activities were 5-fold lower in microsomes from human embryonic kidney 293 cells overexpressing a common allozyme variant of UGT2B15 (tyrosine at codon 85; Y85) compared with the wild-type enzyme (aspartate at codon 85; D85).
The relatively high prevalence of the UGT2B15 Y85 gene variant in the population (45–63%) (Lampe et al., 2000), coupled with the observed influence of this amino acid substitution on oxazepam glucuronidation by recombinant enzyme, suggests that this genetic polymorphism may be the molecular basis for polymorphic oxazepam metabolism in people. Consequently the objective of the present study was to explore this hypothesis through genotype-phenotype study of a well characterized human liver bank (n = 54 individuals). In addition to genotyping all of these livers for the D85Y polymorphism, novel polymorphisms were identified by sequencing each of the exons including exon-intron junctions of the UGT2B15 gene. The results indicate that both D85Y genotype and gender are major determinants of oxazepam glucuronidation by human liver and may explain in part polymorphic oxazepam glucuronidation by human subjects.
Materials and Methods
Reagents. Chemicals were purchased from Sigma-Aldrich (St. Louis, MO) except for acetonitrile, which was from Fisher Scientific Co. (Fairlawn, NJ). Oxazepam and oxazepam glucuronides were gifts from Wyeth Pharmaceuticals (Philadelphia, PA). The oxazepam glucuronide standard preparation was a mixture of the two enantiomers that contained 86% S-oxazepam glucuronide and 14% R-oxazepam glucuronide as determined by high-performance liquid chromatography (HPLC) analysis.
Human Liver Microsomes. Liver samples from donors with no known liver disease were obtained from either the National Disease Research Interchange (Philadelphia, PA) or the Liver Tissue Procurement and Distribution Service (Minneapolis, MN) with the approval of the Tufts University Institutional Review Board. All livers were either intended for transplantation but had failed to tissue match, or were normal tissue adjacent to surgical biopsies. Donors were primarily of Caucasian ethnicity (n = 48), but also included four African-Americans and two Hispanics. Other available demographic information included gender (16 females, 38 males) and age (median 41 years; range 2–75 years). Smoking history was positive for 19 donors, negative for 29 donors, and unknown for six donors, whereas significant alcohol use (defined as 14 or more drinks per week) was positive for 13 donors, negative for 34 donors, and unknown for eight donors.
HLMs were prepared by differential ultracentrifugation as described previously (Court et al., 1997). Microsomal pellets were suspended in 0.1 M potassium phosphate buffer (pH 7.5) containing 20% glycerol and kept at–80°C until use. The protein concentration of HLM samples was determined using the bicinchoninic acid protein assay (Pierce Chemical, Rockford, IL) with bovine serum albumin as the standard. The quality of the liver samples was ascertained by reference to at least eight other cytochrome P450 activities measured in this laboratory using the same set of livers. Livers that consistently showed low activity values (>2-fold lower for all measured activities) relative to the median activity value for the entire liver set had been excluded from the study.
Oxazepam Glucuronidation Assay.S- and R-Oxazepam glucuronidation activities were measured in duplicate using the entire set of HLMs as described in detail previously with minor differences (Court et al., 2002). Briefly, 100-μl incubations were conducted at 37°C for 180 min in 50 mM phosphate buffer (pH 7.5) with 5 mM MgCl2, 5 mM UDP-glucuronic acid, 100 μM oxazepam, 2.5 μg of alamethicin, and 50 μg of microsomal protein. Incubations were terminated by addition of 50 μl of acetonitrile with 1 μg of phenacetin (internal standard), vortexed, and placed on ice. Samples were centrifuged, and the supernatants were analyzed by HPLC with UV absorbance detector set at a wavelength of 214 nm (model 1100; Agilent, Palo Alto, CA). S- and R-Oxazepam glucuronidation activities were calculated by dividing metabolite concentration by protein concentration and incubation time and expressed as picomoles per minute per milligram of protein. Intra- and interassay variabilities were less than 10 and 15%, respectively. S/R-Oxazepam glucuronidation ratios were calculated by dividing the observed S- by the R-oxazepam glucuronidation activity determined for each liver.
Sequencing of the UGT2B15 Gene. The UGT2B15 gene sequence used in this study was identified in the GenBank reference sequence NT_077444.2. UGT2B15 gene positions are relative to the first nucleotide of the predicted translation start codon (nucleotide 195,500 of NT_077444.2 with the gene on the reverse complement strand). Genomic DNA was isolated with DNAzol (Invitrogen, Carlsbad, CA) from the same human liver tissues used to prepare microsomes. Five sets of PCR amplification primers (Table 1) were designed to amplify the six UGT2B15 exons including exon-intron boundaries. One set of primers (Pri 229 and Pri 230) amplified both exons 4 and 5. Primers were selected from regions of the UGT2B15 gene that differed from the closely homologous UGT2B17 gene sequence (found in GenBank reference sequence NT_022778.14). This was confirmed by showing complete match of the PCR-derived sequences with the known reference UGT2B15 sequence and mismatch with the corresponding UGT2B17 sequence. Typical 50-μl PCR reactions contained 100 ng of genomic DNA, 2 mM MgCl2, 0.2 mM dNTPs, 5 μl of 10× PCR buffer, 2.5 U of TaqDNA polymerase (Platinum Taq; Invitrogen), and 0.2 μM forward and 0.2 μM reverse primers. Samples were denatured for 10 min at 95°C and then subjected to 40 cycles of 95°C for 30 s, the appropriate anneal temperature (below) for 30 s, and 72°C for 1 min, followed by a final extension cycle of 72°C for 15 min. Anneal temperatures were 64°C for primers 118 and 119, 65°C for primers 225 and 226, 53°C for primers 227 and 228, 56°C for primers 229 and 230, and 63°C for primers 202 and 203. PCR products were confirmed by agarose gel electrophoresis, purified using a QIAquick PCR purification kit (QIAGEN, Valencia, CA) and sequenced (ABI PRISM 3100; Applied Biosystems, Foster City, CA) using the PCR primers 118, 225, 229, and 202, and an additional internal sequencing primer 237 (5′-TGA CCA TAA AGA ATG TGA CTG TAT G-3′).
Oligonucleotide primers used for genomic PCR in this study
Data Analysis. ARLEQUIN version 2.001 (Schneider et al., 2001) was used to evaluate SNP genotype frequencies for consistency with the Hardy-Weinberg equilibrium by a modified Markov-chain random walk algorithm. The significance of linkage disequilibrium between identified SNPs was also assessed by ARLEQUIN using a pairwise likelihood-ratio test. SNPs identified in the coding regions of each exon were evaluated for effects on the predicted amino acid sequence by use of the virtual translation capability of the program Vector NTI Suite 7.0 (Invitrogen).
Statistical analyses were conducted with the Sigmastat program (version 3.0; SPSS Inc., Chicago, IL). Data were routinely evaluated for normality of distribution and equal variance, and in instances where these tests failed, nonparametric methods of statistical analysis were used. The effect of donor gender and histories of smoking and alcohol use on each of the phenotype parameters (S-oxazepam glucuronidation, R-oxazepam glucuronidation, and S/R-oxazepam glucuronidation ratio) was evaluated by the Mann-Whitney rank sum test, whereas the effects of genotype were analyzed by one-way Kruskal-Wallis ANOVA on rank transformed data. In instances where the ANOVA indicated a significant effect, Dunn's multiple comparison method was used to identify individual groups that were significantly different. Because donor gender seemed to significantly affect some of the phenotypic parameters, data were also stratified by gender and reanalyzed for genotype effects. Separate analyses were also conducted in data sets restricted to those from Caucasian individuals to minimize genetic variability (from sources other than those being measured). Multisite haplotypes were reconstructed from the coding SNP data using the program PHASE version 2.0.1 (Stephens et al., 2001). The effect of diplotype (haplotype pair) on phenotype parameters were evaluated by one-way Kruskal-Wallis ANOVA on ranks. Multiple linear regression analysis was also used to estimate the relative contribution of donor gender and UGT2B15 genotype to the total observed variability in oxazepam glucuronidation measurements.
Results
UGT2B15 Gene Exon and Exon-Intron Junction SNPs. Sequencing of the UGT2B15 gene exons and exonintron junctions in genomic DNA from the livers of 54 individuals revealed 12 different SNPs (Fig. 2). Three of these were nonsynonymous (cSNPs) resulting in the amino acid variants D85Y (253g>t), T352I (15485c>t), and K523T (23476c>a) located in exons 1, 4, and 6, respectively. Both the D85Y and K523T variants were highly prevalent with variant allele frequencies of 0.56 and 0.40, respectively, whereas the T352I variant was only found in two individuals, and both were heterozygotes. One SNP was identified in exon 5 (16420g>a) at a frequency of 0.04 but was not predicted to affect the amino acid sequence (silent). The remaining SNPs were located in the introns (n = 5) and 3′ untranslated region (n = 2). Of the 12 SNPs identified in this study, a number of these (D85Y, 2635a>t, 7746g>a, K523T, 23476c>a, 23669c>t, and 23687a>t) have been reported previously by Iida et al. (2002), whereas the remaining six SNPs (including the T352I cSNP) to our knowledge were considered to be novel unpublished findings.

SNPs identified by sequencing all six exons including exon-intron boundaries of the UGT2B15 gene using DNA obtained from 54 human livers.
The frequency distributions of genotypes for all of the identified SNPs were consistent with Hardy-Weinberg equilibrium (p > 0.05), indicating that the liver bank was likely a representative sample of the population being studied. Highly significant linkage disequilibrium (p < 0.001) was identified between 253g>t (D85Y) and 2635a>t as well as between D85Y and 7743a>c (Fig. 3). Similarly, there was significant linkage between 23476c>a (K523T) and 23669c>t. These associations were similar for both the entire mixed population (Fig. 3A) as well as the Caucasian subpopulation (Fig. 3B).

Genetic linkage analysis of UGT2B15 SNPs identified in a bank of 54 human livers. The significance of linkage disequilibrium between SNP pairs was assessed using a pairwise likelihood-ratio test as implemented in the ARLEQUIN program version 2.001 (Schneider et al., 2001). Analyses were conducted using either the entire mixed race population (A) or the Caucasian subpopulation (B).
Oxazepam Glucuronidation by the Human Liver Bank. The frequency distributions of S- and R-oxazepam glucuronidation activities and S/R-oxazepam glucuronidation ratios measured in the set of HLMs (n = 54) are shown in Fig. 4. S-Oxazepam glucuronidation activities (median, 65 pmol/min/mg protein; range, 10–212 pmol/min/mg protein) and S/R-oxazepam glucuronidation ratios (3.2; 0.6–10) were not normally distributed (p < 0.05; Kolmogorov-Smirnov normality test) with significant skewing toward lower values. Conversely, R-oxazepam glucuronidation activities (median, 21 pmol/min/mg protein; range, 5–49 pmol/min/mg protein) were normally distributed (p > 0.05). Interindividual variability as indicated by the ratio of the maximum value divided by the minimum value was greatest for S-oxazepam glucuronidation (21-fold), intermediate for S/R-oxazepam glucuronidation ratio (16-fold), and least for R-oxazepam glucuronidation (10-fold).

Distribution S-oxazepam glucuronidation activities (A), R-oxazepam glucuronidation activities (B), and S/R-glucuronidation ratios (C) measured using the same set of HLMs.
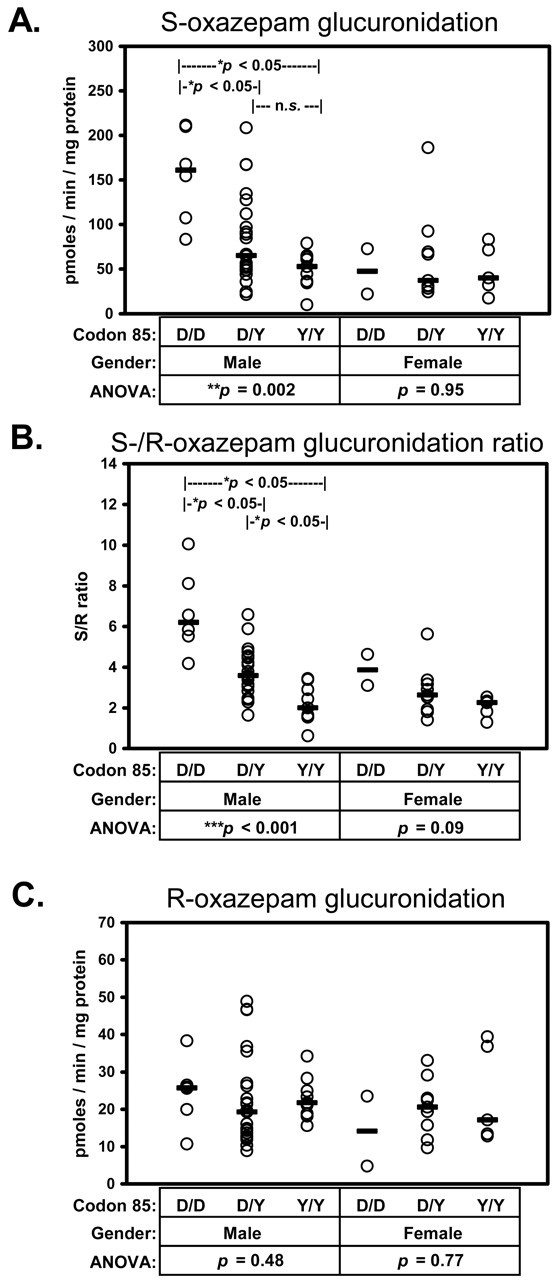
Effect of Liver Donor Demographics and UGT2B15 SNP Genotype on Oxazepam Glucuronidation. As shown in Table 2, univariate statistical analysis identified donor gender (p = 0.042) and the SNP genotypes D85Y (p = 0.012) and T352I (p = 0.046) as having significant effects on S-oxazepam glucuronidation activities, whereas donor gender (p = 0.02) and the SNP genotypes D85Y (p < 0.001) and 7743 a>c (p = 0.005) affected S/R-oxazepam glucuronidation ratios. The effect of gender on S/R-glucuronidation ratios and S-oxazepam glucuronidation was similar, resulting in 40 and 67% higher median activities, respectively, in male livers compared with female livers. The D85Y SNP genotype also affected both S/R-glucuronidation ratios and S-oxazepam glucuronidation activities, resulting in 40 and 50% lower median values, respectively, in heterozygous (85D/Y) livers and 63% lower median values (for both parameters) in homozygous variant (85Y/Y) livers compared with homozygous reference (85D/D) livers. None of the examined variables were found to influence R-oxazepam glucuronidation activities (p > 0.05). Furthermore, none of the remaining SNPs were found to influence oxazepam glucuronidation.
Univariate analysis of effects of liver donor gender and UGT2B15 genotype on oxazepam glucuronidation
Similar analysis of other available demographic data (smoking and alcohol consumption histories) and other genotypes showed no significant effect (p > 0.05).
These genotype-phenotype analyses were then repeated with data stratified to control for gender effect. As shown in Fig. 5, in male subjects D85Y genotype had significant effects on both S-oxazepam glucuronidation activities (p = 0.002) and S/R-glucuronidation ratios (p < 0.001) but not on R-oxazepam glucuronidation activities (p = 0.48). Median S-oxazepam glucuronidation activities in male 85D/Y and 85Y/Y livers were significantly lower (by 60 and 67%) than that of male 85D/D livers. Similarly, S/R-oxazepam glucuronidation ratios in male 85D/Y and 85Y/Y livers were significantly lower (by 42 and 68%, respectively) compared with male 85D/D livers. On the other hand, significant D85Y genotype differences in oxazepam glucuronidation were not observed in female livers (Fig. 5). Analysis of the effect of other genotypes in male livers showed significantly higher S/R-oxazepam glucuronidation ratios (p = 0.025) in K523T heterozygotes (K/T) compared with wild-type homozygotes (T/T) with median (25–75%) values of 4.4 (3.1–5.9; n = 18) and 3.1 (2.0–3.6; n = 16), respectively. Compared with 523T/T livers, S/R-oxazepam glucuronidation ratios were also higher in homozygous 523K/K livers with a median value of 3.6 (2.4–4.2; n = 3), although the difference was not statistically significant probably because of the small number of individuals with this genotype.

Influence of liver donor gender and UGT2B15 codon 85 genotype on rates of S-oxazepam glucuronidation (A) and R-oxazepam glucuronidation (B) and on S/R-glucuronidation ratios (C) measured in a set of 54 HLMs. Also shown are the p values for Kruskal-Wallis one-way ANOVA on rank transformed data. In instances where the ANOVA indicated a significant effect, differences between individual genotype groups were evaluated using Dunn's multiple comparison method (*, p < 0.05).
Significant effects of genotype were not observed in female livers, although there was a trend (p = 0.09) for lower median S/R-oxazepam glucuronidation ratios in female 85Y/Y livers (2.3; 1.7–2.4; n = 5) compared with 85D/Y (2.6; 1.9–3.2; n = 9) and 85D/D (3.9; 3.1–4.6; n = 2) livers. There were no effects of any other genotype on oxazepam glucuronidation in livers of either gender. Essentially identical results were obtained when the subgroup of Caucasian individuals was used for these analyses, except that the previously observed trend for lower S/R-oxazepam glucuronidation ratios in 85Y/Y variant female livers was more apparent, closely approaching statistical significance (p = 0.054).
UGT2B15 Gene Haplotype Structure. Five unique haplotypes could be inferred using genotype data for the three nonsynonymous SNPs (Table 3). The most common haplotype was UGT2B15*2 (D85Y), representing 27% of alleles. Less common haplotypes included UGT2B15*1 (the reference sequence; 17% alleles), UGT2B15*5 (D85Y, K523T; 14% alleles), UGT2B15*4 (K523T; 11% alleles), and UGT2B15*6 (T352I; 2% alleles). The UGT2B15*3 allele (L86S), which was identified previously (http://som.flinders.edu.au/FUSA/ClinPharm/UGT/2B15alleles.html), was not found in any of the individuals in this study population.
Haplotypes inferred from coding SNPs identified by sequencing the UGT2B15 gene exons using genomic DNA from 54 human livers
Haplotypes were inferred using the program PHASE version 2.00.1 (Stephens et al., 2001; Stephens and Donnelly, 2003). n = number of chromosomes evaluated.
For the 54 livers analyzed, nine different haplotype pairs (diplotypes) could be identified. The most common diplotype was*1/*2 (n = 10). Other observed diplotypes included *2/*2 (n = 7), *1/*4 (n = 6), *4/*5 (n = 5), *2/*5 (n = 4), *5/*5 (n = 3) *4/*6 (n = 1), *2/*6 (n = 1), and *1/*1 (n = 1). Although diplotypes for 16 of 54 livers (30%) could not be definitively resolved, all were either *1/*5 or *2/*4. Consequently, in subsequent analyses these livers were considered as a single diplotype group (i.e., “*1/*5 or *2/*4”).
Effect of UGT2B15 Haplotype on Oxazepam Glucuronidation. The effects of UGT2B15 diplotype on oxazepam glucuronidation were then evaluated by Kruskal-Wallis ANOVA on ranks using data from all livers and from male livers only (Fig. 4). The smaller number of female livers precluded meaningful analysis of this group. Significant effects of diplotype on S/R-oxazepam glucuronidation ratios and S-oxazepam glucuronidation were found for the entire set of data (p < 0.001 and p < 0.001, respectively), as well as for male livers only (p = 0.002 and p = 0.005, respectively). Post hoc multiple pairwise comparisons of the entire liver data set showed over 2-fold higher S/R-oxazepam glucuronidation ratios (p < 0.05) in *1/*4 livers compared with *2/*2, *2/*5, and *5/*5 livers with median ratios of 5.6, 2.3, 2.1, and 2.0, respectively (Fig. 4A). Similarly, analysis of data from the male liver only group showed over 2-fold higher ratios (p < 0.05) in *1/*4 livers compared with *2/*2 and *2/*5 livers with median ratios of 5.8, 2.2 and 2.1 (respectively), whereas there was only one male *5/*5 liver (ratio of 2.0). For S-oxazepam glucuronidation, using data from all livers significant differences (p < 0.05) were observed between the *5/*5 (median, 32 pmol/min/mg protein) compared with the *4/*5 (median, 167 pmol/min/mg protein) and the *1/*4 (median, 131 pmol/min/mg protein) livers. No diplotype group differences within the male only group were observed for S-oxazepam activities. Further stratification of diplotype data into Caucasian only and male Caucasian groups yielded results that were essentially identical to results obtained using the entire data set and the male only group, respectively.
Contribution of Donor Gender and D85Y Genotype to Oxazepam Glucuronidation Variability. Multiple linear regression analyses were then performed using donor gender and D85Y genotype as independent variables with either S-oxazepam glucuronidation activity or S/R-oxazepam glucuronidation ratio as the dependent variable. Estimates of the relative contribution of donor gender and D85Y genotype to total variability were 9 and 17%, respectively, for S-oxazepam glucuronidation activities (r2 = 0.26; p < 0.001) and were 14 and 35%, respectively, for S/R-oxazepam glucuronidation ratios (r2 = 0.49; p < 0.001).
Discussion
Previous studies in our laboratory have determined that S-oxazepam is a selective substrate for UGT2B15 and that a common UGT2B15 allozyme variant (D85Y) has altered S-oxazepam glucuronidation activity (Court et al., 2002). In the present investigation, we demonstrate for the first time that the D85Y cSNP, as well as donor gender, are major determinants of interindividual variability of oxazepam glucuronidation by human liver. Specifically, multiple linear regression analysis indicated that as much as 35% of total variability in oxazepam glucuronidation by HLMs could be accounted for by D85Y genotype, whereas as much as 14% of total variability was associated with donor gender.
To our knowledge, this is also the first instance in which altered drug glucuronidation capability in human-derived tissues has been attributed to the presence of a naturally occurring UGT variant. Common amino acid coding variants have also been identified in the UGT2B4 (Levesque et al., 1999), UGT2B7 (Jin et al., 1993; Coffman et al., 1998), UGT1A6 (Ciotti et al., 1997), and UGT1A7 (Guillemette et al., 2000) genes with variable effects on drug glucuronidation capacity when evaluated by assay of recombinant enzymes. However, these predictions have yet to translate into a demonstrated functional impact on drug glucuronidation in human tissues or on drug clearance in human subjects. For example, the H286Y cSNP in the UGT2B7 gene was proposed to result in decreased zidovudine glucuronidation based on showing similar Km values but over 5-fold lower Vmax values for the Y286 variant compared with wild-type H286 expressed in mammalian cells (Barbier et al., 2000). However, recent studies in this laboratory (Court et al., 2003) and by other investigators (Bhasker et al., 2000) using genotyped HLMs failed to show any effect of the H268Y polymorphism on the glucuronidation of a range of UGT2B7 substrates, including zidovudine, morphine, and codeine. Our study (Court et al., 2003) also confirmed that both zidovudine and morphine are relatively selective substrates of UGT2B7 in human liver and so any functional impact of the H268Y polymorphism should have been observed. Furthermore, a recent in vivo study by Holthe et al. (2002) also failed to show any association of this polymorphism with variation in the ratios of morphine glucuronide to morphine in the serum of cancer patients receiving this drug for analgesia. Consequently, although we had previously established that the D85Y polymorphism significantly impacted S-oxazepam glucuronidation by recombinant enzymes, it was important to verify this effect in the current study using HLMs. However, to irrefutably establish that the D85Y cSNP is the molecular basis for polymorphic oxazepam glucuronidation in people, it will be necessary to further corroborate these findings through phenotype-genotype studies of human subjects.
Like other 3-hydroxybenzodiazepines (including temazepam and lorazepam), oxazepam is an optically active compound with a chiral center at the 3-carbon position (Fig. 1). Although it is possible to obtain optically pure preparations of these compounds, racemization is extremely rapid under aqueous conditions with a half-life of less than 5 min (Yang and Lu, 1989). However, glucuronide conjugation normally occurs via the 3-carbon hydroxyl group, yielding stable diastereomeric glucuronides (R- and S-oxazepam glucuronide) that are readily separated and quantitated via HPLC (Court et al., 2002).
Apart from S-oxazepam glucuronidation, we evaluated the ratio of Sto R-oxazepam glucuronidation activities as a phenotypic measure of UGT2B15 function. R-Oxazepam seems to be glucuronidated by multiple UGT isoforms (Court et al., 2002). Consequently, by normalizing S-oxazepam glucuronidation activities to R-oxazepam glucuronidation activities we had hoped to control for effects on S-oxazepam glucuronidation that were independent of UGT2B15 function such as variability related to the experimental procedure and the condition of HLMs. Indeed, variability in S/R-oxazepam glucuronidation ratios (16-fold) was 25% less than that of S-oxazepam glucuronidation activities (21-fold). Furthermore, the effects of gender and D85Y genotype on S/R-oxazepam glucuronidation ratios consistently showed higher p values than effects on S-oxazepam glucuronidation activities. Ratios of S-oxazepam glucuronide to R-oxazepam glucuronide concentrations in urine and blood have also been used to quantify oxazepam metabolism in human subjects (Patel et al., 1995a). In fact, the S/R-oxazepam glucuronide concentration ratio measured in urine collected over the first 8 h after oxazepam administration is an excellent predictor (rs = 0.90) of the plasma clearance of oxazepam (Patel et al., 1995a).
Patel et al. (1995b) previously suggested that UGT2B7 was responsible for S-oxazepam glucuronidation based on inhibition of liver microsomal activity by known UGT2B7 substrates. However, the specificity of these inhibitors was not validated, and so it is possible that S-oxazepam glucuronidation by UGT2B15 rather than by UGT2B7 was being inhibited. In support of this contention, our previous work (Court et al., 2002) showed over 10-fold higher S-oxazepam glucuronidation by recombinant UGT2B15 compared with UGT2B7. The dominant contribution of UGT2B15 to hepatic S-oxazepam glucuronidation is further validated by the results of the present study, which showed that a common coding UGT2B15 gene sequence variation (D85Y) substantially impacted human liver S-oxazepam glucuronidation by human liver.
Two other nonsynonymous SNPs were also identified in the UGT2B15 gene coding region, including the relatively common K523T and the rarer T352I. The K523T cSNP showed a relatively weak effect with approximately 30% higher S/R-oxazepam glucuronidation ratios in variant carrier livers compared with wild-type livers. However, this difference was observed only in male livers but not in female livers. Because the presence or absence of the D85Y cSNP could have influenced these findings, haplotype analysis was used to further isolate any possible effect of K523T. As shown in Fig. 6, in the subgroup of livers that were identified as being homozygous Y/Y at codon 85 (*2/*2, *2/*5, and *5/*5) there was no apparent modifying effect of the 523T variant (*5 livers) compared with the K523 allele (*2 livers) (p > 0.05). Consequently, it is likely that any possible effect of the K523T cSNP on oxazepam glucuronidation is relatively modest compared with the D85Y cSNP. The T352I cSNP also seemed to be associated with enhanced S-oxazepam glucuronidation activity, although the low genotype frequency (only two heterozygous carriers) precludes any meaningful conclusions. Heterologous expression and functional analysis of both the K523T and T352I variants would help to verify the effect of these mutations on glucuronidation by UGT2B15. Initial phenotype-genotype analyses also showed a strong association of the intron 3 SNP 7743a>c with higher S/R-oxazepam ratios. However, further analysis revealed a highly significant correlation (p < 0.001) of this SNP with the D85Y cSNP indicating that the effect was probably indirect through genetic linkage rather than a direct effect of the mutation.

Influence of UGT2B15 diplotype on S/R-glucuronidation ratios measured using the entire set of 54 HLMs (A) or the subset of 38 HLMs prepared from male livers (B). Also shown are the p values for Kruskal-Wallis one-way ANOVA on rank transformed data. In instances where the ANOVA indicated a significant effect, differences between individual diplotype groups were evaluated using Dunn's multiple comparison method (*, p < 0.05).
In addition to genotype, gender was found to have a significant modifying influence on S-oxazepam glucuronidation by HLMs with 67% higher median activities in livers from male donors compared with female donors. This finding corroborates well a previous report by Greenblatt et al. (1980) that showed 40% higher oxazepam clearance in vivo in healthy male (n = 18) compared with female (n = 20) subjects. Furthermore, it suggests that this gender dimorphism may result from higher levels of UGT2B15 in male compared with female livers, although this will need to be confirmed by demonstrating higher UGT2B15 protein and mRNA content. From a biological perspective, this difference is consistent with the known involvement of UGT2B15 in the glucuronidation and inactivation of male steroid hormones, including testosterone and dihydrotestosterone (Levesque et al., 1997).
Although we have shown that the D85Y polymorphism affects oxazepam glucuronidation, it is not yet apparent whether other clinically important substrates of UGT2B15, such as E-4-hydroxytamoxifen (Nishiyama et al., 2002) and 5-hydroxyrofecoxib (Zhang et al., 2003) will be affected to a similar degree. Indeed, our previous work using recombinant D85 and Y85 variants indicates that the magnitude of the effect could be substrate dependent in that a greater than 10-fold difference between variants was observed for naringenin glucuronidation, compared with a 5-fold difference for oxazepam glucuronidation, whereas there was less than a 2-fold difference for dihydrotestosterone glucuronidation (Court et al., 2002). Such substrate-dependent effects could be considered typical of amino acid variants that impact the enzyme active site. Furthermore, although we found no clear effect of K523T on oxazepam glucuronidation, this does not exclude the possibility that there could be effects of this cSNP on the glucuronidation of other substrates. Further work is needed to evaluate the effect of D85Y and the other identified cSNPs on the glucuronidation of other UGT2B15 substrates.
Finally, it should be pointed out that although gender and D85Y genotype seem to account for as much as 50% of the variability in S-oxazepam glucuronidation by human liver, the cause of at least one-half of the total variation remains unexplained. Possible sources might include genetic polymorphisms in the noncoding UGT2B15 gene introns, promoter and 5′-regulatory region, or in transcription factors regulating UGT2B15 gene expression. Environmental factors such as enzyme inducers present in the diet or administered as drugs could also play a role, although this has yet to be explored.
Footnotes
-
This work was supported by Grants GM-61834, DA-05258, MH-58435, DA-13209, DK-58496, AG-17880, AT-01381, and RR-00054 from the National Institutes of Health.
-
DOI: 10.1124/jpet.104.067660.
-
ABBREVIATIONS: UGT, UDP-glucuronosyltransferase; HLM, human liver microsome; HPLC, high-performance liquid chromatography; SNP, single nucleotide polymorphism; ANOVA, analysis of variance; cSNP, nonsynonymous single nucleotide polymorphism.
-
↵1 Current address: Department of Medicine and Pharmacology, Ohio State University, Columbus, OH 43210.
- Received February 26, 2004.
- Accepted March 25, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}