


Abstract
The aim of this study is to investigate the placental transport mechanism of cationic compounds by comparison of the uptake of an organic cation into human placental basal membrane vesicles (BLMVs) with that into organic cation transporter 3 (OCT3)-expressing cells. Reverse transcription-polymerase chain reaction analysis demonstrated that OCT3 is the only OCT isoform expressed in the human placenta. The function of OCT3 was investigated by measuring the uptake of 1-methyl-4-phenylpyridinium (MPP+) into human embryonic kidney (HEK)293 cells stably expressing OCT3 (HEK/OCT3 cells). The OCT3-mediated uptake of MPP+ was sodium- and chloride-independent and saturable, with a Michaelis constant (Km) of 82.5 μM. The OCT3-mediated uptake was inhibited by various cationic drugs in a concentration-dependent manner but not by anionic compounds, such as p-aminohippuric acid and captopril, or a zwitterion, carnitine. Western blotting analysis of membrane vesicles prepared from human term placenta revealed that OCT3 is expressed only in BLMVs but not in microvillous membrane vesicles. The uptake of MPP+ into BLMVs was membrane potential-dependent and saturable, with a Km value of 51.8 μM, which is similar to that in HEK293/OCT3 cells. The inhibitory spectrum of various compounds on MPP+ uptake by BLMVs was also similar to that in HEK293/OCT3 cells. These results suggest that OCT3 is expressed on the basal membrane of human trophoblast cells and plays an important role in the placental transport of cationic compounds.
Throughout gestation, the placenta plays an important role in regulating the supply of nutrients to the fetus, excretion of metabolic waste products from the fetus, and so on. In the placenta, trophoblast cells, which face the maternal blood, are considered to be the functional entity of the blood-placental barrier. Various transporters have been identified on both microvillous membrane and basal membrane, which face the maternal and fetal side, respectively, of trophoblast cells and are considered to regulate the exchange of various materials between mother and fetus.
Monoamines, including serotonin and norepinephrine, and cationic drugs, such as cimetidine and procainamide, are transported by organic cation transport systems in the kidney and liver (Koepsell, 1998; Kamisako et al., 1999; Dresser et al., 2000; Hohage and Gerhardt, 2000; Suzuki and Sugiyama, 2000). Rat organic cation transporter 1 (rOCT1) was first cloned from rat kidney as a component of the organic cation transport system (Gründemann et al., 1994). Whereas mouse and rat OCT1 are expressed in both liver and kidney (Gründemann et al., 1994; Schweifer and Barlow, 1996), hOCT1 is mainly expressed in the liver (Gorboulev et al., 1997; Zhang et al., 1999). It has also been shown by immunohistochemical study that rOCT1 is expressed on the sinusoidal membrane of hepatocytes (Meyer-Wentrup et al., 1998) and the basolateral membrane of renal tubular epithelium (Karbach et al., 2000; Sugawara-Yokoo et al., 2000).
rOCT2 was cloned as a homolog of rOCT1 from rat kidney (Okuda et al., 1996). Unlike hOCT1, OCT2 is expressed predominantly in the kidney (Okuda et al., 1996; Gorboulev et al., 1997) and is localized on the basolateral membrane of renal proximal tubules (Karbach et al., 2000; Sugawara-Yokoo et al., 2000).
Organic cation transporter 3 (OCT3) was first cloned from rat placenta (Kekuda et al., 1998), and its orthologues were also cloned from humans and mice (Gründemann et al., 1998; Verhaagh et al., 1999). Because hOCT3 has high affinity for monoamines, such as histamine, it is also designated as extraneuronal monoamine transporter (Gründemann et al., 1998). In contrast to OCT1 and 2, OCT3 is widely expressed (Kekuda et al., 1998; Verhaagh et al., 1999), although its expression is particularly high in the placenta (Verhaagh et al., 1999). Recently, it has been reported that pregnant OCT3-knockout mice exhibit reduced accumulation of MPP+ in the embryo compared with pregnant control mice, although the MPP+ concentration in placenta and amniotic fluid was not affected, suggesting that OCT3 mediates the transport of MPP+ from the placenta to the fetus but not from the maternal circulation (Zwart et al., 2001). These findings emphasize the importance of OCT3 in the placental transfer of cationic compounds, although its subcellular localization in the placenta and other tissues still remains unknown.
Besides OCTs, various transporters of organic cations have been identified in the human placenta. The expression of OCTN1, a member of a new subfamily of OCTs, has been reported in the human placenta, but its subcellular localization remains to be identified (Tamai et al., 1997). Another OCTN, OCTN2, has been demonstrated to be expressed on the maternal side (microvillous membrane) of trophoblast cells by Western blotting (Lahjouji et al., 2004). An organic cation/proton antiporter, norepinephrine transporter, serotonin transporter, and P-glycoprotein have been found on the microvillous membrane of trophoblast cells (Ganapathy et al., 2000; Ushigome et al., 2003), but the identity of the transporter of organic cations on the basolateral membrane remains unknown. The aim of this study is to investigate the placental transport mechanism of cationic compounds by comparison of the uptake of a model organic cation into human placental basal membrane vesicles (BLMVs) with that into OCT3-expressing cells.
Materials and Methods
Materials and Reagents. [3H]MPP+ (85.0 Ci/mmol), [3H]quinine (14.5 Ci/mmol), and [3H]dihydroalprenolol (60 Ci/mmol) were purchased from American Radiolabeled Chemicals (St. Louis, MO). [14C]Tetraethylammonium (2.4 Ci/mmol) was purchased from PerkinElmer Life and Analytical Sciences (Boston, MA). [3H]Theophylline (18.5 Ci/mmol) was purchased from Moravek Biochemicals (Brea, CA). Anti-OCT3 goat polyclonal antibody OCT3 (C-14) was purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA). Horseradish peroxidase-labeled anti-goat IgG antibody was purchased from Valeant Pharmaceuticals (Costa Mesa, CA). All other chemicals used in this study were commercial products of reagent grade.
Functional Expression of OCT3 in Mammalian Cells. Human OCT3 cDNA was obtained from human kidney total RNA by RT-PCR using KOD-plus-polymerase (Toyobo Engineering, Osaka, Japan). Sequence analysis revealed that the obtained OCT3 cDNA was 300 base pairs shorter than the OCT3 cloned previously (GenBank accession no. AJ001417; Gründemann et al., 1998). The missing part of the OCT3 cDNA was obtained by PCR using an Advantage-GC Genomic PCR kit (BD Biosciences Clontech, Palo Alto, CA) and human liver cDNA (BD Biosciences Clontech) under the following conditions: 1) 94°C × 3 min; 2) 94°C × 30 s, 68°C × 3 min (35 cycles); and 3) 68°C × 3 min. The full-length OCT3 cDNA was generated using the restriction enzyme BglII and subcloned into pIRESneo vector.
Sequencing of the cDNA was carried out with an ABI Prism BigDye Terminator Cycle Sequencing kit and 373 DNA sequencer (Applied Biosystems, Foster City, CA), and the sequence was analyzed by Sequencing Analysis 3.0 (Applied Biosystems). Sequence analysis showed that there was a G1260A mutation compared with the sequence already published by Gründemann et al. (1998), but it was a silent mutation.
An aliquot of 1 μg of OCT3/pIRESneo vector was transfected into HEK293 cells with LipofectAMINE 2000 Reagent (Invitrogen, Tokyo, Japan). After incubation for 24 h, cells were released by trypsin treatment and cultured in minimal essential medium containing 500 μg/ml geneticin. The cells were grown for 3 to 4 weeks and then used as HEK293 cells stably expressing OCT3 (HEK/OCT3 cells).
Preparation of Human Placental BLMVs and Microvillous Membrane Vesicles (BBMVs). Human placental BLMVs were prepared by the method of Inuyama et al. (2002) with minor modifications. Human term placentas from uncomplicated pregnancies were obtained within 15 min after vaginal or cesarean delivery and placed in 0.9% NaCl. After removal of the cord, amniochorion, and decidua, placental tissue was cut from the maternal side and washed in phosphate-buffered saline (PBS) (-). Tissue was stirred in PBS (-) for 30 min and collected on a nylon mesh. The filtrate was washed three times with ice-cold 50 mM Tris-HCl, pH 7.4, collected on a 250-μm pore size nylon mesh, and divided into several equal portions. Each portion was sonicated in 100 ml of the same Tris buffer using a 3/4-inch-high gain probe for 10 s at 240 W (Vibra-cell; Sonics and Materials, Newtown, CT). The suspensions were kept on ice. The sonication procedure selectively removes any remaining microvillous membrane. Sonicated tissue was collected on the mesh, washed three times with 5 mM Tris-HCl, pH 7.4, and then stirred gently for 60 min in the same buffer. Tissue was then collected on the nylon mesh and washed again in the same buffer. This procedure disrupts and removes the intracellular components, thus exposing the basolateral membranes. Tissue portions of 25 to 30 g were resuspended in approximately 100 ml of 50 mM Tris-HCl, pH 7.4, containing 10 mM EDTA and 250 mM sucrose and incubated for 30 min with occasional stirring. The portions were then sonicated twice for 20 s at 250 W to release the basolateral membranes. The suspensions were strained through nylon mesh, and the supernatant was centrifuged at 3430g for 10 min to remove debris. The supernatant from this spin was recentrifuged at 80,000g for 40 min to yield the basolateral membrane pellet, which was resuspended, using a Dounce homogenizer in 25 mM HEPES-Tris, pH 7.4, containing 1 mM EDTA and 275 mM sucrose. This fraction was further purified by centrifugation on a discontinuous gradient of 10% (w/v) Ficoll (Pfizer, Inc., New York, NY) in the resuspension buffer overlaid with 4% Ficoll (as described by Kelley et al., 1983) prepared in 25 mM HEPES-Tris, pH 7.4, containing 1 mM EDTA and 275 mM sucrose. Ficoll gradient tubes were spun at 90,000g for 6 to 8 h. The material at the density gradient interfaces was collected, washed, and resuspended in 25 mM HEPES-Tris, pH 7.4, containing 275 mM sucrose. The suspension from this run was resuspended in 25 mM HEPES-Tris buffer containing 150 mM KCl, pH 7.4 (E buffer) with a 25-gauge syringe needle. All of the operations were carried out at 4°C.
BBMVs were prepared according to the method described by Nakamura et al. (2002) with minor modifications. Human term placentas from uncomplicated pregnancies were obtained within 15 min after vaginal or cesarean delivery and placed in 0.9% NaCl. After removal of the cord, amniochorion, and decidua, placental tissue was cut from the maternal side and washed in 250 mM mannitol, 10 mM HEPES-Tris at pH 7.4 (MHT buffer). The mince was stirred for 1 h to loosen the microvilli and filtered through two layers of woven cotton gauze. A sample of this starting mince was taken for enzyme analysis. The filtrate was centrifuged at 800g for 10 min. The pellet was discarded, and MgCl2 was added to the supernatant to a final concentration of 10 mM. After 10 min, with occasional stirring, the supernatant was centrifuged at 10,500g for 10 min. The pellet was discarded, and the supernatant was centrifuged at 20,000g for 20 min. The pellet from this run was suspended in E buffer with a 25-gauge syringe needle. All of the subsequent procedures were performed at 4°C. BLMVs and BBMVs were quickly frozen and stored at -80°C and used within a month.
Tissue homogenate was prepared by the method previously described by Kelley et al. (1983). Approximately 3 g of whole villous tissue was homogenized in 10 ml of buffer E using a Waring blender (PHYSCOTRON Micro Teq.; Nichion Co., Chiba, Japan) for 2.25 min and further with a homogenizer for eight strokes. The material was filtered through six layers of gauze.
Binding activity of [3H]dihydroalprenolol as a marker of the basal membrane and alkaline phosphatase (ALP) activity as a marker of the microvillous membrane was assayed as reported by Kelley et al. (1983) and Bessey et al. (1946), respectively. The dihydroalprenolol binding of BLMVs was 25.2-fold higher than that of the homogenate, whereas the ALP activity was only 3.07-fold higher. On the other hand, the dihydroalprenolol binding of BBMVs was only 1.07-fold higher than that of the homogenate, whereas the ALP activity was 18.3-fold higher. The amount of protein in the sample was measured by the method of Lowry et al. (1951).
RT-PCR Analysis of OCTs mRNA in Human Placenta and BeWo Cells. The total RNAs of BeWo cells and human placenta were extracted with an RNeasy mini kit (QIAGEN GmbH, Hilden, Germany). Human liver total RNA was purchased from Cell Applications, Inc. (San Diego, CA), and human kidney total RNA was purchased form Stratagene (La Jolla, CA). First-strand cDNA was synthesized from total RNA (50 μg), using random primer and SuperScript II reverse transcriptase (Invitrogen, Carlsbad, CA). PCR was performed with a T-Gradient Thermoblock (Biometra, Göttingen, Germany) using KOD-plus-polymerase (Toyobo Engineering) and primers specific for each family member. Table 1 shows the primer sets used in RT-PCR and the accession number of each transporter.
Oligonucleotides used for RT-PCR Open reading frame positions of OCT1, 2, and 3 are 1-1665, 1-1668, and 1-1671, respectively.
Western Blotting. HEK293/OCT3 cells, BLMVs, and BBMVs were collected and suspended in lysis buffer containing 100 mM Tris-HCl, pH 7.6, 150 mM NaCl, 1 mM CaCl2, 1% Triton X-100, 0.1% SDS, 0.1% Nonidet P-40, 1 mM phenylmethylsulfonyl fluoride, 0.01 mg/ml leupeptin, 0.01 mg/ml aprotinin, and 1 mM sodium vanadate and incubated for 30 to 45 min at 4°C. After incubation, the suspension was centrifuged at 15,000g for 15 min at 4°C. SDS-polyacrylamide gel electrophoresis was performed according to the method of Laemmli (1970). The proteins were transferred electrophoretically onto a 0.2-μm pore size Clear Blot Membrane-P (Atto Corporation, Tokyo, Japan). Blots were blocked overnight at 4°C with 5% nonfat powdered milk in PBS (-). OCT3 (C-14) was used as the primary antibody for OCT3, and horseradish peroxidase anti-goat IgG (Valeant Pharmaceuticals) was used as the secondary antibody. Detection was done with enhanced chemiluminescence reagents (GE Healthcare, Little Chalfont, Buckinghamshire, UK) according to the instructions of the manufacturer.
Uptake Experiment into HEK/OCT3 Cells. HEK/OCT3 cells were seeded at 5 × 104 cells/well on 96-well plates (NUNC A/S, Roskilde, Denmark) and grown for 2 days until used for the uptake study.
After reaching confluence, cells were washed twice with uptake buffer (125 mM NaCl, 4.8 mM KCl, 5.6 mM d-glucose, 1.2 mM KH2PO4, 1.2 mM MgSO4, and 25 mM HEPES, pH 7.4) before the study. Uptake was initiated by adding 100 μl of uptake buffer containing unlabeled MPP+ and 10 nM [3H]MPP+. For the Na+ dependence study, NaCl was replaced with LiCl, mannitol, or sodium gluconate. After incubation at 37°C for an appropriate time, uptake was terminated by aspiration of the buffer followed by two washes with 1 ml of ice-cold uptake buffer. Cells were then dissolved in 150 μl of 1 N NaOH, and the solution was neutralized with 150 μl of 1 N HCl. Scintillation cocktail (Clear-sol I; Nacalai Tesque, Kyoto, Japan) was added, and the radioactivity of [3H]MPP+ was determined with a liquid scintillation counter (LS6500; Beckman Coulter, Fullerton, CA). The amount of protein in the cells was measured by the method of Lowry et al. (1951).
Measurement of Uptake into BLMVs and BBMVs. The uptake of [3H]MPP+ into membrane vesicles was measured by using a rapid filtration technique (Russel et al., 1988). Uptake was initiated by the addition of 90 μl of incubation buffer to 10 μl of BLMV suspension containing 40 to 50 μg of protein. The incubation buffer consisted, in general, of unlabeled MPP+, 25 mM HEPES-Tris, pH 7.4, 150 mM NaCl, and [3H]MPP+ (0.08 μCi/point).
At the designated time, uptake was terminated by adding 1 ml of ice-cold stop buffer followed immediately by filtration (HAWP 0.45 μm; Millipore Corporation, Billerica, MA). The filter was washed twice with 4 ml of ice-cold stop buffer. In general, stop buffer contained 10 mM sucrose in incubation buffer. Nonspecific binding was determined by adding 1 ml of ice-cold stop solution and 90 μl of ice-cold incubation buffer to the ice-cold BLMV suspension followed by the same treatment as in the uptake experiments.
To assay radiolabeled compounds, filters were placed in counting vials and mixed with 4 ml of scintillation fluid, Clear-sol I. Radioactivity was measured with a liquid scintillation counter (LS6500).
Data Analysis. The [3H]MPP+ uptake into HEK/OCT3 cells is expressed as the cell-to-medium ratio calculated from the intracellular uptake per milligram of protein (disintegrations per minute per milligram of protein) of the cells relative to the initial drug concentration (disintegrations per minute per microliter).
As for the uptake into membrane vesicles, obtained radioactivity was normalized with respect to the protein amount of vesicles. Values were determined by subtracting nonspecific binding from total uptake (in the investigation of osmolarity effects as well), and data are presented as the vesicle-to-medium ratio (microliters per milligram of protein).
To determine the kinetic parameters, Kt, Jmax, and kd, the following Michaelis-Menten equation was fitted to the data using the nonlinear least-squares regression analysis program MULTI (Yamaoka et al., 1981): J = Jmax× S/(Kt + S) + kd × S, where J and S represent the transport rate and concentration of substrate, respectively. Jmax (nanomoles per milligram of protein per 30 s), Kt (millimolar), and kd (microliters per milligram of protein per 30 s) represent the maximum uptake rate for a carrier-mediated process, the Michaelis constant, and the rate constant for the nonsaturable component, respectively.
Comparisons between two and among more than three groups were performed with the unpaired Student's t test and with nonrepeated analysis of variance (ANOVA) followed by Dunnett's test, respectively. The Spearman rank correlation test was used to determine the degree of association between the inhibitory effects on MPP+ uptake into HEK/OCT3 cells and BLMVs. A P value of less than 0.05 was considered statistically significant.
Results
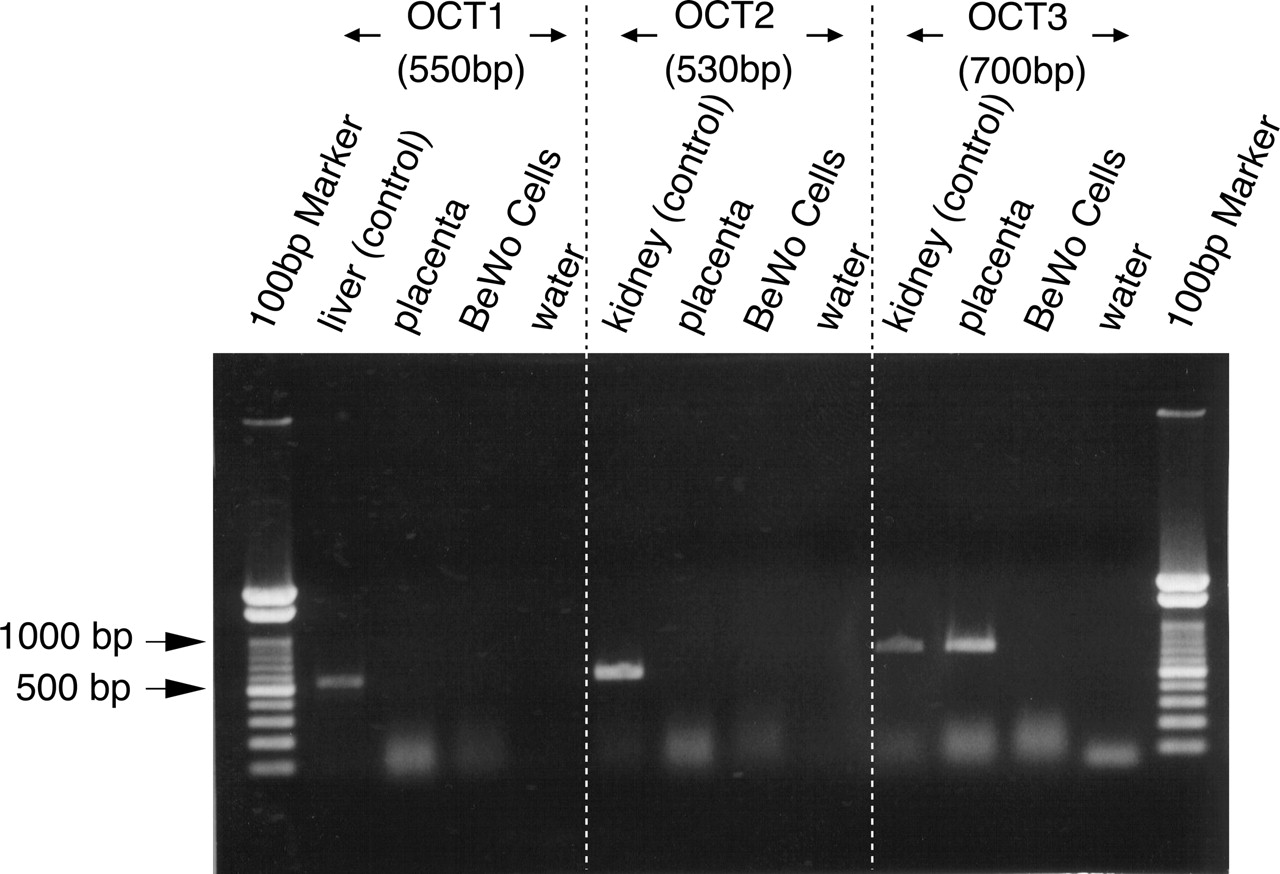
Expression of OCT mRNAs in Human Placenta and BeWo Cells. The expression of OCTs in human placenta and BeWo cells was examined by RT-PCR analysis. Total RNAs from human liver and kidney were used as positive controls for OCT1 and OCT2 and 3, respectively. In placenta, only OCT3 and the positive control were detected (Fig. 1). The PCR product was confirmed to be in accord with the sequence of human OCT3. With regard to BeWo cells, no band was detected, suggesting that OCTs are not expressed in these cells.
Expression of OCT3 Protein in HEK/OCT3 Cells and Human Placenta. The expression of OCT3 was examined by Western blotting analysis of HEK/OCT3 cells and mock cells (HEK293 cells transfected with vector alone). As expected, a band of 70 kDa was detected only in HEK/OCT3 cells (Fig. 2A).
The expression of OCT3 on human placental trophoblast membrane was also examined by Western blotting analysis using BLMVs and BBMVs prepared from human placenta. The band corresponding to OCT3 was detected in BLMVs at about 70 kDa, as well as in HEK/OCT3 cells, but not in BBMVs (Fig. 2B).
Time Course of the Uptake of MPP+. The uptake of 1 μM MPP+ into HEK293/OCT3 cells was significantly higher than that into mock cells (Fig. 3). This uptake was linear for at least 2 min and was attenuated at 4°C. In the following experiments, the initial uptake rate was determined at 30 s.
Effects of Extracellular Ion Composition on the Uptake of MPP+. The effects of Na+ and Cl- on the uptake of MPP+ were examined by isoosmotic replacement of NaCl (Na+, +; Cl-, +) with mannitol (Na+, -; Cl-, -), sodium gluconate (Na+, +; Cl-, -), and LiCl (Na+, -; Cl-, +) in the uptake buffer. As shown in Fig. 4, the uptake of MPP+ was not affected by any replacement of NaCl, suggesting that OCT3-mediated uptake of MPP+ is independent of both Na+ and Cl-.
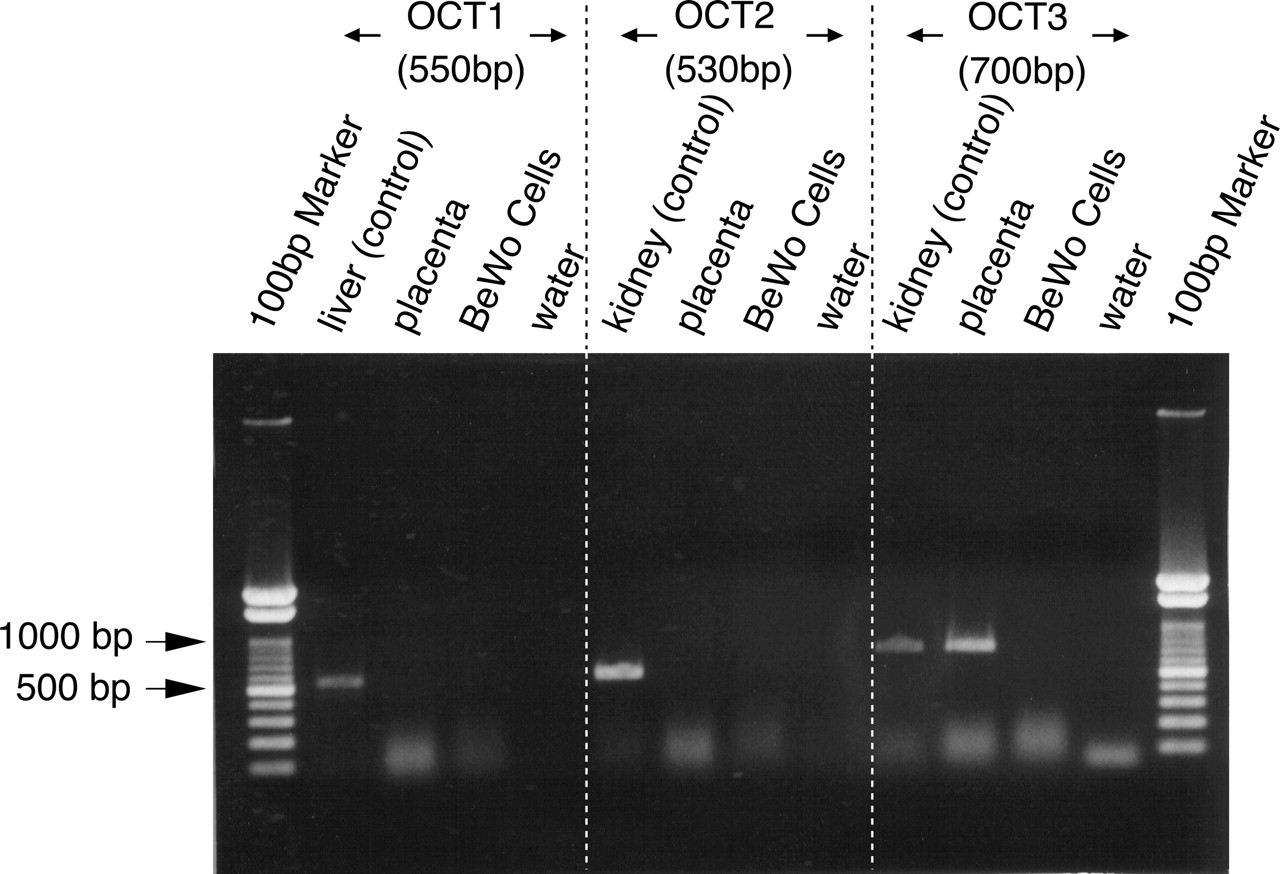
RT-PCR analysis of the distribution of OCT mRNAs in the human placenta and BeWo cells. Total RNAs from the placenta, liver, kidney, and BeWo cells were reverse-transcribed and used for PCR amplification with oligonucleotide primers specific for OCT1, 2, and 3.

Western blotting analysis of OCT3 in HEK/OCT3 cells and human placenta. Ten micrograms each of HEK/OCT3 cells and mock (A) and 50 μg each of BLMVs and BBMVs (B) were resolved by SDS-polyacrylamide gel electrophoresis with a 10% polyacrylamide gel and transferred onto Clear Blot Membrane-P. Immunoblots were performed with OCT3-C-14 and anti-β-actin mouse monoclonal antibody and developed with the enhanced chemiluminescence detection reagent. OCT3 and β-actin were detected at 70 and 40 kDa, respectively.

Time course of OCT3-mediated uptake of 1 μM MPP+. HEK/OCT3 cells were incubated at 37 (closed circle) and 4°C (closed triangle). HEK293 cells transfected with vector alone (open circle) were incubated at 37°C. Each point represents the mean ± S.E.M. of four determinations. *, P < 0.05 versus control (Student's t test).
Concentration-Dependent Uptake of MPP+. The OCT3-mediated uptake was saturable with a Michaelis constant (Kt) of 82.5 μM (59.7-113.8) [mean (mean - S.D. - mean + S.D.)] and a maximal uptake velocity (Jmax) of 2538 ± 567.4 pmol/mg protein/30 s (mean ± S.D.) (Fig. 5) obtained from three separate experiments.
Inhibitory Effects of Various Cationic Compounds on the OCT3-Mediated Uptake of MPP+. The OCT3-mediated uptake of 10 μM MPP+ was significantly inhibited by various cationic drugs, such as cimetidine, ranitidine, verapamil, quinine, quinidine, imipramine, trimethoprim, and procainamide, in a concentration-dependent manner (Figs. 6 and 7). In contrast, anionic compounds [p-aminohippuric acid (PAH), captopril, and phenobarbital], a zwitterion (carnitine), and a cationic compound (lamivudine) showed little or no inhibitory effect.

OCT3-mediated uptake of MPP+ in the presence or absence of Na+ or Cl-. HEK/OCT3 cells were incubated at 37°C for 30 s with 10 μM MPP+. Uptake buffers of different ionic composition were used: NaCl (Na+, +; Cl-, +), mannitol (Na+, -; Cl-, -), sodium gluconate (Na+: +, Cl-: -), LiCl (Na+, -; Cl-, +). Each point represents the mean ± S.E.M. of four determinations.

Concentration-dependent uptake of MPP+ into HEK/OCT3 cells. Uptake of MPP+ into HEK/OCT3 cells and mock was measured at concentrations between 1 μM and 3 mM. Data are presented as the OCT3-specific uptake calculated by subtracting the uptake obtained with mock from that obtained with HEK/OCT3. Each point represents the mean ± S.E.M. obtained from three different experiments.
Uptake of Various Cationic Compounds. To search for novel substrates of OCT3, we examined the uptake of 10 μM tetraethylammonium, quinine, theophylline, and ramosetron into HEK/OCT3 cells. No OCT3-mediated uptake of these compounds was observed (data not shown).
Uptake of MPP+ into BLMVs. The vesicle-to-medium ratio of MPP+ in BLMVs was linear for at least 30 s (Fig. 8). The vesicle-to-medium ratio at 10 min was reduced with increasing extracellular osmolarity, suggesting that MPP+ was not only bound to the vesicles but also was taken up into the vesicles.
This uptake was significantly attenuated by replacing extravesicular NaCl with KCl and was potentiated by the addition of valinomycin, a potassium ionophore. The uptake of MPP+ was not affected by the replacement of NaCl with LiCl (Fig. 9). These results suggested that the uptake of MPP+ into BLMVs is dependent on membrane potential and independent of Na+.
Concentration-Dependent Uptake of MPP+ into BLMVs. The uptake of MPP+ into BLMVs was saturable with a Michaelis constant (Kt) of 51.8 μM [34.9-113.8] (mean [mean - S.D. - mean + S.D.]) and a maximal uptake velocity (Jmax) of 332 ± 30.8 pmol/mg protein/30 s (mean ± S.D.) (Fig. 10) obtained from three separate experiments.

Inhibitory effects of several drugs on the uptake of MPP+ into HEK/OCT3 cells. HEK/OCT3 cells were incubated at 37°C for 30 s with 10 μM MPP+ in the absence or presence of 1 mM various compounds. Results are given as percentage of control uptake measured in the absence of inhibitors. The control and control [+0.5% dimethyl sulfoxide (DMSO)] values were 27.38 and 25.04 μl/mg protein/30 s. Each point represents the mean ± S.E.M. of four determinations. *, P < 0.05 versus respective control (ANOVA; Dunnett's test).
Effects of Various Compounds on the Uptake of MPP+ into BLMVs. We examined the effects of various compounds of the uptake of MPP+ into BLMVs. We selected noninhibitors (PAH, captopril, and lamivudin), modest inhibitors (carnitine and procainamide), and strong inhibitors (cimetidine, imipramine, quinidine, verapamil, and quinine) of OCT3-mediated uptake. As shown in Fig. 11, PAH and procainamide showed little or no inhibitory effect, whereas quinine, quinidine, imipramine, and verapamil inhibited the uptake of MPP+ into BLMVs. The rank order of the inhibitory effects of MPP+ uptake into HEK/OCT3 cells was highly correlated with that in BLMVs (r2 = 0.688, P < 0.05; Fig. 12).
Discussion
To investigate the placental transport mechanism of cationic compounds, we focused on OCTs in this study. Because the only OCT detected in human placenta by RT-PCR analysis was OCT3, this molecule is expected to play an important role in the transport of cationic compounds across the human placenta.
hOCT1 and hOCT2 are predominantly expressed in the liver and kidney, respectively (Gorboulev et al., 1997). Unlike OCT1 and OCT2, OCT3 is widely expressed (Kekuda et al., 1998; Verhaagh et al., 1999). The OCT3 mRNA level in the placenta is particularly high, being 2.5 and 5 times that in the liver and kidney, respectively. OCT3 is considered to be one of the most abundantly expressed transporters in the placenta (Leazer and Klaassen, 2003). Our results are consistent with that finding and underscore the key role of OCT3 in the transport of cationic compounds in the placenta.
We also investigated OCTs in a human choriocarcinoma trophoblast cell line, BeWo cells, and found no expression of OCTs. This finding is in accordance with the results of Wu et al. (2000). Although the BeWo cell line originated from placenta, it has the characteristics of cytotrophoblast cells and is functionally and morphologically distinct from normal syncytiotrophoblast cells (Ugele and Simon, 1999). Thus, it is not surprising that OCT3 is not expressed in BeWo cells.

Inhibitory effects of several drugs on the uptake of MPP+ into HEK/OCT3 cells. HEK/OCT3 cells were incubated at 37°C for 30 s with 10 μM MPP+, in the absence or presence of 10 or 100 μMor 1 mM inhibitors. Results are given as percentage of control uptake measured in the absence of inhibitors. The control and control [+0.5% dimethyl sulfoxide (DMSO)] values were 29.97 μl/mg protein/30 s and 28.54 μl/mg protein/30 s. Each point represents the mean ± S.E.M. of four determinations. *, P < 0.05 versus respective control (ANOVA; Dunnett's test).

Influence of Na+ on the uptake of MPP+ into BLMVs. Uptake was initiated by the addition of 10 μM MPP+ in the presence of Na+ 100 mM/K+ 3 mM (closed circle) or Na+ 3 mM/K+ 100 mM (open circle). Each point represents the mean ± S.E.M. of three determinations. *, P < 0.05 versus control (Student's t test).
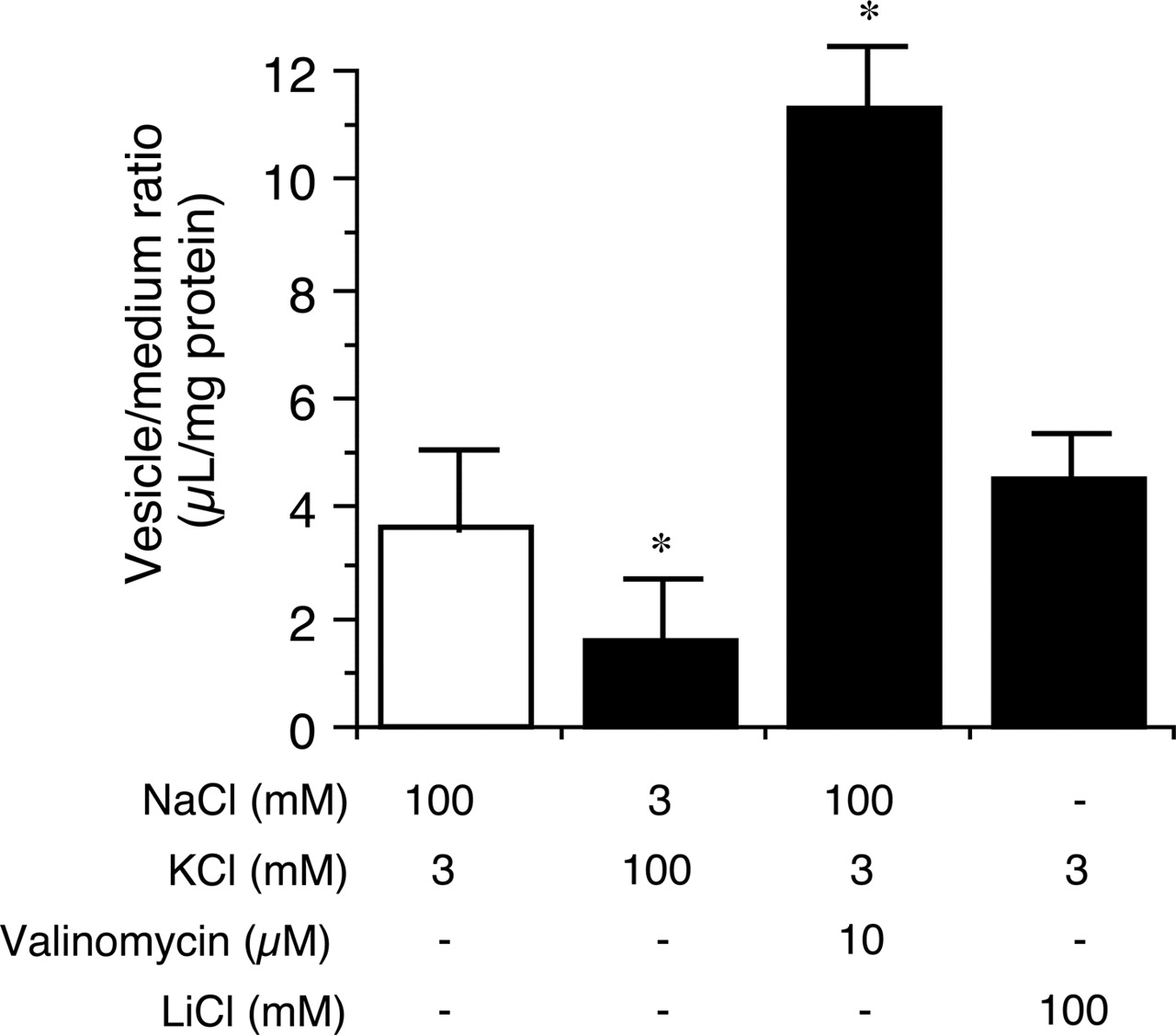
Influence of membrane potential on the uptake of MPP+ into BLMVs. Uptake was initiated by the addition of 10 μM MPP+. Ionic composition of uptake buffers is shown below the figure. Each point represents the mean ± S.E.M. of three determinations. *, P < 0.05 versus control (ANOVA; Dunnett's test).
Western blotting demonstrated predominant expression of OCT3 in HEK/OCT3 cells. It was detected as a band of 70 kDa, which is larger than expected from the amino acid sequence (62 kDa) (Fig. 2). Because both mouse and rat OCT3 have four to five N-glycosylation sites (Burckhardt and Wolff, 2000) and hOCT3 shows 90% homology to them, the difference in size is likely to be attributable to glycosylation. Indeed, the size of the bands of OCT3 from both BLMVs and HEK/OCT3 cells was 70 kDa. The established HEK/OCT3 cells transported MPP+ with a Kt value of 69 μM (Fig. 5), which is in good agreement with the values reported for human retinal pigment epithelial cells (47 μM; Wu et al., 2000) and HEK293 (104 μM; Martel et al., 2001).

Concentration-dependent uptake of MPP+ into BLMVs. Uptake of MPP+ into BLMVs was measured at concentrations between 1 μM and 1 mM. Uptake was initiated by the addition of 10 μM MPP+ in the presence of Na+ 100 mM/K+ 3 mM. Each point represents the mean ± S.E.M. obtained from three different experiments.

Inhibitory effects of several drugs on the uptake of 10 μM MPP+ into BLMVs. BLMVs were incubated at 37°C for 30 s with 10 μM MPP+, in the absence or presence of 1 mM various compounds. Results are given as percentage of control uptake measured in the absence of inhibitors. The control values were 4.02 μl/mg protein/30 s. Each point represents the mean ± S.E.M. of three determinations. *, P < 0.05 versus control (ANOVA; Dunnett's test).
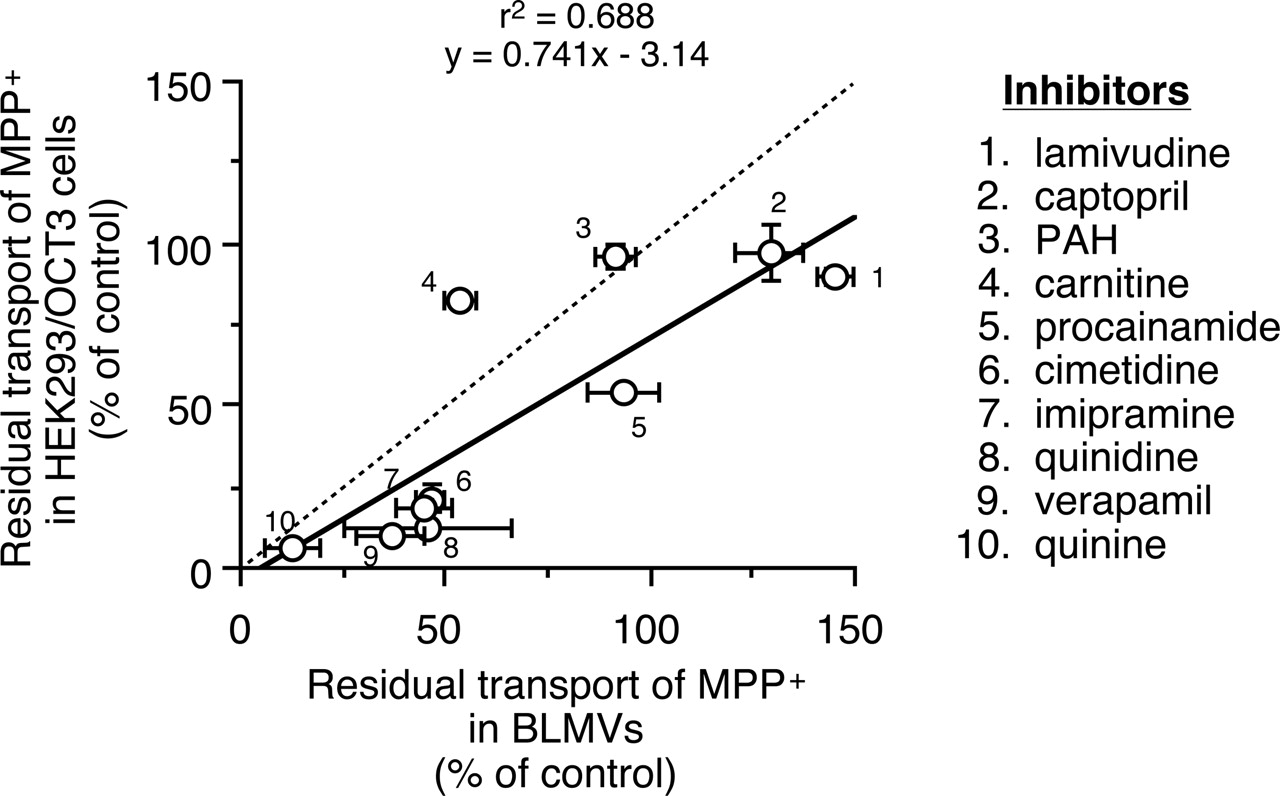
Comparison of inhibitory effects on MPP+ uptake between HEK/OCT3 cells and BLMVs. The solid line shows the linear regression and the dashed line shows the 1:1 line. The concentration of each inhibitor was 1 mM. Each point represents the mean ± S.E.M. of three (BLMVs) or four (HEK/OCT3 cells) determinations.
The uptake of MPP+, a typical substrate of OCT3, into HEK/OCT3 cells was inhibited by cationic compounds, such as cimetidine, verapamil, quinine, quinidine, ramosetron, and trimethoprim, in a concentration-dependent manner. In contrast, anionic compounds, including PAH, captopril, and phenobarbital, and a zwitterion, carnitine, showed little or no inhibitory effect (Figs. 6 and 7). In addition to well known inhibitors of OCT3, such as cimetidine, imipramine, and quinine (Gründemann et al., 1998; Wu et al., 2000), we have identified a variety of cationic drugs that inhibit OCT3. Interestingly, a cationic compound, lamivudine, did not inhibit OCT3 function, suggesting that cationic character alone is not sufficient for recognition by OCT3. Among therapeutic drugs, only cimetidine is known to be a substrate of OCT3 (Gründemann et al., 1999). Because various cationic drugs were found to inhibit OCT3, some of them might be substrates of OCT3.
To search for novel substrates of OCT3, we also examined the uptake of tetraethylammonium, quinine, theophylline, and ramosetron into HEK/OCT3 cells. Although tetraethylammonium is a typical substrate of OCT1 and 2 as well as MPP+, none of the above-mentioned drugs was a substrate for OCT3. Our finding in the case of tetraethylammonium is consistent with previous reports that the affinity of tetraethylammonium for mouse and rat OCT3 is approximately 10-fold weaker than that for OCT1 and 2 (Kekuda et al., 1998; Wu et al., 2000) and that hOCT3 does not interact with tetraethylammonium (Gründemann et al., 1999). In contrast, OCT3 has high affinity for monoamines (Gründemann et al., 1998). Overall, OCT3 seems to have a substrate specificity distinct from those of OCT1 and 2.
Western blot analysis of BLMVs and BBMVs showed that OCT3 is predominantly expressed on the basolateral membrane of trophoblast cells (Fig. 2). Recently, it has been reported that accumulation of MPP+ in the embryo was reduced in OCT3-knockout pregnant mice, although MPP+ concentrations in the placenta and amniotic fluid were comparable (Zwart et al., 2001), suggesting that OCT3 mediates the transport of MPP+ from the placenta to the fetus but not from the maternal circulation to the placenta. This finding is consistent with our results that OCT3 is expressed on the basolateral membrane of trophoblast cells. Although various transporters, such as organic cation/proton antiporter, norepinephrine transporter, and serotonin transporter, were shown to be localized at the microvillous membrane of placental trophoblast cells (Ganapathy et al., 2000), nothing was known about the expression of transporters for cationic compounds on the basolateral membrane of trophoblast cells. Herein, we have demonstrated for the first time the expression and function of OCT3 on the basal membrane, where it presumably plays a role in the excretion of metabolic waste products or xenobiotics from the fetus. With regard to the transport direction, OCT3 has been considered to transport cationic compounds bidirectionally in a concentration-dependent manner (Kekuda et al., 1998). OCT3 may transport the maternally administered cationic compounds from the placenta to the fetal side and transport oppositely the cationic compounds whose level is higher in the fetal blood.
We compared quantitatively the transport properties of cationic compounds in human placental membrane vesicles with those of HEK/OCT3 cells. The uptake of MPP+ into BLMVs was membrane potential-sensitive and Na+-independent (Fig. 9). The Km values of MPP+ uptake into BLMVs and HEK/OCT3 cells were similar (39 and 70 μM, respectively). Moreover, the inhibitory effects of various compounds on the uptake of MPP+ into BLMVs were highly correlated with those on uptake into HEK/OCT3 cells (Fig. 12). Because we did not examine detailed mechanism for the inhibition by drugs, we cannot exclude the possibility that they inhibited OCT3 in a noncompetitive manner by a nonspecific mechanism, such as membrane depolarization and so on. Although the inhibitory mechanism by these compounds remains to be further investigated, the inhibitory nature is unlikely to significantly affect the conclusion that OCT3 plays an important role in the placental transport of cationic compounds.
In summary, we have identified the expression of OCT3 on the basal membranes of trophoblast cells. Because the properties of MPP+ uptake into BLMVs are similar to those in HEK/OCT3 cells, OCT3 is likely to be predominantly responsible for the transport of cationic compounds across the basolateral membranes of trophoblast cells.
Footnotes
-
This study was supported in part by a Grant-in-aid for Young Scientists (A) from the Ministry of Education, Culture, Sports, Science and Technology of Japan.
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.105.086827.
-
ABBREVIATIONS: rOCT, rat organic cation transporter; OCT, organic cation transporter; hOCT, human organic cation transporter; BLMV, human placental basolateral membrane vesicle; MPP+, 1-methyl-4-phenylpyridinium; RT-PCR, reverse transcription-polymerase chain reaction; HEK, human embryonic kidney; BBMV, human placental microvillous membrane vesicle; PBS, phosphate-buffered saline; ALP, alkaline phosphatase; ANOVA, analysis of variance; PAH, p-aminohippuric acid.
- Received March 23, 2005.
- Accepted August 2, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}