


Abstract
Chronic renal failure (CRF) is associated with an increased bioavailability of drugs by a poorly understood mechanism. One hypothesis is a reduction in the elimination of drugs by the intestine, i.e., drug elimination mediated by protein membrane transporters such as P-glycoprotein (Pgp) and multidrug-resistance-related protein (MRP) 2. The present study aimed to investigate the repercussions of CRF on intestinal transporters involved in drug absorption [organic anion-transportingpolypeptide (Oatp)] and those implicated in drug extrusion (Pgp and MRP2). Pgp, MRP2, MRP3, Oatp2, and Oatp3 protein expression and Pgp, MRP2, and Oatp3 mRNA expression were assessed in the intestine of CRF (induced by five-sixth nephrectomy) and control rats. Pgp and MRP2 activities were measured using the everted gut technique. Rat enterocytes and Caco-2 cells were incubated with sera from control and CRF rats to characterize the mechanism of transporters' down-regulation. Protein expression of Pgp, MRP2, and MRP3 were reduced by more than 40% (p < 0.01) in CRF rats, whereas Oatp2 and Oatp3 expression remained unchanged. There was no difference in the mRNA levels assessed by real-time polymerase chain reaction. Pgp and MRP2 activities were decreased by 30 and 25%, respectively, in CRF rats compared with control (p < 0.05). Uremic sera induced a reduction in protein expression and in activity of drug transporters compared with control sera. Our results demonstrate that CRF in rats is associated with a decrease in intestinal Pgp and MRP2 protein expression and function secondarily to serum uremic factors. This reduction could explain the increased bioavailability of drugs in CRF.
Dose adjustment of kidney-excreted drugs is usually done according to the glomerular filtration rate. However, despite dosage adjustment, patients with CRF present a great number of drug adverse side effects (Matzke and Frye, 1997). Part of this phenomenon is related to the fact that CRF also affects the nonrenal route of elimination of drugs, i.e., hepatic and intestinal metabolisms of drugs (Touchette and Slaughter, 1991; Matzke and Frye, 1997; Pichette and Leblond, 2002, 2003; Nolin et al., 2003). Drug accumulation in CRF could also be explained by alterations in the nonmetabolic clearance of drugs by the liver and the intestine, i.e., drug elimination mediated by protein membrane transporters such as P-glycoprotein (Pgp) and multidrug resistance-related protein (MRP) type 2.
Oral bioavailability, i.e., the quantity of drug reaching the blood circulation, depends on different factors, the two most important being presystemic metabolism (intestinal and hepatic) and intestinal drug transport (Hall et al., 1999; Nolin et al., 2003). Presystemic metabolism is primarily associated with intestinal and liver P450, whereas drug extrusion and import are mediated by several membrane proteins (transporters). The most important intestinal extrusion transporters are Pgp and MRP2, whereas the most important intestinal import transporter is the organic anion-transporting polypeptide (Oatp) type 3. Any modification in these systems will have important repercussions in the bioavailability of xenobiotics (Hall et al., 1999).
The effects of CRF on the bioavailability of drugs are poorly documented; however, several studies have shown an increase in the bioavailability of many drugs, such as erythromycin, dextropropoxyphene, dihydrocodeine, oxprenolol, propranolol, tolamolol, and tacrolimus in patients with CRF (Talbert, 1994; Matzke and Frye, 1997; Okabe et al., 2000). This could be related to a decrease in intestinal extrusion or presystemic metabolism of these drugs. We have demonstrated that intestinal activity and expression of P450 are decreased by 40% in rats with CRF (Leblond et al., 2002). On the other hand, very few studies have described the repercussions of CRF on intestinal drug transporters. Veau et al. (2001) reported a decrease in intestinal transport of Rho-123, pointing to an inhibition of intestinal Pgp activity. Furthermore, Huang et al. (2000) demonstrated that the incubation of Caco-2 cells with serum from rats with acute renal failure decreases Pgp-mediated transport of Rho-123 (Huang et al., 2000), suggesting that serum from animals with acute renal failure contains mediators that could down-regulate Pgp. However, the mechanism leading to decreased Pgp activity and the effects of CRF on other intestinal drug transporters are unknown.
The objectives of this study were to determine the effects of CRF on some major intestinal drug transporters and to define the mechanisms leading to their down-regulation. To achieve this goal, we measured the following in CRF and control rats: intestinal Pgp, MRP2, MRP3, Oatp2, and Oatp3 protein expression; Pgp, MRP2, and Oatp3 mRNA expression; and intestinal Pgp and MRP2 activities using specific substrates. Finally, rat enterocytes and Caco-2 cells were incubated with sera from control and CRF rats to determine the effects of uremic factors on intestinal drug transporters and to characterize the mechanism by which these transporters are down-regulated.
Materials and Methods
Experimental Models. All reagents were of ACS grades or higher and, unless otherwise stated, purchased from Sigma Chemicals (St. Louis, MO). Male Sprague-Dawley rats (Charles River, St. Constant, QC, Canada) weighing 200 to 300 g were housed in the Research Center animal care facility. All the experiments were conducted according to the Canadian Council on Animal Care guidelines for care and use of laboratory animals.
Studies were performed in two groups of 25 animals each: control pair-fed and CRF. Protein expression of drug transporters and mRNA levels as well as activities of drug transporters were assessed in the two groups (n = 10 in both groups). Immunochemistry was done in 10 rats (five CRF and five controls), whereas in situ hybridization studies were done in some of these rats. To evaluate the effects of uremic sera, sera from 16 rats were used (eight CRF and eight controls).
Chronic renal failure was induced by two-stage five-sixth nephrectomy, as described previously (Leblond et al., 2001). Control pair-fed rats were fed the same amount of rat chow that was ingested by the CRF rats on the previous day. Rats were sacrificed 42 days after nephrectomy.
Intestinal Membrane Protein Preparation. Intestinal membrane proteins were prepared from enterocytes obtained by elution, based on the methods of Weiser et al. as previously published (Weiser, 1973; Fasco et al., 1993). The third collected fraction was rapidly pelleted and frozen in liquid nitrogen to be used for mRNA level assessment. The six remaining fractions were pooled and homogenized in 0.9% NaCl and 0.1 mM phenylmethylsulfonyl fluoride using a Potter-Elvehjem tissue grinder (Wheaton Science Products, Millville, NJ). Aliquots were stored at -80°C until Western blot analysis.
Western Blot Analysis. Drug transporters Pgp, MRP2, MRP3, Oatp2, and Oatp3 protein expression were assessed by Western blot analysis. Every blot was repeated three times, and results were pooled to obtain the final reported values. The major intestinal cytochrome P450 isoform implicated in the metabolism of drugs in rats (CYP3A2) was also assessed by Western blot analysis as described previously (Leblond et al., 2002).
Pgp and MRP2 were detected using mouse monoclonal antibodies from ID Labs (London, ON, Canada): C219 and M2 III-6, respectively. MRP3 was detected using a rabbit anti-human MRP3, which cross-reacts with rat MRP3 from Sigma. Oatp2 and Oatp3 were detected using rabbit anti-rat Oatp2 and Oatp3 antibody from Alpha Diagnostic International (San Antonio, TX) and U.S. Biological (Swampscott, MA), respectively. Villin (a protein specific to mature enterocytes) was also measured by Western blot using a mouse anti-chicken villin antibody (Chemicon International, Temecula, CA). Villin was used as a loading control for Western analysis. In preliminary experiments, we have demonstrated that the expression of villin was not modified in CRF rats compared with controls (data not shown).
RNA Isolation and Real-Time PCR. Total RNA was extracted from the third eluted fraction of enterocytes using TRIzol reagent (Invitrogen, Burlington, ON, Canada). RNA concentrations were determined by measuring absorbance at 260 nm. One microgram of total RNA was used to prepare cDNA by reverse transcription using an Omniscript RT kit from QIAGEN (Mississauga, ON, Canada) and random primer from Invitrogen. The mRNAs encoding for Pgp (mdr1a), MRP2, and Oatp3 were measured by quantitative real-time PCR using Platinum SybrGreen PCR Supermix UDG from Invitrogen and appropriate primers on an ICycler thermocycler (BioRad, Mississauga, ON, Canada) with 30-s incubation at 94°C, 30-s incubation at 60°C, and 30-s incubation at 72°C. Table 1 shows the sequences of primers used for the quantification of mRNA for each transporter.
Nucleotide sequences of PCR primers Primers were designed based on published cDNA sequences with the help of the Jellyfish and BLASTIN 2.20.1 computer programs (Altschul et al., 1997). The resulting PCR products were sequenced on an ABI Prism 3100 analyzer (Applied Biosystems, Foster City, CA).
Immunohistochemistry Studies. Biopsies of intestines were fixed in buffered 4% paraformaldehyde for 24 h and embedded in paraffin in transversal orientation to allow exposure of the intestinal lumen on the slides. Slides were deparaffined and treated in 0.1 M EDTA, pH 8.0, under microwaves. Endogenous peroxidases were blocked in H2O2-methanol for 10 min. After blocking with protein blocking agent for 30 min, slides were incubated for 60 min with the primary antibody used for Western blotting. Immune complexes were revealed with the LSAB 2 System horseradish peroxidase kit from Dako (Mississauga, ON, Canada), following instructions provided by the manufacturer. Slides were counterstained using hematoxylin and mounted.
In Situ Hybridization Studies. In situ hybridization probes were prepared as follows. Primers for mdr1a and mrp2 were modified to include T7 sequence at the 5′ extremity of the sense primers and SP6 sequence at the 5′ extremity of the antisense primer. PCRs were performed in intestinal cDNA as follows: mdr1a-T7 sense primer with mdr1a antisense primer, thus creating a sense probe used as a negative control, and mdr1a sense primer, with mdr1a-SP6 antisense primer creating an antisense probe for the localization of mRNA. The same PCRs were performed with the mrp2 genes. Digoxigenin (DIG)-labeled RNA probes were obtained using DIG-labeled RNA polymerase kit (Roche Diagnostics, Laval, QC, Canada) with T7 or SP6 RNA polymerases and 1 μg of each PCR product. Slides were prepared as for the immunohistochemistry studies. RNA complexes were colored by incubation of the slides with the alkaline phosphatase substrates nitro blue tetrazolium and 5-bromo-4-chloro-3-indolylphosphate toluidine salt until the apparition of a brown coloration. Finally, slides were washed in phosphate-buffered saline, dried, and mounted for observation.
Pgp and MRP2 Activity in Everted Gut. Pgp function was evaluated by the in vitro transport of Rho-123 (Sigma) across rat everted gut sacs as described previously (Barthe et al., 1999; Veau et al., 2001). The ileum (where Pgp is more abundant) was removed and cut into 6-cm-long segments. These were everted, filled with 0.7 ml of rhodamine 123 solution, and ligated at each extremity. Secretion of Rho-123 from the serosal to the mucosal side was measured by sampling 200 μl of the external medium every 15 min up to 120 min. The rate of Rho-123 transport was expressed in picomoles excreted in the mucosal compartment per minute. To evaluate the residual transport of Rho-123 in this model, the Pgp inhibitor verapamil was added at concentrations of 300 μM (Barthe et al., 1999) to both mucosal and serosal sides after 90 min. Rho-123 was measured by spectrofluorometry (excitation, 485 nm; emission, 530 nm).
MRP2 activity was evaluated by the transport of 5-(and-6)-carboxy-2′,7′dichlorofluorescein (CDF) using also the everted gut sacs system. Everted jejunum (where MRP2 is most abundant) segments were filled with 0.7 ml of 100 μM 5-(and-6)-carboxy-2′,7′dichlorofluorescein diacetate (Molecular Probes, Eugene, OR) in buffer A. CDF was administered in its diacetylated form to facilitate the passive absorption into the cells. Cytoplasmic esterases rapidly digested 5-(and-6)-carboxy-2′,7′dichlorofluorescein di-acetate into CDF immediately after its diffusion into the cells (Zamek-Gliszczynski et al., 2003). Excretion of CDF from the serosal to the mucosal side by MRP2 was measured by sampling 200 μl of the external medium every 15 min up to 120 min. The MRP2 inhibitor probenecid was added at a concentration of 10 mM after 90 min (Zamek-Gliszczynski et al., 2003; Itagaki et al., 2005). CDF was measured by spectrofluorometry (excitation, 485 nm; emission, 530 nm).
Effects of Uremic Serum on Intestinal Drug Transporters. To determine whether uremic serum could affect the expression and function of intestinal drug transporters, two sets of experiments were conducted using normal rat enterocytes or Caco-2 cells. Rat enterocytes were used to study the effects of uremic serum on drug transporter protein expression, whereas Caco-2 cells were used to study the effects of uremic serum on their activity.
Incubation of Enterocytes with Uremic Serum. Normal rat enterocytes were harvested as described above. Instead of preparing a homogenate with these cells, they were seeded into 75-mm uncoated polystyrene flasks (Sarstedt, St. Leonard, QC, Canada) at a density of 12 × 106 cells/flask in culture medium containing 10% CTL or CRF rat serum. Viability was measured by the trypan blue exclusion method. After 48 h of culture, the cells were pelleted and homogenized in 0.9% NaCl, 1% SDS, and 75 μg was loaded for Western Blot analysis.
Rhodamine Transport on Caco-2 Cells. The effects of uremic serum on Pgp function were evaluated in Caco-2 cells using the transepithelial transport of Rho-123 across Caco-2 monolayers as reported previously (Barthe et al., 1999; Yumoto et al., 1999; Huang et al., 2000). Caco-2 cells were cultured on Transwell membranes for 21 to 28 days in DMEM (Invitrogen) containing 10% fetal bovine serum (Medicorp, Montreal, QC, Canada), 1% nonessential amino acids (Invitrogen), 1% l-glutamine (Invitrogen), and 1% Penstrep (a mixture of 20 000 U/mg penicillin and streptomycin). Twenty-four hours before the transport experiment, culture medium was replaced by culture medium containing 10% CTL or CRF rat serum. This medium was replaced 1 h before the experiment by everted gut buffer A containing 10% serum from CTL or CRF rats on the apical and basolateral sides of the Transwell membranes. Finally, Rho-123 was added to the apical chamber, and the transepithelial transport of Rho-123 was measured at 37°C for 150 min. After 90 min, the Pgp inhibitor cyclosporine A (Novartis, Dorval, QC, Canada) was added at a concentration of 0.06 mg/ml on both apical and basolateral sides. Rho-123 was measured by spectrofluorometry (excitation, 485 nm; emission, 530 nm).
Statistical Analysis. The results are expressed as mean ± S.E.M. Differences between groups were assessed by using an unpaired Student's t test or an ANOVA test. Significant ANOVA was followed by a Scheffe post hoc comparison of groups. The threshold of significance was p < 0.05.
Results
Biochemical Parameters and Body Weight in Control and CRF Rats.Table 2 presents the biochemical parameters and body weights of both groups of rats. Compared with control animals, CRF rats had higher levels of serum creatinine and urea and lower creatinine clearances (reduced by 78%, p < 0.001). There was no difference in body weight between control and CRF rats.
Characteristics of control and CRF rats Data are the mean ± S.E.M. Measurements were made at the time of sacrifice. Urinary collection was begun the day before. Blood urea, blood creatinine, and urine creatinine were determined with a Hitachi 717 autoanalyzer (Boehringer Mannheim Canada, Laval, QC, Canada).
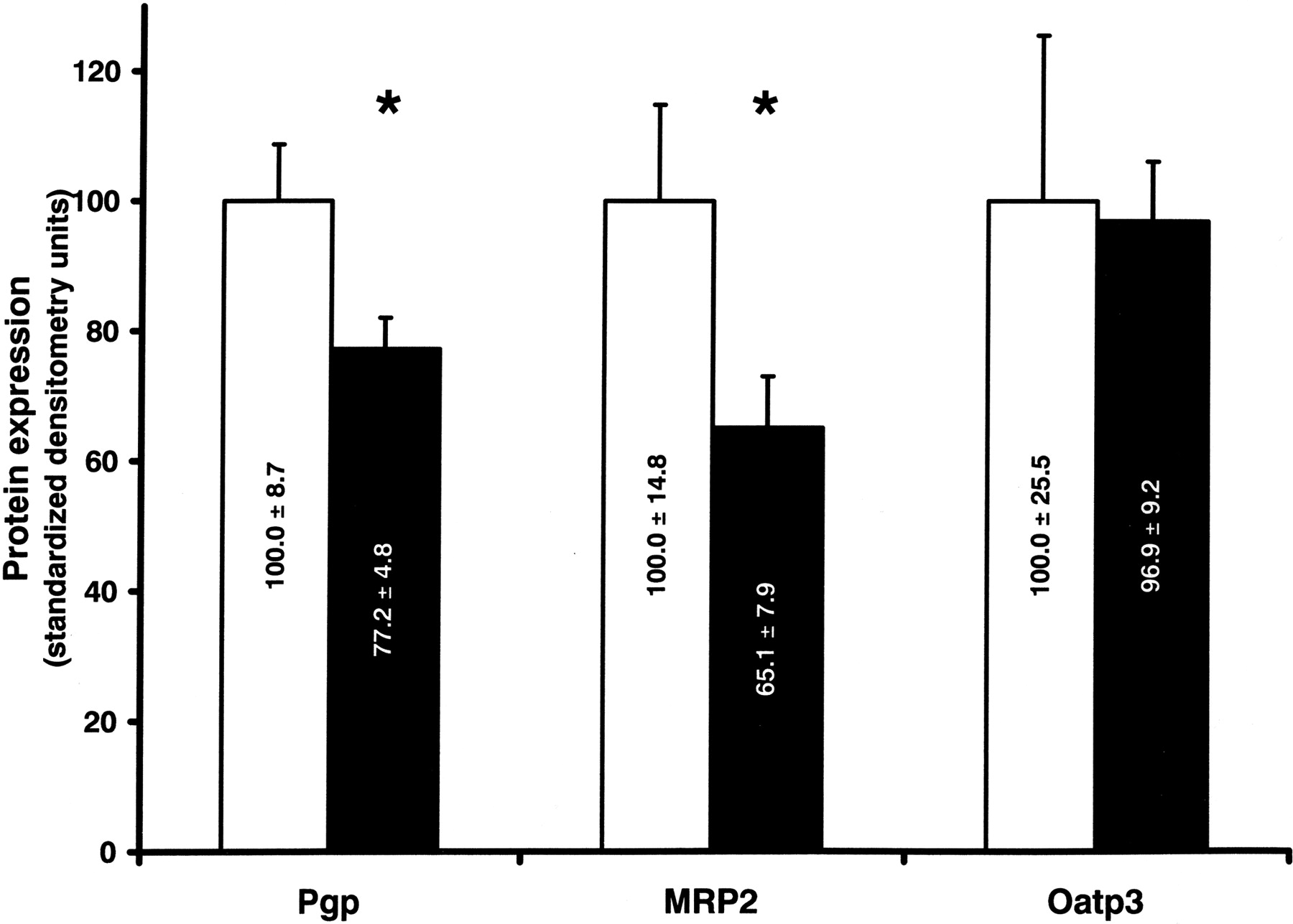
Intestinal Protein Expression of Pgp, MRP2, MRP3, Oatp2, Oatp3, and CYP3A2 in Control and CRF Rats. Compared with control animals, the levels of Pgp, MRP2, MRP3, and CYP3A2 in CRF rats were reduced by 65% (p < 0.001), 60% (p < 0.01), 35% (p < 0.0001), and 30% (p < 0.01) (Fig. 1), respectively, whereas the protein expression of Oatp2 and Oatp3 was not modified.
Immunohistochemistry. To determine whether CRF was associated with a modification in the distribution of Pgp and MRP2 within the intestinal mucosa (crypt versus villi) and within the enterocyte (membranous versus cytoplasmic), immunohistochemical analysis was performed. There was no difference in intestinal mucosa morphology between control and CRF rats (Fig. 2, A and B). Pgp was strongly detected in the surface epithelium in both groups of rats. The distribution of Pgp along the villi was identical in control and CRF rats (Fig. 2, C and D). Similar results were obtained for MRP2 (Fig. 2, E and F).
mRNAs Encoding Intestinal Pgp, MRP2, Oatp3, and CYP3A2 in Control and CRF Rats. There was no difference in mRNA levels of Pgp, MRP2, and Oatp3 between CRF and control rats. On the opposite, mRNA encoding for CYP3A2 was significantly reduced (25%, p < 0.05) in CRF rats (Fig. 3) compared with control animals, as shown previously (Leblond et al., 2002).
In Situ Hybridization. No modulation of the mRNA encoding for transporters was observed on eluted cells, consisting mainly of mature enterocytes (Fig. 3). This does not exclude a modification in the distribution of the mRNA more deeply in the intestinal epithelium. The expression pattern of Pgp and MRP2 mRNAs along the vertical axis of the intestinal villi was thus examined by in situ hybridization. As shown in Fig. 4, Pgp and MRP2 mRNAs were detected in both groups of animals, and the observed expression was mainly in enterocytes located at the crypt-villus junction and in the crypt. Control rats displayed no significant differences in localization compared with CRF animals (Fig. 4).
Pgp and MRP2 Activity in Everted Gut. As shown in Fig. 5A, Rho-123 efflux was reduced by 30% in CRF rat intestine (p = 0.003). For MRP2, CDF efflux was reduced by 23% in CRF rats (p = 0.002) (Fig. 5B). Although these substrates have been shown to be quite specific for these drug transporters, there is nevertheless an efflux that is not mediated by Pgp or MRP2. As shown in Fig. 5, A and B, this residual transport of rhodamine 123 and CDF (measured after the addition of inhibitors verapamil for Pgp and probenecid for MRP2) was identical in CRF and CTL rats, suggesting that CRF affects only the transport of these substrates mediated by Pgp and MRP2. However, because these inhibitors are not highly specific, CRF could still inhibit transport not mediated by these transporters.

Protein expression of intestinal transporters and cytochrome P450 3A2 in intestinal cell homogenate from control (white bars) and CRF (black bars) rats. Protein bands are expressed in densitometry units. The densitometry units measured for transporters and CYP3A2 were standardized by dividing them by the value obtained for villin. The standardized densitometry units of control rats were arbitrarily defined as 100%. Data are the mean ± S.E.M. of eight experiments in each group. *, p < 0.01 compared with control.

Immunolocalization of membrane intestinal transporters in control (C and E) and CRF (D and F) rat intestine. Jejunum biopsies were fixed in 4% phosphate-buffered paraformaldehyde. Tissues were either stained with hematoxylin-phloxin for histological examination (A and B) or incubated with specific antibodies for Pgp (C and D) or MRP2 (E and F). Immune complex were revealed by DAB, leaving a brown coloration on the extremities of the villi (see blue arrows). As shown, CRF does not modify the epithelium structure or the distribution of transporters (brown coloration), which are expressed all along the villi and are absent from crypts. Magnification, 200×.
Correlation of Pgp and MRP2 Protein Expression and Activity with Biochemical Parameters. All the CRF rats present a decrease in Pgp and MRP2 protein expression and activity. However, there was a difference in the magnitude of the decrease. We evaluated whether uremic markers were correlated with intestinal drug transporter reduction. We found significant correlations between the protein expression as well as activity of Pgp and MRP2 with creatinine clearance (Fig. 6). No correlation was found with serum urea (data not shown).
Protein Expression of Pgp and MRP2 in Enterocytes Incubated with Serum from Control or CRF Rats. To characterize the mechanism underlying the decrease in drug transporters, we next evaluated the effects of CRF serum on the expression of membrane transporters. After 48 h of incubation of normal enterocytes with serum (10%) from rats with CRF, drug transporter expression decreased compared with enterocytes incubated with serum from control animals (Fig. 7). The levels of Pgp and MRP2 were reduced by 23 and 35%, respectively (p < 0.01), whereas the levels of Oatp3 and villin were similar in the two groups of enterocytes.

mRNAs encoding intestinal drug transporters and CYP3A2 in intestinal cells from control (white bars) and CRF (black bars) rats were measured by quantitative real-time PCR. mRNAs are expressed in number of copies of the gene. The number of copies of control rats was arbitrarily defined as 100%. Data are the mean ± S.E.M. of eight experiments in each group. p < 0.05 compared with control rats.

In situ localization of mRNA coding for transporters in control (A and C) and CRF (B and D) rats. Jejunum biopsies were fixed in 4% phosphate-buffered paraformaldehyde. RNA probes were produced and labeled with digoxigenin with the DIG-labeled RNA polymerase kit from Roche Diagnostics. Hybridization was revealed with nitro blue tetrazolium and 5-bromo-4-chloro-3-indolyphosphate toluidine salt, which produced a dark deposit in the crypts (see blue arrows). As shown, CRF does not change the site of synthesis of Pgp (A and B) or MRP2 (C and D). Positive signal was detected only in crypt cells and was totally absent from villi mature cells. Magnification, 200×.
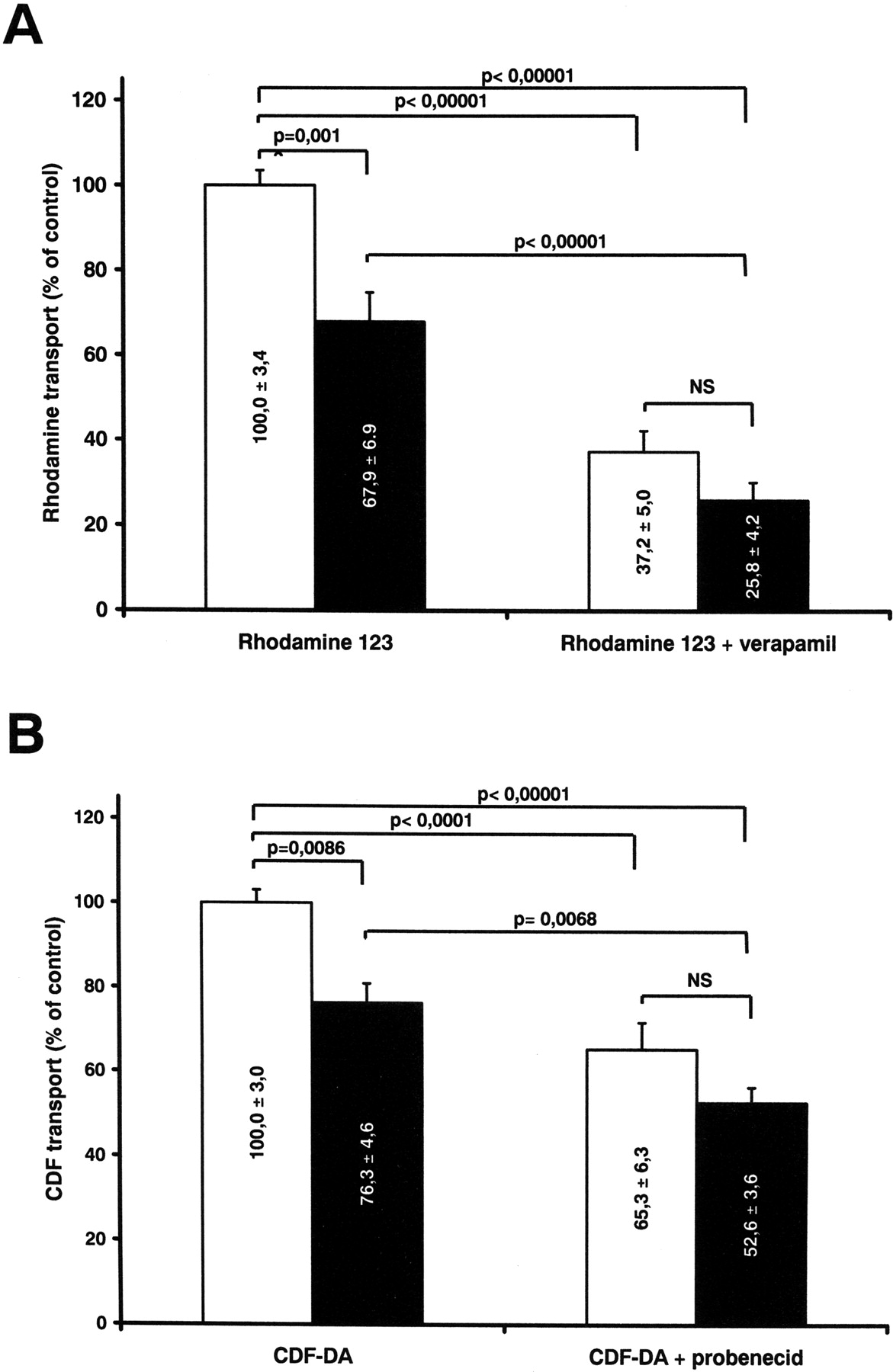
In vitro transport of rhodamine 123 and CDF in everted gut sacs. A, transport of rhodamine 123 in control (white bars) and CRF (black bars) everted gut sacs in the absence and presence of inhibitor verapamil. Control Rho-123 transport was 1092.8 ± 165.4 pmol/min and was defined as 100%. B, CDF transport in control (white bars) and CRF (black bars) in the absence or presence of the MRP2 inhibitor probenecid. Control CDF transport was 122.9 ± 7.9 pmol/min and was defined as 100%. Data are the mean ± S.E.M.
Pgp Activity in Caco-2 Cells Incubated with Serum from Control or CRF Rats. The effect of uremic serum on Pgp function was evaluated in Caco-2 cells (Fig. 8) using the transepithelial transport of Rho-123 across Caco-2 monolayers by measuring its apparent permeability (Papp). Papp was determined using the equation Papp = (ΔQ/Δt)/(AC0), where (ΔQ/Δt) is the linear rate of appearance of Rho-123, A is the area of the Transwell membrane, and C0 is the initial concentration of Rho-123 (Yamashita et al., 2002). We found a significant increase in the Papp of rhodamine from the apical to the basolateral side of the Transwell membranes, arguing in favor of a decrease in Pgp activity.
Discussion
Several studies have shown that the bioavailability of many drugs (such as propranolol, dihydrocodeine, erythromycin, and tacrolimus) is increased in patients with CRF (Talbert, 1994; Matzke and Frye, 1997; Okabe et al., 2000). This suggests that the major determinants of bioavailability, i.e., the first pass metabolism and/or the intestinal extrusion of drugs, are decreased in CRF. We have already shown that intestinal P450 is reduced in CRF rats (Leblond et al., 2002). The purpose of the present study was to demonstrate that the intestinal extrusion of drugs is also affected by CRF.

A, correlation between Pgp protein expression and creatinine clearance of control and CRF rats (R2 = 0.210). B, correlation between MRP2 protein expression and creatinine clearance of control and CRF rats (R2 = 0.246). C, correlation between Pgp transport activities and creatinine clearance of control and CRF rats (R2 = 0.277). D, correlation between MRP2 transport activities and creatinine clearance of control and CRF rats (R2 = 0.424). p < 0.05 for each correlation.

Level of Pgp, MRP2, and Oatp3 protein expression in enterocytes incubated with 10% serum from control (white bars) and CRF (black bars) rats for 48 h. Protein bands are expressed in standardized densitometry units (see Fig. 1). Data are the mean ± S.E.M. of three experiments. *, p < 0.05 compared with enterocytes incubated with serum from control rats.
Our results demonstrate, for the first time, that the protein expression of some major efflux transporters (Pgp, MRP2, and MRP3) is reduced in CRF, whereas the protein expression of the import transporters Oatp2 and Oatp3 and of villin (a structural marker of enterocytes) remains unchanged. Veau et al. (2001) have demonstrated previously a reduction in the activity of Pgp in CRF rats. However, they did not detect any difference in protein expression of intestinal Pgp in CRF rats. This discrepancy could potentially be explained by differences in the technique used for intestinal cell isolation. Indeed, the scraping of intestinal epithelium, as performed by Veau et al., favors the recovery of all types of cell constituting the intestinal mucosa. The elution technique (Weiser, 1973) used in the present study favors the preferential recovery of mature enterocytes from the top of the villi.

Papp of rhodamine 123 from the apical to the basolateral side of Caco-2 cell monolayers incubated with CTL (white bars) or CRF (black bars) rat serum, before and after the addition of the inhibitor cyclosporine A. Papp was determined using the equation Papp = (ΔQ/Δt)/(AC0), where (ΔQ/Δt) is the linear rate of appearance of rhodamine, A is the area of the Transwell membrane, and C0 is the initial concentration of rhodamine 123 (Yamashita et al., 2002). Papp of cells incubated with CTL serum was arbitrarily defined as 100%. Data are the mean ± S.E.M. of six experiments. *, p < 0.05.
The difference in Pgp and MRP2 protein expression does not seem to be a consequence of decreased mRNA expression, as demonstrated by our quantitative PCR results. To further rule out a decrease in gene expression, which could have been missed by the use of an epithelial cell subpopulation, we studied the expression profile of the mRNAs of both Pgp and MRP2 by in situ hybridization. Because the protein distribution is not uniform along the vertical axis of the villus, it is possible that the use of mature enterocytes precludes the demonstration of a difference in the synthesis of the transporter. Despite the semiquantitative nature of the in situ hybridization technique, no evidence could be obtained, demonstrating a difference in the mRNA transcription in CRF intestinal epithelium. As shown in Fig. 4, the mRNAs for Pgp and MRP2 are mostly expressed in crypt cells in control and CRF intestines, and there was no major difference in localization between CRF and control rats.
Thus, it seems that the down-regulation of Pgp and MRP2 in CRF depends on post-translational mechanisms, such as a faulted maturation and folding of the protein, a decrease in its migration to the membrane, or an increase in its internalization as well as its degradation. However, none of these mechanisms has been shown to be involved in CRF. The mechanism by which Pgp and MRP2 protein expression is decreased remains to be determined, and further studies are ongoing.
The repercussions of CRF on intestinal extrusion of drugs are important since we observed a significant reduction in Pgp and MRP2 activity. We have demonstrated, using the everted gut technique, that Pgp and MRP2 activity were reduced by 32 and 24% in CRF rat intestine using specific substrates Rho-123 and CDF (Fig. 5, A and B). No significant difference in the residual transport of the substrates was measured between CRF and control rats, suggesting that CRF mostly impedes the efflux of drugs mediated by Pgp and MRP2. Our results confirm those published by Veau et al. (2001), who demonstrated a 33% decrease in Pgp activity using Rho-123 as a Pgp substrate across CRF rat intestine. The reduction in drug export that we observed in CRF rat intestines could explain the increase in bioavailability of certain drugs described in CRF patients. On the other hand, we have shown a reduction in MRP3 expression, which is an efflux transporter located at the basolateral membrane. However, its role in the bioavailability of drugs remains to be defined (Chan et al., 2004; Shoji et al., 2004; Zimmermann et al., 2005). Further studies are needed to evaluate the effects of CRF on the other basolateral membrane transporters.
A number of studies indicate that intestinal function could be altered in CRF. Several enzymatic reactions carried out by the intestinal mucosa have been shown to be decreased in rats with CRF. For instance, sucrase and maltase activities, as well as xanthine oxidase, were significantly reduced, whereas the activities of other dipeptidases were unchanged (Grimmel et al., 1977). More recently, we have also demonstrated that intestinal cytochrome P450 is reduced in CRF (Leblond et al., 2002). The results of the present study strongly support the hypothesis that CRF is associated with perturbations in intestinal function.
We have tried to determine whether a mediator present in uremic serum could be responsible for the decrease in protein expression and activity of intestinal drug transporters in CRF. First, we have demonstrated a correlation between the clearance of creatinine and the protein expression and the activity of Pgp and MRP2. This suggests that the more severe the renal failure, the more important is the decrease in protein expression and activity of the transporters. This argues in favor of the presence of a serum mediator responsible for these decreases. Second, we have demonstrated that the incubation of cultured mature enterocytes with CRF serum decreases by 25 and 35% protein expression of Pgp and MRP2. We have also demonstrated that uremic serum increases significantly the Papp of Rho-123 from the apical to the basolateral side of a Caco-2 cell monolayer, which argues in favor of a decrease in the transport activity of Pgp, allowing Rho-123 to migrate more easily toward the basolateral side. The addition of the Pgp inhibitor cyclosporine A on the basolateral and apical sides causes an important increase in the Papp of rhodamine, which demonstrates that Rho-123 transport is Pgp-dependent. Once again, this strongly suggests that a factor present in uremic serum has a modulatory effect on Pgp and MRP2 protein expression and activity, similar to what we showed for the P450 (Leblond et al., 2002).
Several endogenous molecules accumulate in CRF, and some of them have been recognized to have an effect on drug transporters either in the liver or in the kidney. Recently, Sun et al. (2004) have shown that 3-carboxy-4-methyl-5-propyl-2-furan-propanoic acid, a uremic toxin, decreases the activity and expression of Oatp2 in the liver. Another study by Shibata et al. (2000) has demonstrated that Pgp-mediated cyclosporine transport is reduced in Caco-2 cells in the presence of uremic toxins 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid and indoxyl sulfate. Whether the presence of these toxins could explain the results in our model remains to be defined.
Other potential uremic mediators are cytokines. Several studies have demonstrated that CRF is associated with a chronic activation of inflammatory response (Oettinger et al., 1994; Ziesche et al., 1994; Akahoshi et al., 1995; Higuchi et al., 1997; Stenvinkel et al., 1999; Yao et al., 2004). Cytokines are able to down-regulate intestinal and liver drug transporters in vitro and in vivo (Sukhai et al., 2001; McRae et al., 2003). This supports the hypothesis that the mediators contained in the plasma of CRF rats that down-regulate the intestinal transporters could be some specific cytokines that are increased in CRF. Furthermore, interleukin-1β has been demonstrated to decrease Pgp protein expression and activity via a post-translational mechanism (Sukhai et al., 2001) but to decrease CYP3A2 expression via a decrease in mRNA (Abdel-Razzak et al., 1993; Abdel-Razzak et al., 1995). This could also explain why we demonstrated a decrease in CYP3A2 mRNA expression, whereas the transporter mRNA remained unchanged.
In conclusion, CRF induces a decrease in the activity of the two main intestinal drug efflux transporters, Pgp and MRP2, secondary to a marked reduction in their protein expression, whereas influx transporters Oatp2 and Oatp3 are not affected. The down-regulation of Pgp and MRP2 induced by CRF rat serum was similar to that produced by CRF in in vivo models, suggesting that uremic mediator(s) are responsible for the reduction in intestinal drug extrusion in vivo.
Acknowledgments
We thank François Boudreau (Department of Anatomy and Cellular Biology, Université de Sherbrooke, Sherbrooke, QC, Canada) for generous help in the realization of in situ hybridization.
Footnotes
-
This work was supported by the Canadian Institute for Health Research and by the Fonds de la Recherche en Santé du Québec. V.P. and J.N. are scholars of the Fonds de la Recherche en Santé du Québec.
-
This work was presented in the Master's thesis of J.N. at the Universitéde Montréal, Département de Pharmacologie in December 2005.
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.106.112631.
-
ABBREVIATIONS: CRF, chronic renal failure; Pgp, P-glycoprotein; MRP, multidrug resistance-related protein; Oatp, organic anion-transporting polypeptide; Rho-123, rhodamine 123; DIG, digoxigenin; PCR, polymerase chain reaction; CDF, 5-(and-6)-carboxy-2′,7′dichlorofluorescein; CTL, control; Papp, apparent permeability; P450, cytochrome P450.
- Received August 17, 2006.
- Accepted November 27, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}