


Abstract
The low-affinity sodium glucose cotransporter (SGLT2) plays an important role in renal glucose reabsorption and is a remarkable transporter as a molecular target for the treatment of diabetes. We have discovered remogliflozin etabonate, which is a novel category of selective SGLT2 inhibitors. Remogliflozin etabonate is a prodrug based on benzylpyrazole glucoside and is metabolized to its active form, remogliflozin, in the body. We identified remogliflozin to be a potent and highly selective SGLT2 inhibitor by examining COS-7 cells transiently expressing either high-affinity sodium glucose cotransporter (SGLT1) or SGLT2. Orally administered remogliflozin etabonate increased urinary glucose excretion in a dose-dependent manner in both mice and rats. By increasing urinary glucose excretion, remogliflozin etabonate inhibited the increase in plasma glucose after glucose loading without stimulating insulin secretion in normal rats. Remogliflozin etabonate also showed antihyperglycemic effects in both streptozotocin-induced diabetic rats in oral glucose tolerance and in db/db mice in the fed condition. Chronic treatment with remogliflozin etabonate reduced the levels of fasting plasma glucose and glycated hemoglobin, and it ameliorated glucosuria in db/db mice. In high-fat diet-fed Goto-Kakizaki rats, remogliflozin etabonate improved hyperglycemia, hyperinsulinemia, hypertriglyceridemia, and insulin resistance. This study demonstrates that treatment with remogliflozin etabonate exhibits antidiabetic efficacy in several rodent models and suggests that remogliflozin etabonate may be a new and useful drug for the treatment of diabetes.
Type 2 diabetes is characterized by insulin resistance and impaired glucose-stimulated insulin secretion by the pancreatic β-cells (Dwarakanathan, 2006), and it has increased in incidence with changes in lifestyles that lead to weight gain and obesity (Walker et al., 2007). The basic management of type 2 diabetes is achieved by lifestyle interventions such as dietary changes and exercise (Nathan et al., 2006). When a restricted lifestyle is not maintained, chronic hyperglycemia leads to progressive impairment of insulin secretion and to insulin resistance of peripheral tissues, a phenomenon often referred to as glucose toxicity (Rossetti et al., 1990; Leahy et al., 1992), which further worsens the blood glucose level. In recent studies, instead of lifestyle intervention, stimulating glucose excretion in urine has been proposed as a new approach for the treatment of type 2 diabetes (Isaji, 2007).
In the kidney tubules, the bulk of the filtrated glucose is mainly reabsorbed by the low-affinity sodium glucose cotransporter (SGLT2) (Wells et al., 1992; Kanai et al., 1994), and the residual glucose is reabsorbed by the high-affinity sodium glucose cotransporter (SGLT1) (Pajor and Wright, 1992; Wright, 2001). Mutations in the SGLT1 gene cause glucose/galactose malabsorption, and patients with this disorder suffer from significant gastrointestinal dysfunction, but they show only mild renal glucosuria (Turk et al., 1991; Martín et al., 1996). On the other hand, mutations in the SGLT2 gene result in renal glucosuria (van den Heuvel et al., 2002; Calado et al., 2004). These findings indicate that SGLT2 plays a dominant role in the control of glucose transport in the kidney. In addition, our previous results obtained with a selective SGLT2 inhibitor confirmed that SGLT2 plays an important role in renal glucose reabsorption and that SGLT2 is an appropriate molecular target for the treatment of diabetes (Katsuno et al., 2007).
To date, a number of SGLT inhibitors have been evaluated. The first one to be evaluated was phlorizin, which is a natural product and dietary constituent found in a number of fruit trees; and it has been used as a pharmaceutical and tool for physiology research (Ehrenkranz et al., 2005). Phlorizin increased urinary glucose excretion (UGE), lowered the plasma glucose without increasing insulin secretion, and normalized insulin sensitivity in animal models of diabetes (Starke et al., 1985; Rossetti et al., 1987; Harmon et al., 2001). However, phlorizin was not developed as a drug for the treatment of diabetes because of its nonselectivity (Oku et al., 1999; Katsuno et al., 2007) and low bioavailability. Subsequently, an orally absorbable SGLT inhibitor, T-1095, was developed to overcome this latter disadvantage of phlorizin (Oku et al., 1999). Being a phlorizin derivative, the active form of T-1095 (T-1095A) inhibited both SGLT1 and SGLT2 (Oku et al., 1999). There is no consensus as to whether SGLT1 inhibition is needed in addition to SGLT2 inhibition. Several researchers reported on other SGLT inhibitors (Kees et al., 1996; Link and Sorensen, 2000; Ohsumi et al., 2003; Zhang et al., 2005, 2006; Isaji, 2007). We have developed a selective SGLT2 inhibitor, sergliflozin, which is a prodrug based on benzylphenol glucoside (Katsuno et al., 2007). In this report, we describe the properties and potency of another new selective SGLT2 inhibitor, remogliflozin etabonate, for the treatment of type 2 diabetes, the structural scaffold of which is different from that of phlorizin, T-1095, or sergliflozin.
Materials and Methods
Chemicals. Remogliflozin etabonate (4-[(4-isopropoxyphenyl)-methyl]-1-isopropyl-5-methyl-1H-pyrazol-3-yl 6-O-ethoxycarbonyl-β-d-glucopyranoside) and remogliflozin (4-[(4-isopropoxyphenyl)-methyl]-1-isopropyl-5-methyl-1H-pyrazol-3-yl β-d-glucopyranoside) were synthesized by Kissei Pharmaceutical Co., Ltd. (Azumino, Nagano, Japan). Phlorizin dihydrate and methyl-α-d-glucopyranoside (AMG) were purchased from Sigma-Aldrich (St. Louis, MO). Methyl-α-d-[U-14C]glucopyranoside was obtained from GE Healthcare BioSciences (Little Chalfont, Buckinghamshire, UK).
Inhibitory Effects on Human and Rat SGLTs. Human SGLT1 and SGLT2 expression plasmids were constructed as described previously (Katsuno et al., 2007). Rat SGLT1 and SGLT2 genes were amplified by reverse transcriptase-polymerase chain reaction from the total RNA of the small intestine and kidney (Origene, Rockville, MD), respectively. The 2-kb cDNA of rat SGLT1 (accession no. D16101) or the 2-kb cDNA of rat SGLT2 (accession no. U29881) was ligated into the pCMV-Script vector (Stratagene, La Jolla, CA). For the AMG uptake experiment, COS-7 cells (RIKEN Cell Bank, Tsukuba, Japan) were seeded into a 96-well collagen-coated culture plate at a density of 5 × 104 cells/well in Dulbecco's modified Eagle's medium (Sigma-Aldrich) containing 10% fetal bovine serum. After 4 h, each plasmid was transfected into COS-7 cells using Lipofectamine 2000 (Invitrogen, Carlsbad, CA), and the cells were used 2 days after transfection. The AMG uptake experiment was performed as described previously (Katsuno et al., 2007). In this experiment, 0.3 and 1 mM AMG concentrations in the uptake buffer were used to calculate Ki values.
Animals. Male Sprague-Dawley rats were purchased from Japan SLC, Inc. (Hamamatsu, Japan), and male Wistar rats were obtained from Charles River Japan, Inc. (Yokohama, Japan). Female C57BL/6J mice, male db/+m (BKS.Cg-m +/+ Leprdb/Jcl) mice, male db/db (BKS.Cg-+ Leprdb/+ Leprdb/Jcl) mice, and male Goto-Kakizaki (GK) rats were obtained from CLEA Japan Inc. (Tokyo, Japan). These rodents were housed under a 12-h/12-h light cycle (light on 8:00 AM–8:00 PM) under controlled conditions (room temperature, 20–26°C; humidity, 35–65%) and fed a laboratory chow diet (CE-2 pellets; CLEA Japan, Inc.) and water ad libitum. All animal experiments were performed in accordance with the guidelines approved by the Laboratory Animal Committee of Kissei Pharmaceutical Co., Ltd.
Urinary Glucose Excretion in Mice and Rats. Remogliflozin etabonate was suspended (5 ml/kg) in 0.5% sodium carboxymethylcellulose (CMC) and orally administered to male Sprague-Dawley rats (7 weeks of age) and female C57BL/6J mice (9 weeks of age). Urine was collected for 24 h after administration while the animals were kept in metabolic cages. After the urine volume had been measured, the glucose concentration in the urine was determined using a Glucose B-test Wako (Wako Pure Chemicals, Osaka, Japan), and then urinary glucose excretion was calculated.
Oral Glucose Tolerance Test in Normal Rats. Male Sprague-Dawley rats (7 weeks of age) were fasted for 16 h. Remogliflozin etabonate (5 ml/kg, in 0.5% CMC) and glucose solution (400 g/l, 5 ml/kg) were administered orally. Blood was obtained in heparinized and aprotinin-treated tubes from a tail vein at each sampling point. Plasma glucose concentrations were determined using a Glucose B-test Wako (Wako Pure Chemicals). Plasma insulin was determined using an enzyme-linked immunosorbent assay kit (Morinaga Institute of Biological Science, Inc., Yokohama, Japan).
Oral Glucose Tolerance Test in Streptozotocin-Induced Diabetic Rats. Male Wistar rats (8 weeks of age) received i.v. injections of streptozotocin [(STZ) 45 mg/kg; Wako Pure Chemical] to make diabetic model rats. After 10 days, the oral glucose tolerance test was performed as described above. Plasma glucose concentrations were determined using a Glucose CII-test Wako (Wako Pure Chemicals). The area under the curve (AUC)0–2 h for plasma glucose was calculated from the plasma glucose concentration during the oral glucose tolerance test.
Single Administration in db/db Mice. Remogliflozin etabonate (5 ml/kg, in 0.5% CMC) was orally administered to male db/db mice (10 weeks of age) in the fed condition. Blood was obtained from a tail vein at each sampling point. Blood glucose concentrations were measured with a glucose analyzer, ANTSENSE II (Bayer Medical Ltd., Tokyo, Japan). The AUC0–6 h for blood glucose was calculated from the blood glucose concentration.
Chronic Administration in db/db Mice. Remogliflozin etabonate (5 ml/kg, in 0.1% methylcellulose) was orally administered to male db/db mice (8 weeks of age) twice a day for 6 weeks. Body weight and food intake were measured every 2 weeks. These mice were fasted for 16 h every 2 weeks (in this regard, the second administration was not performed before fasting), and blood was obtained from a tail vein. Fasting plasma glucose (FPG), glycated hemoglobin (GHb), fasting plasma insulin, and fasting plasma triglyceride were monitored. At the end of the administration period, the mice were kept in metabolic cages to collect urine for 24 h. The plasma and urinary concentrations of glucose and the plasma insulin were determined as described above. Plasma triglyceride was determined using a Triglyceride E-test Wako (Wako Pure Chemicals), and glycated hemoglobin was determined by HLC-723GHbV (Tosoh, Tokyo, Japan).
Chronic Administration in High-Fat Diet-Fed GK Rats. Male GK rats (6 weeks of age) were fed either a normal diet (CE-2) or a high-fat one (QuickFat, 30% fat kcal; CLEA Japan Inc.) for 4 weeks. After allocation into groups, the latter were continuously fed the high-fat diet or the high-fat diet containing 0.01, 0.03, or 0.1% remogliflozin etabonate for 8 weeks. Wistar rats and GK rats were fed the normal diet (CE-2) throughout the experiment. The dose of remogliflozin etabonate consumed was calculated from the body weight and food intake every week. Blood was obtained from a tail vein of the rats in the fed condition, and plasma glucose, plasma insulin, plasma triglyceride, and GHb were monitored every 4 weeks. These rats were fasted for 16 h every 4 weeks, and the FPG and fasting plasma insulin were monitored. All parameters were determined as described above. The homeostasis model assessment ratio (HOMA-R) was calculated from the following formula: fasting plasma insulin (μU/ml) × FPG (mg/dl)/405 (Matthews et al., 1985). After 8 weeks of administration, the oral glucose tolerance test (1 g/kg at a volume of 5 ml/kg) was performed.
Statistical Analysis. Data were presented as means ± S.E.M. for each group. Statistical analysis was performed by using SAS Systems version 8.2 (SAS Institute, Cary, NC). Statistical significance was determined with univariate repeated-measures analysis as a split-plot design and multiple comparisons by each time period, t test, or Dunnett's test as appropriate.
Results
Chemical Structure of Remogliflozin Etabonate. The structures of remogliflozin etabonate and remogliflozin are shown in Fig. 1. Remogliflozin etabonate is metabolized to its active form, remogliflozin, the structural scaffold of which is based on benzylpyrazole glucoside. In view of its structure, remogliflozin etabonate belongs to a new category of SGLT2 inhibitors, because its skeleton differs from that of sergliflozin, phlorizin, or T-1095.
Inhibitory Effects on Human and Rat SGLTs. The inhibitory effects of remogliflozin etabonate, remogliflozin, and phlorizin on AMG uptake into COS-7 cells transiently transfected with human or rat SGLT expression plasmid, given in term of Ki, are shown in Table 1. For human SGLT2, the inhibitory effect of remogliflozin was approximately three times greater than that of phlorizin, but for SGLT1, it was only one twentieth of that of phlorizin. The ratio of selectivity (Ki value of human SGLT1/Ki value of human SGLT2) of remogliflozin and phlorizin was 365 and 7, respectively. Toward rat SGLT2, the inhibitory effect of remogliflozin was two times greater than that of phlorizin, but toward SGLT1, it was one seventh of that of phlorizin. The ratio of selectivity (Ki value of rat SGLT1/Ki value of rat SGLT2) of remogliflozin and phlorizin was 38 and approximately 3, respectively. These data suggest that remogliflozin, the active form of remogliflozin etabonate, is a potent and selective SGLT2 inhibitor.
Ki values of remogliflozin etabonate, remogliflozin, and phlorizin for human and rat SGLTs Data are presented as means ± S.E.M. from four experiments. SGLT1 or SGLT2 expression plasmid was transfected into COS-7 cells. An AMG uptake experiment was performed using transiently transfected cells, and the Ki values for each SGLT were calculated.

Chemical structures of remogliflozin etabonate (A), remogliflozin (B), sergliflozin (C), phlorizin (D), and T-1095 (E). Remogliflozin etabonate is metabolized to remogliflozin, its active form.
Effects on Urinary Glucose Excretion in Mice and Rats. To assess whether administration of remogliflozin etabonate increased urinary glucose excretion by inhibiting renal glucose reabsorption, we orally administered remogliflozin etabonate to normal mice and rats. In these animals, urinary glucose excretion was increased by the oral administration of remogliflozin etabonate in a dose-dependent manner (Fig. 2). Good linearity between dose and effects was observed. Thus, it was confirmed that remogliflozin etabonate is a selective SGLT2 inhibitor, and that oral administration of it increased urinary glucose excretion in a dose-dependent manner.
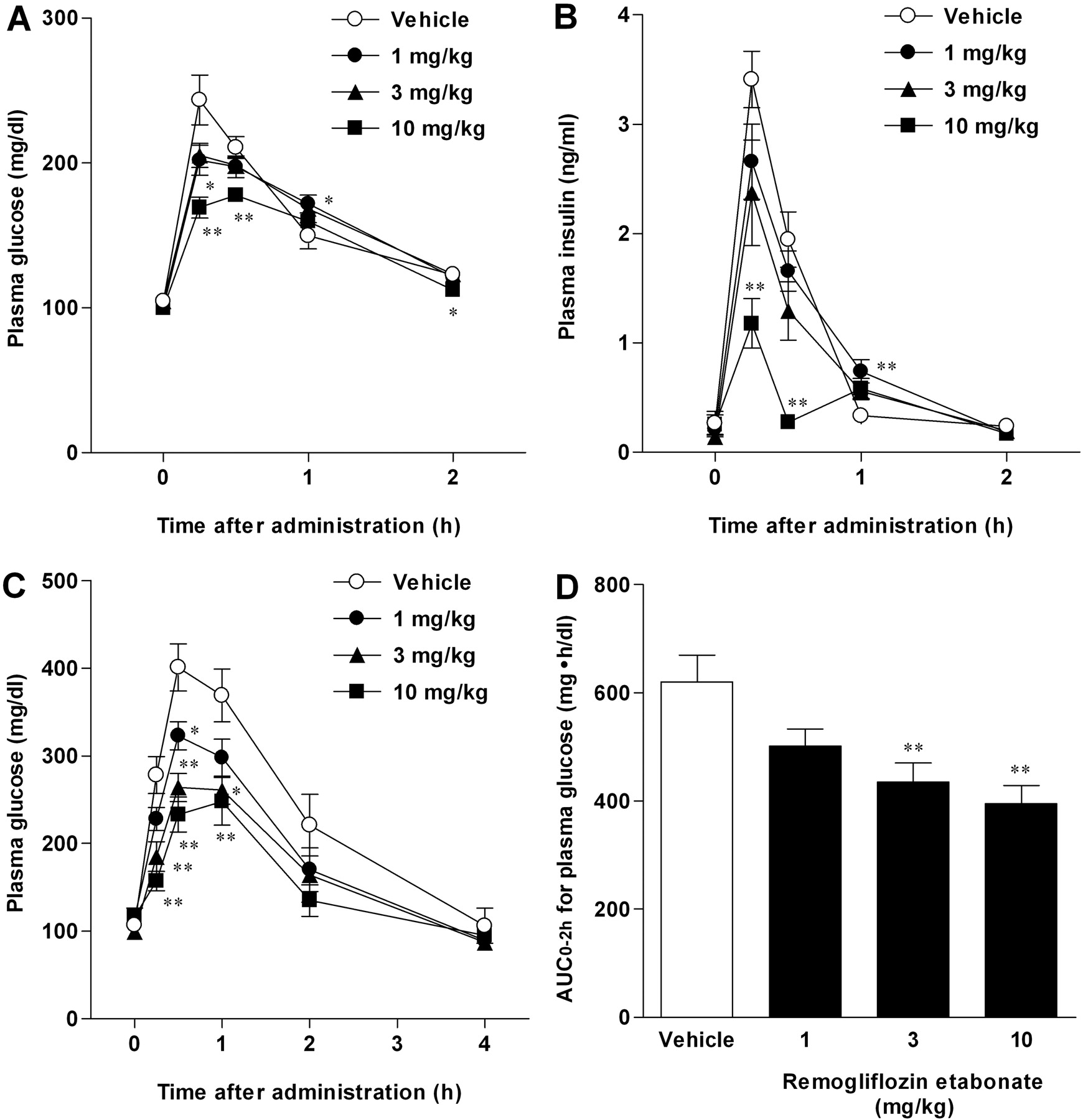
Antihyperglycemic Effects in the Oral Glucose Tolerance Test. Because remogliflozin etabonate increased urinary glucose excretion, we examined the antihyperglycemic effect of a single administration of remogliflozin etabonate in the oral glucose tolerance test. In normal rats, remogliflozin etabonate dose-dependently inhibited the increase in plasma glucose after glucose loading (Fig. 3A). At the same time, the plasma insulin was decreased by remogliflozin etabonate, corresponding to the reduction in the plasma glucose level (Fig. 3B). Thus, remogliflozin etabonate reduced the plasma glucose without stimulating insulin secretion. We next examined the antihyperglycemic effects in STZ-induced diabetic rats. In diabetic model rats, remogliflozin etabonate significantly inhibited the increase in plasma glucose after glucose loading in a dose-dependent manner (Fig. 3C), and the antihyperglycemic effects were more effective in the STZ-induced diabetic than in the normal rats (Fig. 3, A and C). In addition, remogliflozin etabonate reduced the AUC0–2 h for plasma glucose during the oral glucose tolerance test in a dose-dependent manner (Fig. 3D). The antihyperglycemic effects of remogliflozin etabonate were markedly enhanced in the diabetic state.
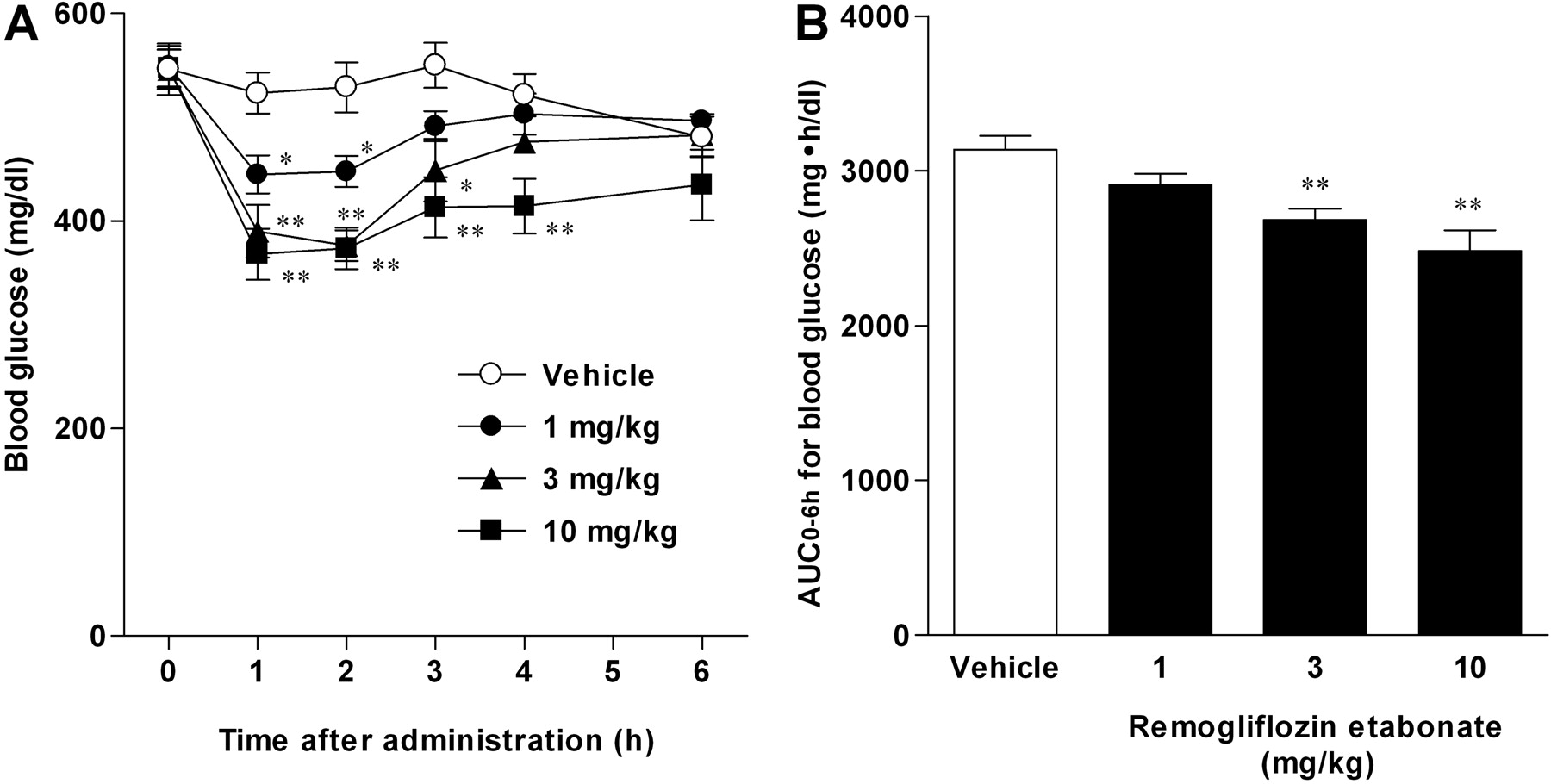
Antihyperglycemic Effects of Single Administration in db/db Mice. Because remogliflozin etabonate showed antihyperglycemic effects in STZ-induced diabetic rats, we next examined the effect in db/db mice with hyperinsulinemia and obesity. Remogliflozin etabonate decreased the blood glucose at the dose of 1–10 mg/kg (Fig. 4A), and it reduced the AUC0–6 h for blood glucose in a dose-dependent manner (Fig. 4B). Thus, remogliflozin etabonate showed an antihyperglycemic effect in diabetic mice with insulin resistance.

Oral administration of remogliflozin etabonate increased UGE in mice (A) and rats (B). Remogliflozin etabonate was orally administered to mice and rats. Urine was collected for 24 h after administration of remogliflozin etabonate, and UGE was determined. Data are presented as means ± S.E.M. (n = 4 for mice and n = 5 for rats). *, p < 0.05 and **, p < 0.01 versus vehicle group.
Antidiabetic Effects of Chronic Administration in db/db Mice. To examine the effects of chronic administration of remogliflozin etabonate, we divided db/db mice at the age of 8 weeks into three groups. Vehicle or remogliflozin etabonate (10 or 30 mg/kg) was orally administered twice daily for 6 weeks. The chronic effects of remogliflozin etabonate on physiological and metabolic parameters in db/db mice are shown in Table 2. Hyperglycemia was markedly advanced in the vehicle-treated db/db mice over the 6-week period. Remogliflozin etabonate significantly reduced both the FPG and GHb levels in a dose-dependent manner. In the vehicle-treated db/db mice, hyperinsulinemia was observed at the beginning of the experiment, but the plasma insulin level was decreased with the progression of diabetes. Remogliflozin etabonate inhibited the decrease in this level in these mice. Corresponding to reduction in plasma insulin, vehicle-treated db/db mice did not gain weight; and weight loss was observed after 6 weeks. At the same time, body weight gain was observed at a high dose of remogliflozin etabonate. However, remogliflozin etabonate did not affect the food intake throughout the treatment. In addition, hypertriglyceridemia was also observed with the progression of diabetes, and remogliflozin etabonate suppressed the development of it. After 6 weeks of administration, remogliflozin etabonate reduced both urine volume (data not shown) and urinary glucose excretion with ameliorated the hyperglycemia. These data show that remogliflozin etabonate was effective in improving disturbed glucose metabolism in experimental type 2 diabetes.
Chronic effects of remogliflozin etabonate on physiological and metabolic parameters in db/db mice Data are presented as means ± S.E.M. (n = 6 for db/+m mice, n = 8 for db/db mice). Remogliflozin etabonate was orally administered to db/db mice twice daily for 6 weeks. These mice were fasted for 16 h every 2 weeks, and FPG, GHb, fasting plasma insulin, and fasting plasma triglyceride were monitored. After 6 weeks of administration, these mice were kept in metabolic cages to collect urine for 24 h, and UGE was then determined.
Antidiabetic Effects of Chronic Administration in High-Fat Diet-Fed GK Rats. GK rats were fed either a normal or high-fat diet for 4 weeks. Thereafter, hyperglycemia, hyperinsulinemia, hypertriglyceridemia, and insulin resistance were observed in high-fat diet-fed GK rats (Table 3). To examine the chronic effects in this model, the high-fat diet containing 0.01, 0.03, or 0.1% remogliflozin etabonate was given to these rats for 8 weeks. The chronic effects of remogliflozin etabonate on physiological and metabolic parameters in these rats are shown in Table 3. The calculated doses of remogliflozin etabonate consumed were 5.9 (for the 0.01% drug group), 18.7 (0.03% group), and 68.4 (0.1% group) mg/kg/day. Weight loss was observed at a high dose of remogliflozin etabonate. However, remogliflozin etabonate had a tendency to increase food intake throughout the treatment (data not shown). Remogliflozin etabonate reduced the levels of plasma glucose, plasma insulin, and GHb in a dose-dependent manner, and it suppressed the development of hypertriglyceridemia. In the high-fat diet-fed GK rats, the FPG and fasting plasma insulin levels were elevated, and a higher value of HOMA-R was observed because of the insulin resistance. Remogliflozin etabonate reduced fasting plasma levels of glucose and insulin as well as the value of HOMA-R. After 8 weeks of administration, the oral glucose tolerance test (1g/kg) was performed. The values for the plasma glucose, plasma insulin, and the AUCs for these parameters are shown in Fig. 5. In the high-fat diet-fed GK rats, these values were increased compared with those of normal diet-fed GK rats. Chronic treatment with remogliflozin etabonate reduced the AUCs for plasma glucose and plasma insulin. These data show that remogliflozin etabonate enhanced insulin sensitivity and improved insulin resistance in experimental type 2 diabetes.
Chronic effects of remogliflozin etabonate on physiological and metabolic parameters in high-fat diet-fed GK rats Data are presented as means ± S.E.M. (n = 8 for Wistar rats, n = 8 for normal diet-fed GK rats, n = 10 for high-fat diet-fed GK rats). These rats were fed for 8 weeks a normal diet, high-fat diet, or high-fat diet containing 0.01, 0.03, or 0.1% remogliflozin etabonate. Plasma glucose, plasma insulin, GHb, and plasma triglyceride were monitored with the rats in the fed condition every 4 weeks. These rats were fasted for 16 h every 4 weeks, and the FPG and fasting plasma insulin were monitored. HOMA-R was also calculated.

Antihyperglycemic effects of remogliflozin etabonate in oral glucose tolerance test. A, plasma glucose (normal rats). B, plasma insulin (normal rats). C, plasma glucose (STZ-induced diabetic rats). D, AUC0–2 h for plasma glucose (STZ-induced diabetic rats). Remogliflozin etabonate and glucose solution (2 g/kg) were orally administered to normal rats and STZ-induced diabetic rats after 16-h fasting. Plasma glucose and plasma insulin were determined, and the AUC0–2 h for plasma glucose was calculated after the oral glucose tolerance test. Data are presented as means ± S.E.M. (n = 8 for normal rats and n = 6 for STZ-induced diabetic rats). *, p < 0.05 and **, p < 0.01 versus vehicle group.
Discussion
Multiple studies have demonstrated the antihyperglycemic effect of phlorizin in diabetic animal models. However, negative aspects of phlorizin (nonselectivity and low bioavailability) may be the reason for the lack of the usefulness of phlorizin as an antidiabetic drug. T-1095, a phlorizin derivative, was developed to overcome the low bioavailability of phlorizin. T-1095A, the active form of T-1095, inhibited both SGLT1 and SGLT2 as well as phlorizin.
To develop a selective SGLT2 inhibitor, we selected a basal scaffold structure different from that of phlorizin or T-1095 as the starting point for experimental approach. We obtained remogliflozin etabonate (prodrug) and remogliflozin (active form) by means of a screening system using recombinant SGLTs. Because its skeleton differs from that of phlorizin, T-1095, or sergliflozin, remogliflozin etabonate belongs to a new category of SGLT2 inhibitors. Based on Ki values, we confirmed remogliflozin to be potent and highly selective for human SGLT2, with a selectivity ratio of 365 (Table 1). According to Oku et al. (1999), the selectivity ratio of T-1095-A is approximately 4. In our previous study, sergliflozin-A, the active form of sergliflozin, showed a high selectivity ratio of 296 for SGLT2 (Katsuno et al., 2007). These results indicate that remogliflozin exhibited higher selectivity for SGLT2 than sergliflozin-A.

Antihyperglycemic effects of remogliflozin etabonate in db/db mice. Remogliflozin etabonate was orally administered to fed db/db mice. Blood glucose concentrations were measured (A), and the AUC0–6 h for blood glucose was calculated (B). Data are presented as means ± S.E.M. (n = 8). *, p < 0.05 and **, p < 0.01 versus vehicle group.
To examine the inhibitory effect on SGLT2, we orally administered remogliflozin etabonate to normal mice and rats. Urinary glucose excretion was increased by the oral administration of remogliflozin etabonate in a dose-dependent manner (Fig. 2). We also determined urinary glucose excretion by remogliflozin etabonate in normal dogs (data not shown). A dose-dependent effect was observed in each of these three species. We confirmed that remogliflozin etabonate suppressed the renal glucose reabsorption and increased urinary glucose excretion.
We examined the antihyperglycemic effects of remogliflozin etabonate induced by increasing urinary glucose excretion. In the oral glucose tolerance test, remogliflozin etabonate suppressed postprandial hyperglycemia in normal and STZ-induced diabetic rats (Fig. 3, A and C). The antihyperglycemic effect of remogliflozin etabonate was markedly enhanced in the diabetic state, because urinary glucose excretion was increased by remogliflozin etabonate to a greater degree in the hyperglycemic state than in the normoglycemic state. Remogliflozin etabonate also showed the antihyperglycemic effect in db/db mice (Fig. 4), which have more severe hyperglycemia than STZ-induced diabetic rats. Thus, remogliflozin etabonate promoted excretion of glucose in urine depending on the blood glucose level, and we presumed that remogliflozin etabonate would be useful over a range of diabetes from mild to severe. In addition, the antihyperglycemic effect of remogliflozin etabonate did not depend on an increase in insulin secretion (Fig. 3B). Remogliflozin etabonate is supposed to barely cause hypoglycemia, which is a major side effect with sulfonylureas. In fact, remogliflozin etabonate did not significantly alter the plasma glucose level in 16-h fasted normal rats (data not shown). The ability to induce lower blood glucose without stimulating insulin secretion is a property of remogliflozin etabonate that represents a considerable advantage for clinical use.
Chronic treatment with remogliflozin etabonate reduced both FPG and GHb levels in db/db mice and exhibited an antidiabetic profile (Table 2). In db/db mice, insulin is transiently oversecreted, reaching a maximal serum concentration by 8 to 10 weeks of age, and this oversecretion leads to β-cell dysfunction (Lenhard et al., 1999). By 3 months of age, male db/db mice lose weight, and their death rate increases (Lenhard et al., 1999). In this study, we confirmed that the vehicle-treated db/db mice lost weight corresponding to the reduction in plasma insulin. In contrast, remogliflozin etabonate inhibited the decrease in plasma insulin levels while improving hyperglycemia, and body weight gain was observed at a high dose of remogliflozin etabonate. The preservation of plasma insulin levels by remogliflozin etabonate was due to an antihyperglycemic mechanism that was not dependent on insulin secretion. A similar observation has been reported in T-1095-treated db/db mice (Arakawa et al., 2001). We suggest that amelioration of hyperglycemia with remogliflozin etabonate prevents the development of β-cell exhaustion and improves glucose metabolism. It is known that chronic hyperglycemia leads to the impairment of insulin secretion and to insulin resistance of peripheral tissues. Thus, excretion of excess plasma glucose via urine can contribute to the normalization of glucose metabolism by suppressing glucose toxicity. The chronic exposure of the kidney to excess glucose due to the action of remogliflozin etabonate may be of concern regarding renal dysfunction. However, renal glucosuria is defined as urinary glucose excretion in the presence of a normal blood glucose level and in the absence of any signs of general renal tubular dysfunction (Santer et al., 2003). The acute treatment with remogliflozin etabonate increased the urinary glucose excretion (Fig. 2), whereas the chronic treatment with it ameliorated the glucosuria by improving glycemic control (Table 2). Thus, the glucose content in the glomerular filtrate would decrease along with a reduction in the hyperglycemia, and the chronic treatment with remogliflozin etabonate might thus have a protective effect on the kidneys.
High-fat intake and obesity are considered as causative environmental factors in the development of type 2 diabetes, hyperinsulinemia, and insulin resistance (Storlien et al., 1986). GK rats are generally used as a diabetic model with age-dependent impaired insulin secretion (Portha et al., 1991; Hughes et al., 1994). In this study, GK rats were fed a high-fat diet to induce hyperinsulinemia and insulin resistance. Hyperglycemia, hyperinsulinemia, hypertriglyceridemia, and insulin resistance were observed, as shown in Table 3. As in the case of db/db mice, chronic treatment with remogliflozin etabonate improved hyperglycemia and hypertriglyceridemia in the high-fat diet-fed GK rats (Table 3). In addition, remogliflozin etabonate reduced both the fasting plasma levels of glucose and insulin as well as the value of HOMA-R, and it reduced AUCs for plasma glucose and plasma insulin in the oral glucose tolerance test (Fig. 5). We suggest that remogliflozin etabonate enhanced the insulin sensitivity, decreased the gross amount of insulin secretion, and thus normalized the glycemic response after glucose loading. Thus, chronic treatment with remogliflozin etabonate enhanced insulin sensitivity and improved insulin resistance. Weight loss was observed at a high dose of remogliflozin etabonate in the high-fat diet-fed GK rats. The epididymal and retroperitoneal fat weights were also decreased in these rats (data not shown). Metabolic syndrome, a condition that is associated with type 2 diabetes, requires lifestyle modifications aimed at reducing obesity and insulin resistance (Garber, 2004). Remogliflozin etabonate may have therapeutic potential for the treatment of metabolic syndrome and obesity. This potential will need to be assessed in further studies.

Chronic effects of remogliflozin etabonate on plasma glucose (A), AUC0–4 h for plasma glucose (B), plasma insulin (C), and AUC0–4 h for plasma insulin (D) during the oral glucose tolerance test. Wistar, Wistar rats; GK (ND), normal diet-fed GK rats; GK (HFD), high-fat fed-GK rats. ††, p < 0.01 versus Wistar rats. ##, p < 0.01 versus normal diet-fed GK rats. *, p < 0.05 and **, p < 0.01 versus high-fat fed-GK rats.
Type 2 diabetes has increased in incidence as lifestyles involving excessive calorie intake and lack of exercise have become more prevalent. The basic management of type 2 diabetes involves controlling the body's energy balance by dietary means and by increasing exercise levels. In addition, several types of oral drugs are available for the clinical control of the blood glucose level. However, it is difficult to maintain good glycemic control in diabetic patients, and there is a lack of drugs that can be used to control energy balance in a negative direction. In this study, we demonstrated that remogliflozin etabonate was a highly selective SGLT2 inhibitor that increased urinary glucose excretion and consequently had antihyperglycemic effects, without stimulating insulin secretion. An increase in urinary glucose excretion via inhibition of SGLT2 by remogliflozin etabonate leads to a negative energy balance. These properties of remogliflozin etabonate enable it to be suitable for blood glucose control with preservation of insulin secretion. In this sense, this SGLT2 inhibitor offers some advantages over existing antidiabetic drugs such as sulfonylureas, biguanides, α-glucosidase inhibitors, and thiazolidinediones. Multiple studies on T-1095 have demonstrated its beneficial effects on postprandial hyperglycemia, FPG, GHb, insulin resistance, β-cell function, and diabetic complications (nephropathy and neuropathy) in diabetic rodent models (Adachi et al., 2000; Nawano et al., 2000; Oku et al., 2000; Arakawa et al., 2001; Nunoi et al., 2002; Ueta et al., 2005; Fujimoto et al., 2006). We suppose that remogliflozin etabonate may also be useful for a wide range of diabetic conditions, and it can be beneficial for the treatment of diabetes when used in combination with a number of antidiabetic drugs.
In summary, remogliflozin etabonate may achieve long-term glycemic control, improve insulin resistance, and preserve pancreatic β-cell function without either inducing body weight gain or increasing the risk of hypoglycemia. Clinical trials on remogliflozin etabonate are currently in progress (Ashiya and Smith, 2007; Isaji, 2007), and proof-of-concept studies should clarify the suitability of remogliflozin etabonate for the treatment of type 2 diabetes.
Footnotes
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.108.140210.
-
ABBREVIATIONS: SGLT2, low-affinity sodium glucose cotransporter; SGLT1, high-affinity sodium glucose cotransporter; UGE, urinary glucose excretion; T-1095, 3-(benzo[b]furan-5-yl)-2′,6′-dihydroxy-4′-methylpropiophenone-2′-O-(6-O-methoxycarbonyl)-β-d-glucopyranoside; T-1095A, active form of T-1095; AMG, methyl-α-d-glucopyranoside; GK, Goto-Kakizaki; CMC, sodium carboxymethylcellulose; STZ, streptozotocin; AUC, area under the curve; FPG, fasting plasma glucose; GHb, glycated hemoglobin; HOMA-R, homeostasis model assessment ratio.
- Received April 17, 2008.
- Accepted June 25, 2008.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}