


Abstract
Two H+/peptide cotransporters, PEPT1 and PEPT2, are expressed in the kidney, mediating the renal tubular reabsorption of oligopeptides and β-lactam antibiotics. We examined the interactions of β-lactam antibiotics with peptide transporters in rat renal brush-border membranes by evaluating the inhibitory potencies of the antibiotics against glycylsarcosine transport. Western blot analysis revealed that PEPT1 and PEPT2 were expressed in the renal brush-border membranes with the apparent molecular masses of 75 and 105 kDa, respectively. Using renal brush-border membrane vesicles, the uphill transport of glycylsarcosine was observed in the presence of an inward H+ gradient and an inside-negative membrane potential. Two transport systems with high affinity (Km of 50 μM) and low affinity (Km of 1.2 mM) appeared kinetically to mediate the glycylsarcosine uptake. The inhibition constants of the antibiotics for glycylsarcosine transport were more closely correlated with those in stable LLC-PK1 cells transfected with rat PEPT2 rather than PEPT1 cDNA. The β-lactam antibiotics with an α-amino group showed trans-stimulation effects on the glycylsarcosine uptake, suggesting that these antibiotics and glycylsarcosine share a common peptide transporter. However, the antibiotics lacking an α-amino group failed to show thetrans-stimulation effect. It is concluded that amino-β-lactam antibiotics at therapeutic concentrations interact predominantly with PEPT2 localized in the brush-border membranes of rat kidney.
Peptide transporters expressed in brush-border membranes of the intestinal and renal epithelial cells are involved in the efficient absorption of oligopeptides, thereby contributing to the maintenance of protein nutrition (Leibach and Ganapathy, 1996). It has been demonstrated that the process of peptide transport is electrogenic H+-coupled cotransport driven by the presence of an inward H+ gradient and a negative membrane potential (Ganapathy and Leibach, 1983; Takuwaet al., 1985). Peptide transporters play pharmacological roles in some medications, because peptide transporters mediate the transport of peptide-like drugs such as β-lactam antibiotics (Okanoet al., 1986; Tsuji et al., 1987), bestatin (an antineoplastic drug) (Inui et al., 1992; Saito and Inui, 1993) and angiotensin converting enzyme inhibitors (Swaan et al., 1995). Using purified brush-border membrane vesicles from rat kidney cortex, we provided the first evidence to show that oral β-lactam antibiotics and dipeptide share a common peptide transport system (Inui et al., 1984). In addition, it has been suggested that there are at least two distinct peptide transporters in the brush-border membranes of renal proximal tubules (Daniel et al., 1991; Miyamoto et al., 1988; Naasani et al., 1995).
Molecular cloning studies have clarified that two distinct peptide transporters, designated PEPT1 and PEPT2, are expressed in the cell plasma membranes of rat (Saito et al., 1995, 1996), rabbit (Boll et al., 1996; Fei et al., 1994) and human (Liang et al., 1995; Liu et al., 1995;Ramamoorthy et al., 1995). Rat PEPT1 appeared to be localized at the brush-border membranes along the digestive tract (duodenum, jejunum and ileum) and to a lesser extent at brush-border membranes from the kidney cortex (Ogihara et al., 1996). A Northern blot analysis suggested that rat PEPT2 mRNA was expressed predominantly in the kidney, but not in the intestine (Saito et al., 1996). Although the findings suggest that both PEPT1 and PEPT2 are expressed in the kidney, their relative contributions to the tubular reabsorption of oligopeptides and peptide-like drugs remain unknown. We recently established a renal cell line stably transfected with rat PEPT1 (Terada et al., 1996, 1997a) and with PEPT2 (Terada et al., 1997b), and have compared their affinities for various β-lactam antibiotics. We found that rat PEPT2 has much higher affinity for β-lactam antibiotics with an α-amino group than does PEPT1 (Terada et al., 1997b). In our study, we investigated the interactions of β-lactam antibiotics with these two peptide transporters by comparing the inhibitory potencies of the antibiotics against glycylsarcosine transport in the renal brush-border membrane vesicles with those in stable transfectants expressing rat PEPT1 or PEPT2.
Materials and Methods
Materials.
Cefixime (Fujisawa Pharmaceutical Co., Osaka, Japan), ceftibuten and cephalexin (Shionogi and Co., Osaka, Japan), cefadroxil (Bristol Meyers Co., Tokyo, Japan), cephradine (Sankyo Co., Tokyo) and cyclacillin (Takeda Chemical Industries, Osaka) were gifts from the respective suppliers. [14C]Glycylsarcosine (1.78 GBq/mmol) was obtained from Daiichi Pure Chemicals Co. (Ibaraki, Japan). Glycylsarcosine, valinomycin and ampicillin were obtained from Sigma Chemical Co. (St. Louis, MO). All other chemicals used were of the highest purity available. Figure 1shows the structures of the penicillin and cephalosporin antibiotics used.

Chemical structures of penicillins and cephalosporins.
Preparation of brush-border membrane vesicles.
The brush-border membrane vesicles were isolated from the renal cortex of male Wistar albino rats (200–240 g) by the Mg++/EGTA precipitation method as described previously (Hori et al., 1993). The isolated membrane vesicles were suspended in an experimental buffer to give a final protein concentration of 10 mg/ml. In general, the experimental buffer consisted of either 100 mM mannitol, 100 mM potassium gluconate and 10 mM HEPES (pH 7.5) or 100 mM mannitol, 100 mM potassium gluconate and 10 mM MES (pH 6.0), and the pH was adjusted with KOH.
Uptake studies.
The uptake of [14C]glycylsarcosine by brush-border membrane vesicles was measured by a rapid filtration technique. Usually, membrane vesicles suspended in an appropriate buffer were preincubated for 10 min at 37°C before the initiation of uptake. For the measurement of the trans-stimulation effect, the vesicles were preincubated for 60 min at room temperature to equilibrate the vesicles with a compound to be tested. The uptake was initiated by the addition of 180 μl of a buffer containing 22.2 μM [14C]glycylsarcosine (final 20 μM) to 20 μl of membrane suspension at 37°C. At the specified periods, the incubation was terminated by diluting the reaction mixture with 1 ml of ice-cold stop solution containing 150 mM KCl and 20 mM HEPES/TRIS (pH 7.5). The mixture was poured immediately onto Millipore filters (HAWP, 0.45 μm, 2.5-cm diameter), and the filters were washed once with 5 ml of ice-cold stop solution. In separate experiments, the nonspecific association of the ligand with vesicles was estimated by the addition of substrate mixture to 1 ml of ice-cold stop solution containing 20 μl of membrane vesicles. This value was subtracted from the uptake data to evaluate the specific uptake of the ligand. The radioactivity of [14C]glycylsarcosine trapped in membrane vesicles was determined in ACS II (Amersham International, Buckinghamshire, UK) by liquid scintillation counting. The protein content was determined by the method of Bradford (1976), using a protein assay kit (Bio-Rad, Richmond, CA) with bovine γ-globulin as the standard.
Western blot analysis.
Crude plasma membrane fractions from rat tissues were prepared as described (Ogihara et al., 1996). The crude plasma membranes and renal brush-border membranes were analyzed for the expression of the PEPT1 and PEPT2 by immunoblotting as reported previously (Saito et al., 1995). The antiserum against rat PEPT1 was raised previously, and the antiserum for rat PEPT2 was raised against the synthetic peptide corresponding to the deduced 697 to 710 amino acid sequence of rat PEPT2 after being coupled to cystein at the carboxy-terminus and linked to keyhole limpet hemocyanin as described (Saito et al., 1995).
Results
Immunoblotting.
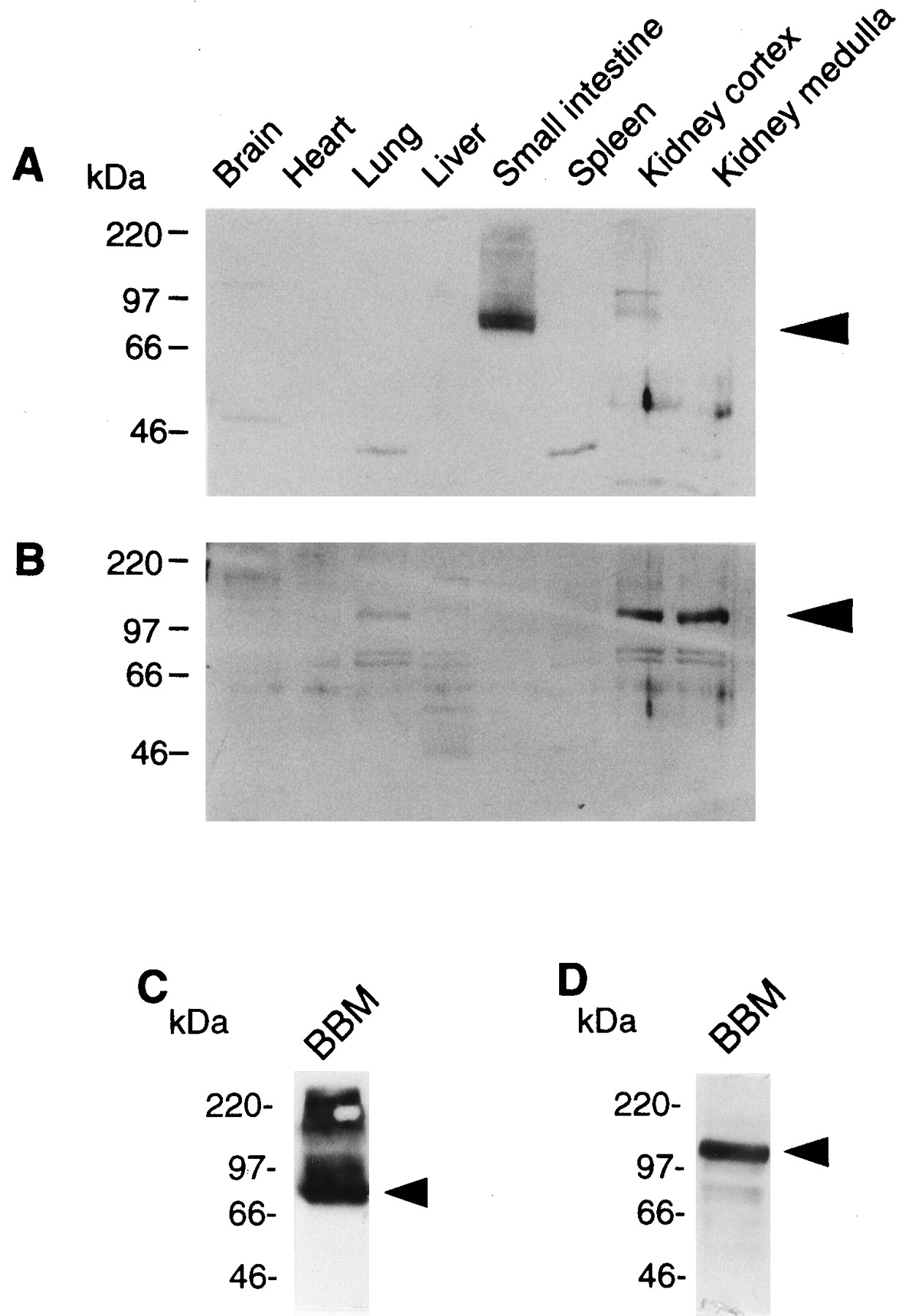
The tissue distributions of PEPT1 and PEPT2 were examined by Western blot analysis. As shown in figure2A, PEPT1 was expressed predominantly in the small intestine, and to a lesser extent in the kidney cortex. In contrast, PEPT2 was found almost exclusively in the kidney cortex and medulla, and faintly in the lung, but not in the small intestine (fig.2B). In addition, PEPT1 and PEPT2 proteins were detected on the renal brush-border membranes with the apparent molecular masses of 75 and 105 kDa, respectively (fig. 2, C and D). These results indicate that both PEPT1 and PEPT2 are expressed at the brush-border membranes in the kidney.

Western blot analysis of crude plasma membranes from rat tissues (A and B) and of renal brush-border membranes (C and D) for PEPT1 (A and C) and PEPT2 (B and D). Crude plasma membranes (25 μg) from each tissue and renal brush-border membranes (100 μg) were separated on sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) (10%) and blotted on a polyvinylidene difluoride (PVDF) membrane. The antisera (1:1000 dilution) were used as primary antibodies. A horseradish peroxidase-conjugated antirabbit IgG antibody was used for the detection of bound antibodies, and the strips of blots were visualized by chemiluminescence on x-ray films. The arrowheads indicate the positions of each transporter.
Effects of H+ gradient and membrane potential on glycylsarcosine uptake.
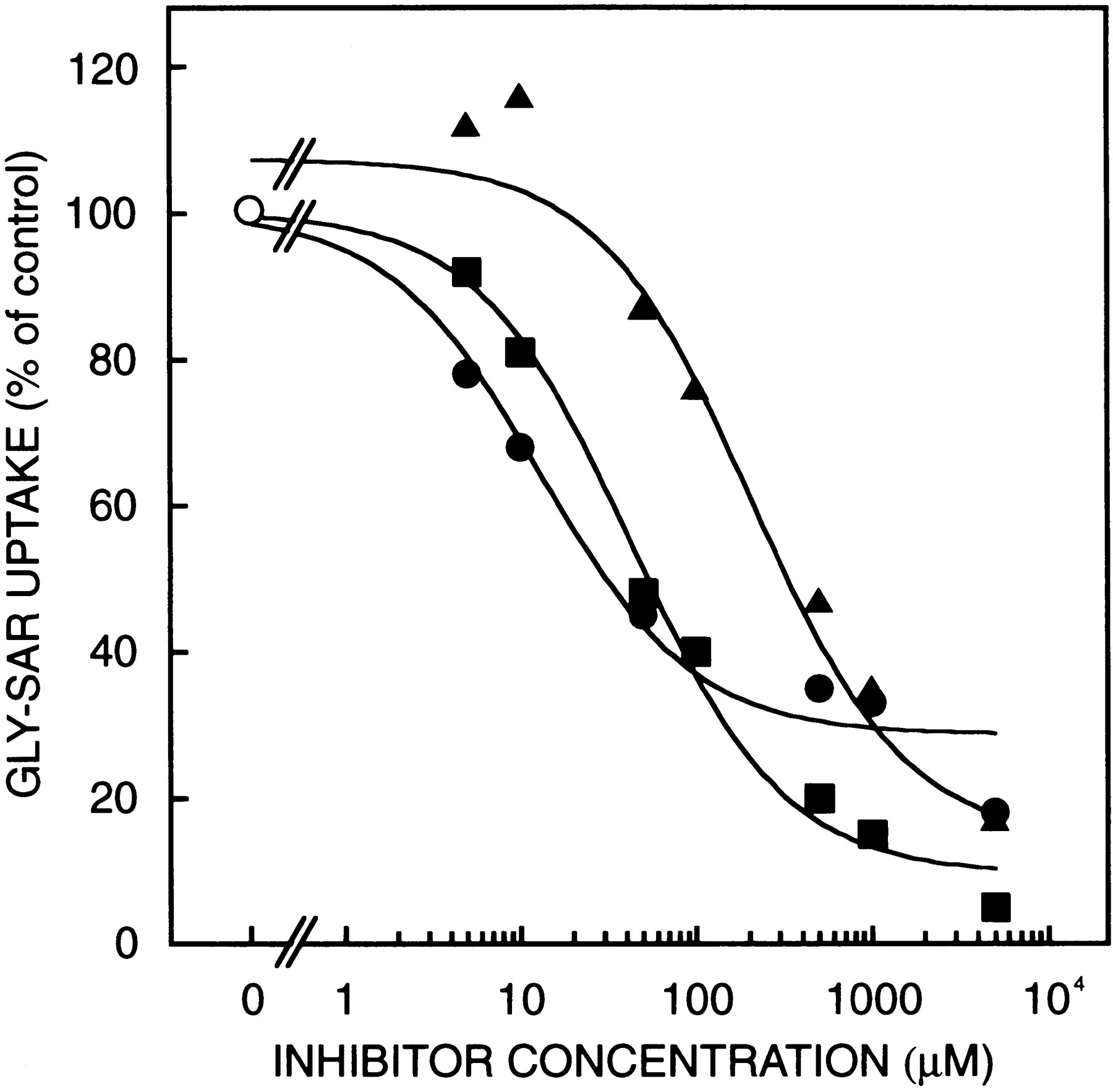
The effects of an inward H+gradient and K+ diffusion potential (interior-negative) induced by valinomycin on glycylsarcosine uptake were examined. As shown in figure 3, the initial rate of glycylsarcosine uptake was markedly stimulated in the presence of an interior-negative membrane potential compared to that in the absence of membrane potential, showing transient uphill transport, although the initial rate of glycylsarcosine uptake was not stimulated in the presence of H+ gradient. When an inward H+gradient and an interior-negative membrane potential were imposed simultaneously, the glycylsarcosine uptake was further stimulated. These results clearly demonstrated that functional peptide transporter(s) which could be driven by the H+ gradient and membrane potential exist in the rat brush-border membranes.

Effects of H+ gradient and membrane potential on the glycylsarcosine uptake by rat renal brush-border membrane vesicles. Membrane vesicles (20 μl) suspended in 100 mM mannitol, 100 mM potassium gluconate and 10 mM HEPES (pH 7.5) were incubated at 37°C with the substrate mixture (180 μl) comprised of 100 mM mannitol, 100 mM sodium gluconate, 22.2 μM [14C]glycylsarcosine, either 10 mM MES (pH 6.0) (•, ○), or 10 mM HEPES (pH 7.5) (▴, ▵) and 0.4% ethanol in the presence (•, ▴) or absence (▵, ○) of 3 mg/ml valinomycin. Each point represents the mean ± S.E. of three experiments.
Kinetic analysis of glycylsarcosine uptake.
To investigate the kinetic features of glycylsarcosine uptake by brush-border membrane vesicles, the initial uptake rate of glycylsarcosine was examined as a function of the glycylsarcosine concentration. The uptake measurements were done in the presence of both an inward H+ gradient and an inside-negative membrane potential. As illustrated in figure4, the resulting glycylsarcosine uptake was curvilinear. With the use of a nonlinear least-square regression analysis with the Michaelis-Menten equation, the kinetic parameters of this uptake were calculated. The Eadie-Hofstee plot of the data after correcting the nonsaturable component is shown in the inset of figure4. These data indicate that the glycylsarcosine uptake consists of two saturable components: one with an apparent Km , the Michaelis-Menten constant, of 1235 μM and Vmax, the maximum uptake rate, of 2038 pmol/mg protein/10 sec (low affinity), and another with an apparent Km of 50 μM and Vmax of 175 pmol/mg protein/10 sec (high affinity). The nonsaturable component had an apparent Kd , the coefficient of simple diffusion, of 103 pmol/mg protein/10 sec/mM. TheKm values for the low affinity and high affinity components estimated in the brush-border membranes were comparable with the Km values of the cloned PEPT1 (1100 μM) and PEPT2 (110 μM) evaluated in the stable transfectants, respectively (Terada et al., 1997b). These results suggest that glycylsarcosine uptake in rat renal brush-border membranes is mediated by both PEPT1 and PEPT2.

Concentration dependence of glycylsarcosine uptake by rat renal brush-border membrane vesicles. Membrane vesicles (20 μl), suspended in 100 mM mannitol, 100 mM potassium gluconate and 10 mM HEPES (pH 7.5), were incubated at 37°C for 10 sec with the substrate mixture (180 μl) comprised of 100 mM mannitol, 100 mM sodium gluconate, 10 mM MES (pH 6.0) and 0.4% ethanol in the presence of 3 mg/ml valinomycin and various concentrations of [14C]glycylsarcosine. Each point represents the mean ± S.E. of three experiments. Inset, Eadie-Hofstee plot of the uptake after correction for the nonsaturable component. V, uptake rate (nmol/mg protein/10 sec); S, glycylsarcosine concentration (mM).
Inhibition of glycylsarcosine uptake by various β-lactam antibiotics.
To investigate the relative contributions of PEPT1 and PEPT2 to the renal reabsorption of β-lactam antibiotics, the ability of various oral β-lactam antibiotics to inhibit the uptake of glycylsarcosine was studied. As shown in figure5, cefadroxil (aminocephalosporin), cyclacillin (aminopenicillin) and ceftibuten (anionic cephalosporin without an α-amino group) inhibited glycylsarcosine uptake in a dose-dependent fashion. The glycylsarcosine uptake was inhibited by the antibiotics in the following order of inhibitory potency: cefadroxil > cyclacillin > ceftibuten. Based on the nonlinear least-square regression analysis for the competition curves, the apparent inhibition constants (Ki ) for several antibiotics were estimated. Figure6 shows the correlations of the apparentKi values of β-lactam antibiotics between the brush-border membranes and the PEPT1 or PEPT2 transfectants (Teradaet al., 1997b). The Ki values of β-lactam antibiotics for the brush-border membranes were closely correlated with those for PEPT2 (r = 0.98), but not for PEPT1 (r = 0.35).

Inhibition of glycylsarcosine uptake by β-lactam antibiotics in rat renal brush-border membranes. Membrane vesicles (20 μl), suspended in 100 mM mannitol, 100 mM potassium gluconate and 10 mM HEPES (pH 7.5), were incubated at 37°C for 10 sec with the substrate mixture (180 μl) comprised of 100 mM mannitol, 100 mM sodium gluconate, 10 mM MES (pH 6.0) and 0.4% ethanol in the presence of 3 mg/ml valinomycin and 22.2 μM [14C]glycylsarcosine in the presence of increasing concentrations of cyclacillin (■), cefadroxil (•) and ceftibuten (▴). Uptake of [14C]glycylsarcosine measured in the absence (○) of the inhibitors was taken as 100% (104.0 ± 3.9 pmol/mg protein/10 sec). Each point represents the mean of three experiments.
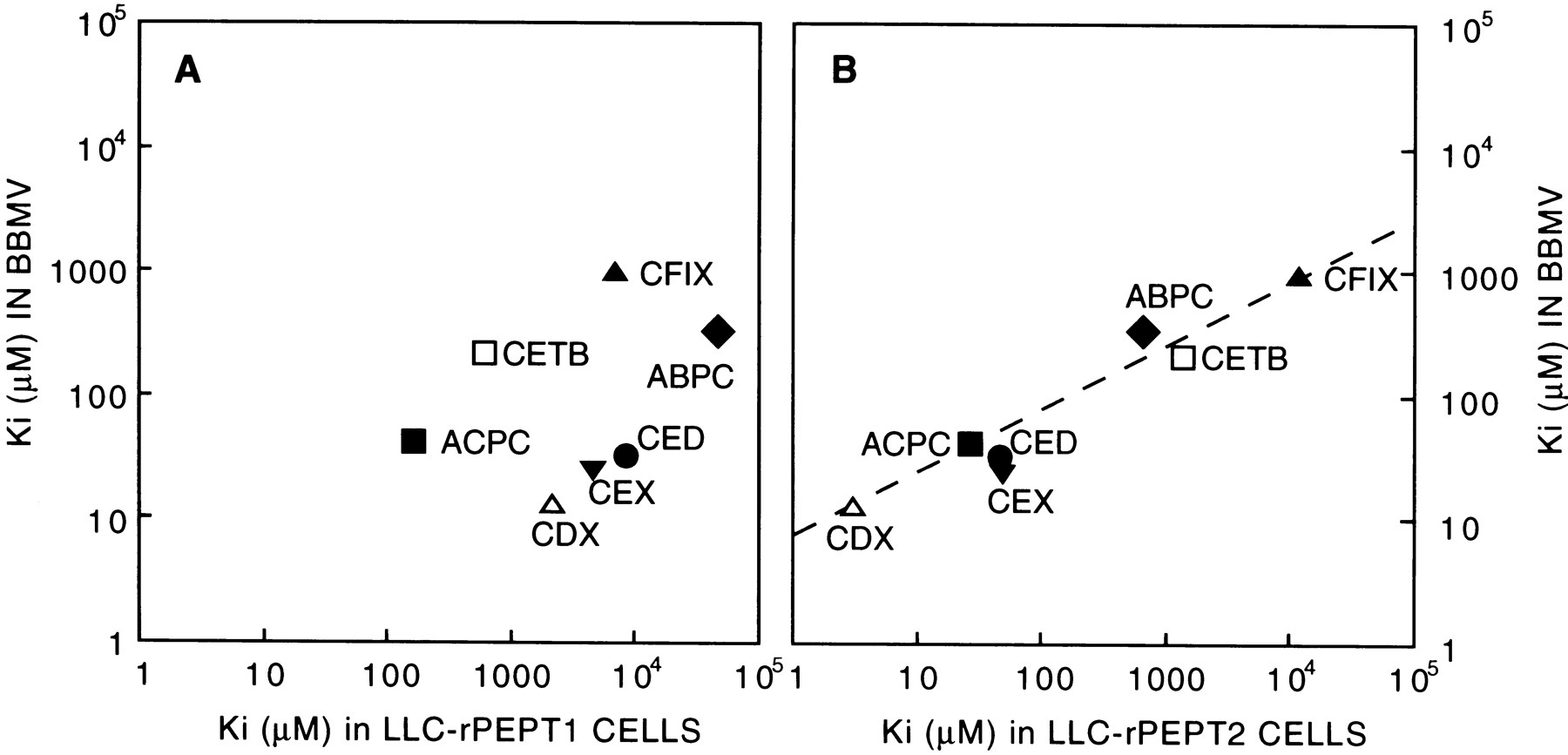
Correlations of the apparentKi values of various β-lactam antibiotics for [14C]glycylsarcosine uptake between rat renal brush-border membrane vesicles (BBMV) and PEPT1 (A) or PEPT2 (B) transfectant. The Ki values in the PEPT transfectants were taken from the reference (Terada et al., 1997b). ABPC, ampicillin; ACPC, cyclacillin; CDX, cefadroxil; CED, cephradine; CETB, ceftibuten; CEX, cephalexin; CFIX, cefixime.
Trans-stimulation effects of β-lactam antibiotics on glycylsarcosine uptake.
To determine directly whether the β-lactam antibiotics and glycylsarcosine share a common peptide transporter in rat renal brush-border membranes, thetrans-stimulation effects of the β-lactam antibiotics on glycylsarcosine uptake were studied. As depicted in figure7, the initial uptake rate of [14C]glycylsarcosine was markedly enhanced in the vesicles preloaded with unlabeled glycylsarcosine. The [14C]glycylsarcosine uptake was also stimulated in the vesicles preloaded with cephalexin > cephradine > cyclacillin (in the order of the stimulatory potency). Ampicillin, ceftibuten and cefixime had less effect on the uptake. The uptake of glycylsarcosine was inhibited by cefadroxil (1 mM). Cefadroxil at 1 mM might act as an inhibitor in the trans-stimulation study, because cefadroxil has a very high affinity to PEPT2 with an apparentKi value of 3 μM (Terada et al., 1997b). In a separate experiment, the initial uptake rate of glycylsarcosine showed a tendency to be stimulated in the vesicles preloaded with cefadroxil at a concentration of 0.1 mM: control, 23.2 ± 1.9; 0.1 mM cefadroxil, 26.9 ± 0.6, pmol/mg protein/10 sec, mean ± S.E. of three determinations. These results suggested that the cephalosporins with an α-amino group in the molecules shared a common peptide transporter, presumably PEPT2, with glycylsarcosine, whereas anionic cephalosporins such as ceftibuten and cefixime had much lower affinity for the transporter, showing notrans-stimulation effect.

Trans-stimulation effects of β-lactam antibiotics on glycylsarcosine uptake by rat renal brush-border membrane vesicles. Membrane vesicles were preincubated at room temperature for 1 hr in 100 mM mannitol, 100 mM potassium gluconate and 10 mM HEPES (pH 7.5) with 1 mM β-lactam antibiotics and then aliquots (20 μl) were incubated at 37°C for 10 sec with the substrate mixture (180 μl) comprised of 100 mM mannitol, 100 mM potassium gluconate, 10 mM HEPES (pH 7.5) and 22.2 μM [14C]glycylsarcosine. Each column represents the mean ± S.E. of three experiments. *P < .05, significant difference from control using analysis of variance followed by Fisher’s t test.
Discussion
Studies using isolated membrane vesicles have shown that the uptake of oligopeptides in renal brush-border membranes is mediated by at least two distinct transport systems: the high affinity/low capacity and the low affinity/high capacity peptide transporters (Danielet al., 1991). We recently isolated two cDNA encoding rat H+/peptide cotransporters, PEPT1 and PEPT2 (Saito et al., 1995, 1996). By Northern blot analysis and polymerase chain reaction amplification, we found that PEPT1 and PEPT2 are expressed in the kidney, suggesting that these two transporters would contribute to the tubular reabsorption of oligopeptides. However, relative contribution of each cloned peptide transporter to the renal reabsorption of β-lactam antibiotics has not been fully elucidated.
By Western blot analysis of plasma membrane fractions prepared from the rat kidney, both PEPT1 and PEPT2 were expressed on the brush-border membranes (fig. 2, C and D). PEPT2 protein was also detected in the lung (fig. 2B), as observed by the Northern blot analysis of rat PEPT2 mRNA (Saito et al., 1996). Meredith and Boyd (1995)suggested the presence of a proton-coupled peptide transport protein in the lung; thus, PEPT2 may play an important role in lung homeostasis.
We examined the effects of an inward H+ gradient and an interior-negative membrane potential on the glycylsarcosine uptake by the brush-border membrane vesicles. In the presence of both the H+ gradient and membrane potential, glycylsarcosine uptake was markedly stimulated, showing a transient uphill transport (fig. 3). It is noteworthy that the uphill transport of the glycylsarcosine was also observed in the presence of an inside-negative membrane potential alone, but not in the presence of H+ gradient alone. Therefore, our results suggest that both the inward H+gradient and the interior-negative membrane potential play important roles in the active transport of oligopeptides in renal tubular cells.
Our kinetic analysis clearly indicated that the glycylsarcosine uptake in the renal brush-border membranes was mediated by two transport systems (fig. 4). One system was characterized as the low-affinity and high-capacity type, and the other as the high-affinity and low-capacity type. It was reported previously that there are at least two distinct dipeptide transporters in the renal brush-border membranes (Danielet al., 1991). The differences in the substrate affinity for glycylsarcosine transport suggest that these transporters play distinct nutritional roles in the renal tubular reabsorption of oligopeptides. Importantly, our findings revealed that the Km values in rat PEPT1 and PEPT2 were comparable to those of the low-affinity and high-affinity transport systems in renal brush-border membranes, respectively. Therefore, it seems reasonable to speculate that the two distinct peptide transport systems in the renal brush-border membranes are constituted of PEPT1 and PEPT2.
We reported that various β-lactam antibiotics showed different inhibitory potencies against glycylsarcosine uptake between rat PEPT1 and PEPT2 transfectants (Terada et al., 1997b), and found that PEPT1 had much higher affinity for cyclacillin than for cefadroxil, whereas PEPT2 appeared to prefer cefadroxil to cyclacillin. The findings are similar to those reported for the human PEPT1 and PEPT2, suggesting the differential recognition of β-lactam antibiotics (Ganapathy et al., 1995). In our study, the inhibitory potencies of β-lactam antibiotics in the brush-border membranes were more closely correlated to those observed in rat PEPT2 transfectant rather than rat PEPT1 transfectant (fig. 6). The results suggest that these antibiotics interact preferentially with PEPT2 rather than with PEPT1 when evaluated by the competitive inhibition of glycylsarcosine transport at a concentration of 20 μM. Indeed, the uptake rate of glycylsarcosine estimated by the Michaelis-Menten equation including two carrier-mediated components at 20 μM is 90 pmol/mg protein/10 sec, to which the high-affinity and low-affinity systems contribute 56 and 36%, respectively. Similarly, the contributions of the high- and low-affinity systems at 1 mM glycylsarcosine are 14 and 77%, respectively. Therefore, the contribution of the higher affinity system, namely PEPT2, to the net transport of the dipeptide should be greater with low substrate concentrations.
The initial uptake rate of glycylsarcosine was stimulated in the vesicles preloaded with cephalosporins with an α-amino group in molecules (fig. 7), suggesting that these antibiotics are transported via the same transport system for glycylsarcosine uptake. Ampicillin, ceftibuten and cefixime, with relatively higher apparentKi values than those of the aminocephalosporins, failed to show the trans-stimulation effect. By using PEPT transfectants, we found that PEPT2 had much lower affinity for cephalosporins lacking an α-amino group (Terada et al., 1997b). The finding that cephalosporins without an α-amino group showed no trans-stimulation also suggests that predominantly PEPT2 contributes to the net transport of glycylsarcosine in rat renal brush-border membranes.
In conclusion, we have confirmed that two peptide transporters, PEPT1 and PEPT2, are localized to rat renal brush-border membranes. Our correlation analysis based on the apparent Ki values for glycylsarcosine transport suggests that β-lactam antibiotics at therapeutic concentrations interact predominantly with the high-affinity transporter PEPT2 rather than with the low-affinity transporter PEPT1 in rat renal brush-border membranes. Further studies on the distribution and expression levels of PEPT1 and PEPT2 along the nephron will clarify their relative contributions to the tubular reabsorption of β-lactam antibiotics.
Footnotes
-
Send reprint requests to: Professor Ken-ichi Inui, Department of Pharmacy, Kyoto University Hospital, Sakyo-ku, Kyoto 606-8507, Japan.
-
↵1 This work was supported in part by a Grant-in-Aid for Scientific Research on Priority Areas of “Channel-Transporter Correlation” from the Ministry of Education, Science, and Culture of Japan, and by a Grant-in-Aid from Yamanouchi Foundation for Research on Metabolic Disorders.
- Abbreviations:
- HEPES
- N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid
- MES
- 2-(N-morpholino)-ethanesulfonic acid
- TRIS
- 2-amino-(2-hydroxymethyl)-1,3-propanediol
- Received December 3, 1997.
- Accepted April 13, 1998.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}